










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencia forestal en México
versión impresa ISSN 1405-3586
Rev. Cien. For. Mex vol.34 no.105 México ene./jun. 2009
Nota técnica
Efecto de Tillandsia recurvata L. (Bromeliaceae) sobre el éxito reproductivo de Fouquieria splendens Engelm. (Fouquieriaceae)
Iván Castellanos–Vargas1, Zenón Cano–Santana1 y Berta Hernández–López1
1 Departamento de Ecología y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Autónoma de México. Correo–e: icv@ciencias.unam.mx.
Fecha de recepción: 07 de septiembre de 2007.
Fecha de aceptación: 01 de abril de 2009.
RESUMEN
Algunas plantas epífitas son únicamente parásitas de espacio. Se describe a continuación el efecto que provoca la infestación de Tillandsia recurvata sobre el éxito reproductivo de Fouquieria splendens. El estudio se desarrolló en un matorral micrófilo–espinoso en Peña Blanca, Querétaro. En abril de 2005 se seleccionaron 66 individuos de F. splendens, a los cuales se les registró la cobertura, el número de agregaciones de T. recurvata, la cobertura promedio de tres agregaciones por cada forofito, el número de panículas, el número de flores con frutos y el número de flores muertas por panícula. El nivel de infestación de T. recurvata sobre F. splendens estuvo directamente correlacionado con el potencial reproductivo por unidad de área de cobertura del forofito (r = 0.31, P = 0.014); asimismo, hubo una correlación positiva y significativa entre el nivel de infestación y el número de flores muertas (r = 0.33, P = 0.007). Sin embargo, no se encontró correlación significativa entre el nivel de infestación y la proporción de frutos producidos por unidad de área de cobertura de F. splendens (r = 0.05, P = 0.71). Los resultados sugieren la existencia de una compensación reproductiva. Es probable que cuando existen altos niveles de infestación por T. recurvata, el forofito de F. splendens incrementa la producción de flores para así asegurar que el número de éstas con semillas se mantenga en la misma proporción que tendría si no estuviera infestada.
Palabras clave: Epífita, éxito reproductivo, Fouquieria splendens, panículas, Querétaro, Tillandsia recurvata.
ABSTRACT
Some epiphyte plants are space parasites only. The effect that causes infestation of Tillandsia recurvata aggregations on reproductive success of Fouquieria splendens was analyzed. The study was carried out in a microphyll–thorny xeric scrub at Peña Blanca, Queretaro. In April 2005, sixty six individuals of F. splendens were selected, to register the cover, number of T. recurvata aggregations guests, cover mean of three T. recurvata aggregations by each phorophyte, panicles number, fowers with seeds and dead fowers by panicle. The infestation level of T. recurvata on F. splendens was correlated directly with number of fowers produced by cover phorophyte unit area (r = 0.31, P = 0.014); also, a signifcant positive correlation between infestation level and number of dead fowers was found (r = 0.33, P = 0.007). Nevertheless, there was not a signifcant correlation between infestation level and seeds produced by cover unit area of F. splendens (r = 0.05, P = 0.71). Results suggest that there is a possible reproductive compensation. When high infestation leve Isby T. recurvataexist, F. splendens phorophyte responds by increasing the production fowers in order to assure that the same number of structures with seeds stays in a similar proportion as if it was not infested.
Key words: Epiphyte, reproductive success, Fouquieria splendens, panicles, Queretaro, Tillandsia recurvata.
Las plantas epífitas tienen la ventaja de colonizar el dosel de árboles y arbustos sin la necesidad de invertirrecursos en la producción de sistemas vasculares complejos (Madison, 1977; Smith et al., 1986; Benzing, 1990; Reyes–García, 2001) y en algunas ocasiones, no establecen relaciones metabólicas con el forofito (Lüttge, 1989; Richards, 1995); sin embargo, han sido llamadas "parásitas de espacio" debido a que compiten con el hospedero por la luz (Valdivia, 1977; Lüttge, 1989; Richards, 1995). En contra parte, algunas veces son consideradas como comensalistas, excepto cuando modifican significativamente la morfología del forofito y además le ocasionan daños físicos (Benzing, 1990).
Por ejemplo, Stevens (1987) indica que las lianas son parásitos estructurales de los árboles femeninos de Bursera simaruba (L.) Sarg. Las lianas para crecer necesitan estar en contacto físico con el forofito pero no extraen agua ni nutrimentos de su sistema vascular y sin embargo, su presencia reduce la producción de frutos del forofito.
Se ha registrado que una alta densidad de epífitas en una rama induce infecciones microbianas que pueden desembocar en su putrefacción y ruptura (Lüttge, 1989). La epifitosis promueve que la planta asigne mayor proporción de sus recursos a la defensa contra la infestación, que a su reproducción (Valdivia, 1977).
El objetivo del presente trabajo fue determinar el efecto de la infestación de Tillandsia recurvata L. (Bromeliaceae) sobre el éxito reproductivo (número de flores capaces de producir semillas) de Fouquieria splendens Engelm. (Fouquieriaceae). Se espera que mientras mayor sea la infestación, disminuya el éxito reproductivo de F. splendens.
Este estudio se realizó en un área de 0.5 ha dentro de un matorral micrófilo–espinoso cerca de Peña Blanca, en el estado de Querétaro, próximo a la Reserva de la Biosfera Sierra Gorda, localizada a los 20° 55' 23.6'' N, 99° 43' 52.4'' O, con una altitud de 1591 m. La precipitación en el área oscila entre 280 y 400 mm y la temperatura anual varía de 18 a 22°C.
Las especies más frecuentes son Acacia perniciosa L., Condalia mexicana Schlecht., Fouquieria splendens (ocotillo), Koebeñinia spinosa Zuce, Larrea tridentata (Sessé & Moc. ex DC.) Coville. (gobernadora) y Prosopis laevigata (Humb. & Bonpl. ex Willd.) M.C. Johnston (mezquite) entre otras (Rzedowski, 1981; Zamudio, 1984).
Fouquieria splendens (Fouquieriaceae) es un arbusto resinoso de 2 a 6 m de alto, con abundantes ramas delgadas, espinosas y extendidas desde la base; tiene hojas pequeñas de 2 a 3 cm de largo, éstas son caedizas en época de lluvias. Los tallos son fotosintéticos y tienen el metabolismo típico de las crasuláceas (Rzedowski, 1981; Arreguín–Sánchez et al., 1997). Posee flores rojas o anaranjadas agrupadas en panículas que raramente son blancas (Bowers, 2005). Esta especie se distribuye desde el norte del país hasta Querétaro; habita zonas semi–desérticas dominadas por matorral xerófilo y es abundante sobre suelos rocosos de origen calizo (Miranda y Hernández, 1963; Arreguín–Sánchez et al., 1997).
Tillandsia recurvata es una planta epífita de 8 cm de radio, agrupada por lo general en varias masas esféricas con un sistema rudimentario de raíces (Madison, 1977). Suele colonizar árboles y cables telefónicos Posee tricomas foliares higroscópicos que le permiten condensar la humedad del viento y así absorber nutrimentos (Rzedowski, 1981). No ataca el sistema vascular de sus forofitos, sólo los utiliza como soporte y tiene fotosíntesis tipo C3 (Smith et al., 1986). Las semillas son fusiformes, muy pequeñas y poseen vellos blancos y sedosos que le facilitan su adherencia a cualquier superficie (Miranda y Hernández, 1963).
En abril de 2005 se seleccionaron al azar 66 individuos de F. splendens. Se les midió la cobertura (CobF) mediante la fórmula propuesta por Kennet (1973) y Southwood (1978) (ecuación 1):
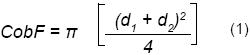
Donde:
d1 = diámetro mayor (cm)
d2 = diámetro perpendicular a d1 (cm)
A cada individuo de F. splendens se le identificó el número de panículas, así como las flores totales por panícula (Ni); de éstas se eligieron tres al azar y en ellas se contabilizó el número de flores muertas (Nc) consideradas como no fecundadas y que se reconocieron por dejar una cicatriz en la panícula. Posteriormente se contaron las flores con frutos (Nf). Además, se registraron las agregaciones de T. recurvata (NT), de las cuales se eligieron al azar tres, a las que se les midió la cobertura (CobT) mediante la ecuación 2:
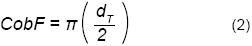
Donde:
dT = diámetro de T. recurvata (cm) a partir de su follaje.
La infestación de T. recurvata sobre F. splendens (It) se calculó con la ecuación 3:

Donde:
CobT = cobertura de T. recurvata (cm2)
NT = número de agregaciones de T. recurvata
CobF = cobertura de F. splendens (cm2)
La proporción de flores muertas (Pe) se calculó con la ecuación 4:
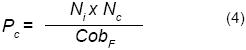
Donde:
Ni = número total de flores por panícula de F. splendens
Nc = número de flores muertas por panícula de F. splendens
Con la ecuación 5 se calculó el potencial reproductivo de F. splendens (Pf):
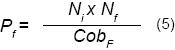
Donde:
Nf = número de flores con fruto por panícula de F. splendens
La proporción de frutos producidos (P) fue calculada con la ecuación 6:

Donde
N = proporción de las flores con semillas
(Nf) = ponderadas en función de las flores muertas
(Nc) = calculada con la ecuación 7:

Se hicieron análisis de correlación entre It vs Pf, It vs Pce It vs Ps.
De F. splendens, la CobF fue 5998.7 ± 15.2 cm2 (promedio ± error estándar). Se contaron 396 panículas; cada individuo tuvo en promedio 6.6 ± 1.5 panículas. Cada panícula estuvo integrada por 25 ± 3.5 flores (mínimo 21 flores, máximo 35 flores). Se encontraron 175 cicatrices de flores desprendidas y 127 flores con fruto (n = 198 panículas).
Se registraron 315 agregaciones de T. recurvata sobre F. splendens. Su CobT fue 16.5 ± 5.3 cm2 (n = 198 agregaciones). La infestación de T. recurvata sobre F. splendens (It) fue de 0.866 (casi 87%). La proporción de flores muertas (Pc) de splendens fue de 11.55 flores/cm2. La proporción de frutos producidos (Ps) por splendens alcanzó 0.048 frutos/cm2. Por su parte, se estimó un potencial reproductivo (Pf) de F. splendens de 8.42 flores con semilla por panícula/cm2.
Se verificó una correlación positiva y significativa entre la infestación de T. recurvata y el potencial reproductivo (Pf) de F. splendens (r63 = 0.31, P = 0.014) (Figura 1). La fracción de varianza (r2) explicada por esta correlación fue de 9.37%.

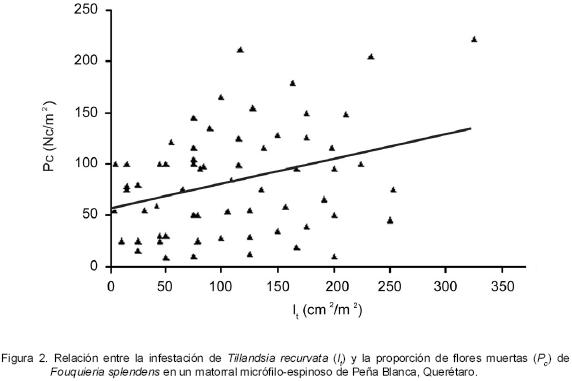
Existe una correlación positiva y significativa entre la infestación de T. recurvata (It) y la producción de flores muertas (Pc) (r63 = 0.33, P = 0.007) (Figura 2). La fracción de varianza (r2) explicada por esta correlación fue de 11.03%.
Se registró una correlación no significativa entre la infestación de T. recurvata (It) y la proporción de frutos producidos (Ps) por m2 de cobertura (r63 = 0.05, P = 0.71) (Figura 3). En este caso, la fracción de varianza explicada (r2) fue de 0.22%.

La infestación de T. recurvata sobre F. splendens estuvo correlacionada directamente con el número de flores no exitosas producidas por unidad de área. Este hecho sugiere que la presencia de T. recurvata tuvo un efecto negativo sobre el forofito, ya que reduce las posibilidades de éxito para la producción de frutos.
Los resultados obtenidos revelaron la existencia de una alta proporción de flores muertas (Pc) (ca. 11.55 flores/cm2 de cobertura de F. splendens), así como también una muy baja proporción de frutos producidos (Ps), pues este valor apenas alcanzó 0.048 frutos/cm2 de cobertura de F. splendens. Estos resultados pueden deberse a los efectos indirectos que provoca la presencia de T. recurvata sobre su planta hospedera como son la reducción de la captación de luz solar y el incremento de la mortalidad de tejidos (Valdivia, 1977; Stevens, 1987; Lüttge, 1989; Richards, 1995).
Lo anterior sugiere la posible existencia de un fenómeno de compensación por parte del forofito, ya que al estar más infestado por T. recurvata, el potencial reproductivo (Pf) también se incrementó de manera proporcional (Figura 1). Al adoptar esta estrategia, es posible que F. splendens logre asegurar que se fecunde la misma cantidad de óvulos que si no estuviera infestada. Los estudios en otras especies acerca de esta forma de compensación han mostrado que las áreas foliares no dañadas son capaces de acelerar la maduración de nuevas hojas, de manera que puedan absorber grandes cantidades de carbono y así destinar los recursos fotosintéticos a las estructuras reproductoras (Lehtila y Syrjänen, 1995; Kaitaniemi y Honkanen, 1996; Thompson et al., 2003).
En este estudio, la reducción del área fotosintética fue causada por la infestación de T. recurvata y, aunque no se midió directamente la tasa fotosintética de F. splendens, se puede intuir que los recursos obtenidos por el arbusto fueron asignados a una producción adicional de flores para compensar la proporción de las que no fueron exitosas (Figura 2).
Por su parte, la infestación de T. recurvata y la proporción de frutos producidos por unidad de cobertura (Figura 3) no estuvieron correlacionadas. Este resultado refeja un patrón muy similar al reportado por Bowers (2005) quien demostró que el tamaño de la panícula (considerado como el promedio de flores con semillas) no estuvo correlacionado significativamente con el tamaño de la planta, ni con el número de las ramas con flores.
Según esta autora, el tamaño de las panículas es un estimador confiable de la cantidad de frutos que el forofito produce por unidad de cobertura. La cantidad de flores y panículas que F. splendens produce puede ser afectada por la humedad edáfica y atmosférica que prevalecen en la localidad. Los datos de su investigación para un año seco mostraron la presencia de muchas panículas con pocas flores; mientras que en un año húmedo consecutivo, se encontraron menos panículas pero con muchas más flores por panícula. En su discusión, incluye que aunque las flores parecieron normalmente abundantes en el año con más lluvia, el promedio de flores por rama en realidad fue 39% menor que en relación con el año seco. El porcentaje de frutos producidos por panícula durante el año húmedo fue 36% más del doble registrado durante el año seco. Estos datos reflejan que la producción de flores por rama fue suficiente durante el año seco para compensar la baja producción de frutos en ese año, lo que resultó en casi la misma proporción para el siguiente ciclo húmedo.
Se requieren estudios más detallados para precisar el efecto que tiene la humedad en el éxito del establecimiento, crecimiento y reproducción de T. recurvata sobre F. splendens, así como determinar el de la humedad sobre el número de frutos producidos por cada rama, su calidad en términos de biomasa y su capacidad germinativa.
AGRADECIMIENTOS
Los autores manifiestan su más sincero agradecimiento a los miembros del grupo 5175 de Ecología I (semestre 2005–II) por su invaluable ayuda en el trabajo de campo, así como a Marco A. Romero por el apoyo técnico y a José G. García Franco por el apoyo bibliográfico. Dos revisores anónimos mejoraron el manuscrito.
REFERENCIAS
Arreguín–Sánchez M. L., L. G. Cabrera, N. R. Fernández, L. C. Orozco, C. B. Rodríguez y B. M. Yepez. 1997. Introducción a la flora del Estado de Querétaro. Consejo de Ciencia y Tecnología del Estado de Querétaro (CONCYTEQ). Santiago de Querétaro, Qro. México. 361 p. [ Links ]
Benzing, D. H. 1990. Vascular epiphytes: general biology and related biota. Cambridge University Press. Cambridge, MA. USA. 690 p. [ Links ]
Bowers, J. E. 2005. Infuence of plant size and climatic variability on the floral biology of Fouquieria splendens (ocotillo). Madroño 52: 158–165. [ Links ]
Kaitaniemi, P. and T. Honkanen. 1996. Simulating source–sink control of carbon and nutrient translocation in a modular plant. Ecological Modelling 88: 227–240. [ Links ]
Kennet, A. K. 1973. Quantitative and dynamic plant ecology. Edward Arnold Ltd. London, UK. 309 p. [ Links ]
Lehtila, K. and K. Syrjänen. 1995. Compensatory responses of two Melampyrum species after damage. Functional Ecology 9: 511–517. [ Links ]
Lüttge, U. 1989. Vascular plants and epiphytes: evolution and ecophisiology. Nature Wissenschaften 72: 557–566. [ Links ]
Madison, M. 1977. Vascular epiphytes: their systematic occurrence and salient features. Selbyana 2: 1–13. [ Links ]
Miranda, F. y E. Hernández X. 1963. Los tipos de vegetación de México y su clasificación. Bol. Soc. Bot. Méx. 28: 29–179. [ Links ]
Reyes–García, C. 2001. Ecofisiología de epífitas de selva baja caducifolia del genero Tillandsia (Bromeliaceae): estacionalidad y fotosíntesis. Tesis de Licenciatura. Facultad de Ciencias, Universidad Nacional Autónoma de México. México, D. F. México. 81 p. [ Links ]
Rzedowski, J. 1981. Vegetación de México. Editorial Limusa. México D. F. México. 430 p. [ Links ]
Richards, P. W. 1995. Tropical rain forest: an ecological study. Cambridge University. Press, Cambridge, MA. USA. 670 p. [ Links ]
Smith, J. A. C., H. Griffiths and U. Lüttge. 1986. Comparative ecophysiology of CAM and C3 Bromeliads: the ecology of the Bromeliaceae in Trinidad. Plant Cell and Environment 9: 359–376. [ Links ]
Southwood, T. R. E. 1978. Ecological Methods. Chapman and Hall. London, UK. 255 p. [ Links ]
Stevens, G. S. 1987. Lianas as structural parasites: the Bursera simaruba example. Ecology 68: 77–81. [ Links ]
Thomson, V. P., S. A. Cunningham, M. C. Ball and A. B. Nicotra. 2003. Compensation for herbivory by Cucumis sativus through increased photosynthetic capacity and effciency. Oecologia 134: 167–175. [ Links ]
Valdivia, E. P. 1977. Las epífitas. Estudio botánico y ecológico de la región del Río Uxpanapa, Veracruz. Biotica 2: 55–71. [ Links ]
Zamudio R., S. 1984. La vegetación de la cuenca del río Estórax en el estado de Querétaro y sus relaciones fitogeográficas. Tesis de Licenciatura. Facultad de Ciencias, Universidad Nacional Autónoma de México. México, D. F. México. 275 p. [ Links ]

