











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
Beetles represent a major component of terrestrial and also freshwater ecosystems. Their ecological roles are manifold and are often highly differentiated between adults and larvae. Not surprisingly, many beetles, adults and larvae, fulfil highly specialised ecological functions in modern ecosystems. We can assume that this was already similar in the past.
Yet, certain assumptions of past ecosystems are more reliable than others. It seems common to conclude from the presence of a specific adult morphology at a specific time slice to the contemporary presence of a specific larval morphology. However, as demonstrated in other lineages of Euarthropoda, such an assumption can be highly misleading (Baranovet al., 2019). Especially in other crustacean groups (Insecta is an ingroup of Crustacea sensu lato,e.g.discussion in Haug and Haug, 2015) it could be demonstrated that larvae and adults have a certain evolutionary independence. This independence can lead to the phenomenon that modern appearing adults are already present, while the specialised larvae evolved much later, or vice versa, with larval morphologies preceding specialised adult morphologies (Hauget al.,2015). Also phylogenetic inferences may be misleading in this aspect, as convergence in closely related lineages may lead to very similar morphologies and developmental patterns (Hauget al., 2013a).
For more reliably reconstructing ecological interactions or even entire ecosystems in the past, it is therefore crucial to indeed look for fossils of larvae, including beetle larvae. Yet, not all types of lifestyles will make it readily likely that such a larva becomes preserved as a fossil. Larvae associated with resin-producing trees have, for example, a higher chance to be preserved in resin. This makes it easy to understand why larvae living directly on such trees are preserved in amber.
Nevertheless, also more unusual larvae such as aquatic ones have been found in various types of ambers. While there have been quite some different interpretations of the occurrence of aquatic organisms in amber (e.g.Wichardet al., 2009) it seems now to be more widely accepted that such organisms indeed became trapped in the amber within their natural habitat (e.g.discussion in Schädelet al., 2019 and references therein). Specialised larvae associated with running waters, but not necessarily living within in them, seem even less likely to be preserved in amber.
We here report a small-sized beetle larva that shows characters of the group Heteroceridae. Extant larvae of this group are known to live in sandy shores of, for example, rivers. We discuss indications of this new find.
2. Material and Methods
Material:In the centre of this study is a single specimen preserved in early Miocene (Aquitanian) Chiapas amber, Mexico (Serrano-Sánchezet al. 2015). The specimen was legally purchased from the amber trader gemgazer.com. The specimen is now deposited in the Palaeo-Evo-Devo Research Group Collection of Arthropods, Ludwig-Maximilians-Universität München, Germany, under repository number PED 0895.
Microscopic documentation:The specimen was initially documented on a Keyence VHX-6000 digital microscope, following standard procedures (e.g., Hauget al., 2013b; Haug and Haug, 2019). This includes composite imaging in x-, y- and z-axis (Hauget al., 2011) as well as HDR (Hauget al., 2013c).
X-ray micro-computed tomography (µCT): The specimen was further documented with a XRadia MicroXCT-200 (Carl Zeiss Microscopy GmbH, Jena, Germany), which is equipped with switchable scintillator-lens units. 1600 projections were recorded with following settings: 10x objective; source settings 40 kV, 8 W; 6s exposure time; binning 2. System-based calculated pixel size is 1.43 µm, 1005 x 1005 px. Projections were further processed with binning 1 using the XMReconstructor software (Carl Zeiss Microscopy GmbH, Jena, Germany), resulting in image stacks (TIFF format). Volume renderings based on the µCT data were processed using OSIRIX.
3. Description of the specimen
General:Small-sized beetle larva with total body length of ~1.61 mm. Body elongate cylindrical, only slightly dorso-ventrally flattened (Figure 1). Organised in anterior head and elongate trunk (Figure 2). Trunk further differentiated into anterior (thorax) and posterior region (abdomen) (Figure 2). Head slightly flattened dorso-f2ventrally, prognathous (mouth parts facing forwards) (Figures 2C to 2I). Antennae discernible, mouth parts prominent. Anterior trunk with three segments (pro-, meso-, metathorax). Thorax with one pair of appendages (legs) on each segment (Figure 1). Posterior trunk, abdomen, with ten discernible units, nine segments and terminal end (possibly compound structure; Figure 1). Abdomen segment 9 with posteriorly drawn-out tergite (Figure 3). Terminal end elongate, orientated postero-ventrally, not discernible in dorsal view (Figures 1A-1C, 3B-3E).
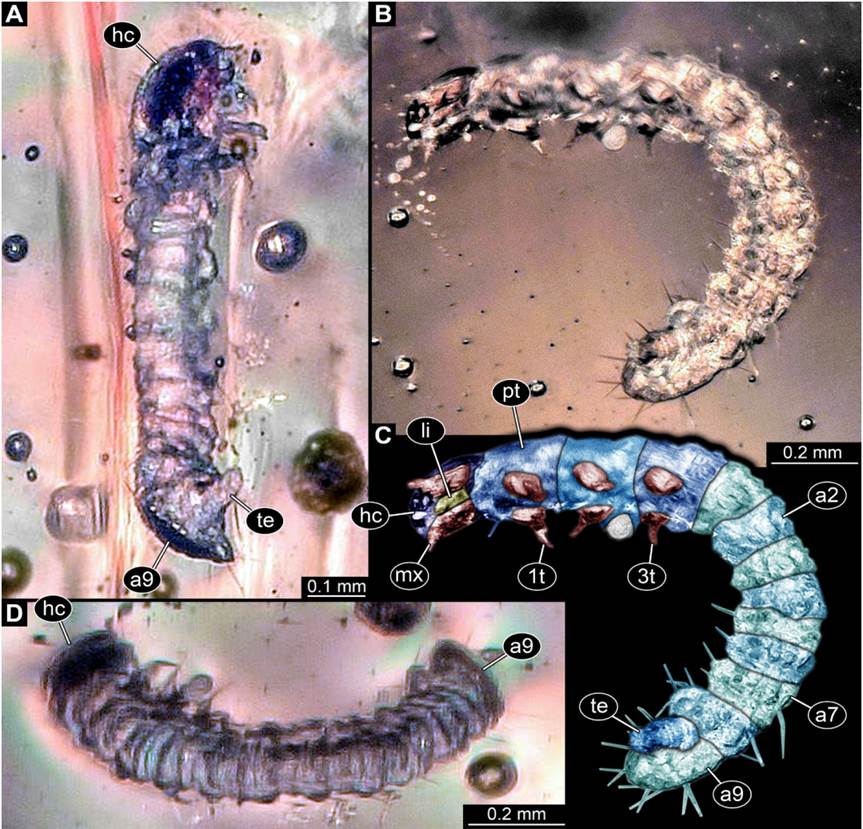
Figure 1 New beetle larva from Mexican amber, PED 0895, light microscopy. A. Oblique ventral view. B. Oblique lateral view. C. Colour-marked version of B. D. Oblique dorsal view. Abbreviations: 1t-3t= trunk appendages (“legs”) 1-3; a2-a9 = abdomen segments 2-9; hc = head capsule; li = labium; mx = maxilla; pt = prothorax; te = trunk end.

Figure 2 New beetle larva from Mexican amber, PED 0895, volume renderings based on µCT. A-D. Red-cyan stereo anaglyphs. A, B. Overviews. A. Oblique dorsal view. B. Oblique lateral view. C. Anterior body, ventral view. D. Antero-dorsal view on head. E. Colour-marked version of D. F-H. Red-cyan stereo anaglyphs of head region. F. In dorsal view. G. In antero-ventral view. H. In ventral view. I. Colour-marked version of H. J. Red-cyan stereo anaglyph of anterior body region in postero-ventral view. Please use red-cyan glasses to view stereo anaglyphs. Images not to scale. Abbreviations: at = antenna; hc = head capsule; li = labium; lr = labrum; md = mandible; mp = maxillary palp; mx = maxilla.

Figure 3 New beetle larva from Mexican amber, PED 0895, volume renderings based on µCT, continued. A, B. Red-cyan stereo anaglyphs. A. Overview in oblique lateral view. B. Posterior trunk region in lateral view. C. Colour-marked version of B. D. Red-cyan stereo anaglyph of posterior trunk region in posterior view. E. Colour-marked version of D. Please use red-cyan glasses to view stereo anaglyphs. Images not to scale. Abbreviations: a9 = abdomen segment 9; te = trunk end.
Head:Almost circular in dorsal view, flattened in lateral view (Figures 2A, 2B and 3A), as long as wide (~0.13 mm), dorsal surface with Y-shaped suture (ecdysis lines) (Figure 2F). No stemmata discernible. Antenna (appendage of post-ocular segment 1) short, with two discernible elements (Figure 2E and 2I), ~0.03 mm long. Second element with a process almost as long as main part of element (Figure 2E).
Mouth parts discernible (Figure 2). Labrum (derivative of ocular segment) relatively large, sub-rectangular in antero-dorsal view (Figure 2E). No spines or setae discernible.
Mandibles (appendages of post-ocular segment 3) partially accessible (Figures 2E, 2I), yet without details.
Maxillae (appendages of post-ocular segment 4) elongate, subrectangular in ventral view (Figure 2I), longer than wide, 2.5x (~0.15 mm long). Proximally slightly wider due to medially drawn-out part, distally with palp (~0.03 mm long), presumably with multiple elements.
Labium (conjoined appendages of post-ocular segment 5) elongate, triangular in ventral view (Figure 2I), longer than wide, 3x (~0.09 mm long), distally with a pair of palps (Figure 2I).
Thorax:Prothorax sub-trapezoidal in dorsal view, anterior rim slightly wider than posterior rim, lateral sides rounded (Figure 2A), as long as wide at the posterior rim (~0.19 mm). Tergite of prothorax (pronotum) with prominent median longitudinal line, lateral sides prolonged and drawn-down ventrally. Single seta discernible (Figures 1B,1C).
Mesothorax sub-rectangular in dorsal view, slightly shorter than prothorax (~0.18 mm), as wide as the posterior rim of prothorax (Figure 2A).
Metathorax sub-similar to mesothorax, shorter than prothorax, (~0.16 mm), as wide as the posterior rim of prothorax (Figure 2A). Tergites of meso- and metathorax with transverse lines.
Legs discernible on all three segments, ~0.11 mm long. Yet, details, ase.g.exact number of elements not accessible (Figures 1A-1C, 2C and 2J).
Abdomen:Abdomen segments 1-8 subsimilar (Figures 1B to 1D), wider than long, measuring between 0.06-0.11 mm in length (with segment 5 being shortest and segments 1 and 8 longest) and between 0.19-0.22 mm in width (with segment 8 being widest). Lateral sides of tergites of segments 1-8 prolonged and drawn-down ventrally (Figures 2A and 2B).
Abdomen segment 9 semi-ovoid in ventral and dorsal view, longer than wide, 1.4x (~0.22 mm long), tergite drawn-out posteriorly, longer than ventral region of segment 3.1x (ventral region ~0.07 mm long). Few setae discernible on abdomen segments 5-9, no exact arrangement (chaetotaxy) reconstructible.
Terminal end longer than wide, 2.3x (~0.16 mm long) hidden by posteriorly drawn-out part of tergite of segment 9 in dorsal view. Ventrally wider, tapering distally (Figure 3B and 3C). Distal part ending in two symmetrical, short processes (Figures 3D and 3E).
4. Discussion
Beetle larvae preserved in amber:Klausnitzer (2003) assumed that beetle larvae are rare in (Baltic) amber and difficult to interpret. This evaluation seems to be based on the fact that beetle larvae are only rarely reported as fossils, leaving the impression that they are rare. Yet, fossil beetle larvae may in fact be much more common, but only rarely investigated and published.
For other groups of Holometabola it could be well demonstrated that the seeming rareness of larvae in amber is simply an effect of low sampling and especially non-specific sampling. Aimed search for specific larvae revealed that some of the presumed rare forms are more common than originally expected (Neuroptera:e.g., Pérez-de la Fuenteet al., 2020; Hauget al.,2020a, 2021a; Diptera:e.g., Baranovet al., 2019, 2020, 2021).
Why is it worth to search for fossil larvae at all? Numerous reasons have been given why the focus on adults is in fact not based on logical reasons but a convention, and why larvae add to research in various ways (see, for example, discussions in Haug and Haug 2016, 2017 and references therein). Especially in holometabolan groups the often strongly differing larvae with their usually also strongly differing ecology can be taken as prime examples. The relative evolutionary independence of adults and larvae (Scholtz, 2005) is so strongly expressed in beetles, butterflies and their relatives that ignoring their immatures, especially in the fossil record, is not comprehensible.
The practical challenges of dealing with beetle larvae in amber are another part of the story. Most features that a determination key for extant beetle larvae requires are not accessible in fossils. Klausnitzer (2003) suggested that only larvae with very pronounced features can be properly determined. It appears indeed easier to recognise very special larvae such as, for example, the larvae of texas beetles (Brachypsectridae) with their specific head and a trunk with very unusual and complex lateral processes (Zhaoet al., 2020; Hauget al., 2021b). Also larvae of wriggling beetles (Gyrinidae; Zhaoet al.,2019; Gustafsonet al., 2020) or certain larvae of false flower beetles (Scraptiidae; Haug and Haug, 2019; Zippelet al., 2022) with enlarged trunk ends can be easily identified as such based on prominent structures. Another problem occurs with larvae that are very special in appearance, but unknown in the modern fauna (Hauget al.,2020b). This lack of knowledge does not mean that such larvae are not present in the modern fauna, as the immature stages of the majority of modern beetles (and many other holometabolans) is still unknown.
Especially in very small beetle larvae it can be challenging to provide any guess of their identity as even fewer details are often accessible. Access with optical methods is limited in amber due to the optical properties of the amber, usually meaning that the lower border of resolution is way above the Abbe limit. This phenomenon is mainly caused by irregularities of the amber, resulting in refractive artefacts. Also modern µCT imaging does not necessarily improve this specific aspect, since depending on the preservation of the specimen, the x-ray absorption of inclusion and surrounding amber can be very similar (resulting in low contrast), and metallic particles can cause strong artefacts. This is especially problematic with small specimens in amber, however, µCT is still able to improve access to concealed and verlumt details. Luckily, the specimen at hand has some features which can be quite informative.
Comparison of the fossil to extant beetle larvae:The posterior end of the fossil larva is quite prominent, with the trunk end being cone-like, protruding from abdomen segment 9 postero-ventrally. Such an arrangement is well known in a number of beetle larvae: For example, in at least some larvae of the groups Lymexilidae (Wheeler, 1991, Figure 34.498-34.500 p. 447), Cleridae (Foster and Lawrence, 1991, Figure 34.508 p. 451), Nitidulidae (Lawrence, 1991a, Figure 34.514a p. 458), Rhizophagidae (Lawrence, 1991b, Figure 34.523a p. 461), Cryptophagidae (Lawrence, 1991c, Figure 34.539 p. 470), Languriidae (Lawrence, 1991d, Figure 34.546 p. 472), Erotylidae (Lawrence 1991e, Figure 34.548 p. 474), Melandryidae (Lawrence, 1991f, 34.643a p. 507) and Elateridae (Becker, 1991, Figures 34.437a, 34.439, 34.440 all on p. 415) we find an overall similar arrangement. Yet, in all these cases abdomen segment 9 bears prominent processes or urogomphi, which are clearly not present in the larva at hand. There is also no indication that such structures were originally present in the fossil, but broke off.
The similarity to certain larvae of Elateridae is even higher concerning the shape of abdomen segment 9, at least in dorsal view. (e.g.Becker, 1991, Figure 34.436 p. 415, Figures 34.450, 34.451 both on p. 417). Yet, these larvae differ in other aspects. In lateral or ventral view, abdomen segment 9 differs from that of the new larva in lacking a distinct ventral region. Also larvae of Elateridae have a special arrangement of the moulting sutures on the head capsule, with a vase-shaped middle part (e.g.Becker, 1991, Figure 34.437b p. 415). In the fossil, the sutures form a simple Y.
A more distantly similar morphology of the trunk end is also known in few larvae of Tenebrionidae (Lawrence and Spilman, 1991, Figure 34.672a, d p. 525). However, in these larvae the trunk end appears quite differentiated, while in the fossil it appears rather simple, soft and tubular. Such a pygopod-like morphology of a trunk end can also be seen in some larvae of Chrysomelidae (Lawson, 1991,e.g.Figure 34.835 p. 582), yet these differ significantly in overall appearance compared to the state in the new fossil.
Distantly similar trunk ends are also present in larvae of Byrrhoidae (Lawrence, 1991f, Figure 34.387-389 p. 385), Ptilodactylidae (Lawrence, 1991g, Figure 34.403 p. 392) and Artematopidae (Cooper, 1991, Figure 34.433a p. 408). Yet, the largest similarity concerning the shape of the trunk end is in fact seen in larvae of Heteroceridae. Here the trunk end is distinctly pygopod-like (e.g.Vaninet al., 1995, Figures 11, 12 both on p. 107).
Also other characters are compatible with an interpretation of the fossil larva as representative of the group Heteroceridae. The moulting sutures of the head capsule are simple and Y-shaped in extant larvae, yet, unlike in the fossil, the middle branch is much shorter. The antennae in larvae of Heteroceridae are very small (Lawrence 1991h p. 402; Vaninet al., 1995, Figure 15 p. 108, Figure 19 p. 109) and are distally bifid (Vaninet al., 1995, Figure 15 p. 108), which seems also to be the case in the new fossil (Figures 3D and 3E). Also the prominent maxillo-labial complex is very similar in the modern larvae and the fossil with short maxillary palps, very broad proximal (posterior) end of the labium and very short labial palps.
There are certain differences of the fossil larvae, as for example the rather wide abdomen segment 9, which is about as wide as the head capsule. In the known larvae of Heteroceridae, abdomen segment 9 is always narrower than the head capsule, but in fact only very few larvae of Heteroceridae are known (examples in Figure 4). Moreover, all known larvae seem to be late stage larvae, while the fossil, based on its small size, is likely an early stage larva. The phenomenon that mostly late stage larvae are known in the modern fauna, but only early stage larvae are known as fossils, has been noticed in at least one other beetle group (Scraptiidae), and the resulting problems have been discussed (Haug and Haug, 2019; Zippelet al., 2022).

Figure 4 Examples of extant larvae of Heteroceridae, modified from the literature. A-D. From Pierre (1945). A. His figure 1. B. His figure 18. C. His figure 19. D. His figure 20. E. From Vanin et al., (1995), their figure 10.
Certain morphological aspects of the fossil larva, which could further support an interpretation as a larva of Heterocerdiae, such as scattered stemmata, details of the mandibles or specialised spiracles (Lawrence, 1991h, pp. 402-403) are not accessible in the fossil.
We can therefore only conclude that an interpretation of the larva as representative of Heteroceridae is compatible with the observable details and represents the currently best interpretation. Details possibly further corroborating this interpretation are not available.
Ecology of modern-day larvae of Heteroceridae and fossil record of the group:Modern larvae of Heteroceridae live on the sandy shores of water bodies (rivers, ponds, but also bodies of brackish or even salty water) and dig tunnels through the wet sand. They seem to ingest substrate, extracting algae and other micro-organisms as food (e.g, see discussion in Vaninet al., 1995 p. 104; Sazhnev, 2018, 2020).
So far, very few fossils of Heteroceridae have been reported, largely restricted to adult specimens (Ponomarenko, 1986; Prokin and Ren, 2011; Liet al., 2020). The only other record is a supposed trace fossil of tunnels (Clark and Ratcliffe 1989). Yet, as could be shown, very similar traces can be left also by other organisms, making a clear identification challenging (Metz, 1990).
What does this mean for the new larva? We know that the earliest representatives of the group were already present about 100 million years ago (Liet al., 2020), hence it is reasonable to have fossils in 23-million-year-old amber from early Miocene (Serrano-Sánchezet al., 2015). Also the group is represented in the modern fauna of Mexico (e.g.Skalicky, 2018, 2021). Although it has been speculated that aquatic organisms preserved in amber can only be explained by very unusual circumstances (e.g.Wichardet al., 2009) it indeed appears that in certain amber types aquatic organisms are preserved in situ (e.g.Schmidtet al., 2018; Xinget al., 2018; Yanget al., 2019; Zhaoet al.,2019; Gustafsonet al., 2020; Wanget al., 2020; Schädelet al., 2020, 2021a, 2021b; Hauget al., 2021c; Baranovet al., 2022). Also Mexican amber is well known to preserve aquatic organisms (Huyset al., 2016; Serrano-Sánchezet al., 2016; Duet al., 2019; Matzke-Karaszet al., 2019). Hence the preservation of animals living directly associated to water bodies, such as larvae of Heteroceridae, seems well likely. In summary, an interpretation of the new larva as a representative of Heteroceridae, with a similar lifestyle to modern forms, is not contradicted by ecological and preservational circumstances.
Larvae in Chiapas amber:As pointed out, larvae preserved in amber are less often reported than adults. So far only few larvae have been explicitly reported from Chiapas amber (e.g.Poinar and Poinar, 2005; Serrano-Sánchezet al., 2016; Hauget al., 2021d). Yet, apparently such individuals can well be preserved.
It is well possible to use larval forms for comparisons of diversity, if sufficient material can be gathered for different time slices (e.g.Hauget al., 2020a, 2021a). While Mexican amber is not the only source of amber from the Miocene, it is especially important, as it preserves components of the aquatic fauna and animals associated with water (e.g.Solórzano-Kraemer, 2010, Serrano-Sánchezet al., 2015), as presumed for the new larva. Such components appear to be largely absent in Miocene Dominican amber. Hence, Mexican amber is a good source of information for making our view on the Miocene fauna more complete. It will also be important to continue to report more larvae, such as the fossil here, to add ecological roles so far not directly recognised.

