











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
In modern marine benthic environments, brachyurans are well represented by approximately 7,250 species (Davie et al., 2015). The most primitive ones, sometimes controversially discussed, have been recorded from clayey marine strata of Early Jurassic (Pliensbachian) age (Förster, 1986; Schweitzer and Feldmann, 2010d; Haug and Haug, 2014; Krzemińska et al., 2020; Scholz, 2020). Brachyuran records from the Middle Jurassic are still scarce (Krobicki and Zatoń, 2008; Fraaije et al., 2013). In the Late Jurassic, siliceous sponge-microbial magnafacies as well as coeval coral reefs became widespread along the northern margin of the Tethys Ocean (e.g., Leinfelder et al., 1994, 1996). This probably triggered the first great diversification and biogeographical expansion of crabs (Wehner, 1988; Müller et al., 2000; Klompmaker et al., 2013). The Upper Jurassic of southern Germany, which was part of this northern Tethyan reef belt, is not only a classical area for early brachyuran studies (von Meyer, 1842, 1860) but was probably also a diversification hotspot of this group, although geographically isolated brachyuran records even come from the western Pacific area (Karasawa and Kato, 2007; Kato et al., 2010; Karasawa and Hirota, 2019). Early records from the southern Tethys (Förster, 1985) are of dubious origination (see Krobicki and Zatoń, 2008).
In the course of an extensive revision and update of the ʻTreatiseʼ volume dedicated to decapod crustaceans (Glaessner, 1969), numerous taxonomic revisions of the original type material from old collections became necessary, and additional taxa based on newly discovered or differently interpreted Jurassic crab material were occasionally added (Feldmann et al., 2006; Schweitzer et al., 2007, 2018; Schweitzer and Feldmann, 2008a, 2008b, 2009a, 2009b, 2009c, 2009d; 2010a, 2010b, 2010c; Franţescu, 2011; Schweigert and Koppka, 2011; Starzyk et al., 2012; Hyžný et al., 2015; Starzyk, 2015a, 2015b, 2020; Schweigert and Robins, 2016; Schweigert, 2019; Klompmaker et al., 2020).
The aim of this study is to report a new genus and species of the Prosopidae. The abbreviation SMNS is used for Staatliches Museum für Naturkunde Stuttgart, Germany.
2. Geological setting
The herein described specimen was collected from thick-bedded limestones of the Treuchtlingen Formation (Niebuhr and Pürner, 2014) quarried in the Petersbuch-Erkertshofen district, which is part of the township of Markt Titting in Bavaria (Germany) (Figure 1). Due to its usage as a building stone these limestones are well-known and traded as “Treuchtlingen Marble”. The limestones of the Treuchtlingen Formation yield abundant ammonites, belemnite rostra, siliceous and calcareous sponges, brachiopods, and echinoids. In contrast, hermatypic corals are totally absent from this lithology. In the finer rock matrix macroscopically visible foraminifera with a whitish wall (“Tubiphytes”) are extremely common and characteristic for this lithology. The ammonite fauna allows a precise dating of these rocks within the late Kimmeridgian (Zeiss, 1964; Bantz, 1970; Schlampp, 2004, 2009). The environmental conditions during which the Treuchtlingen Formation was deposited have been discussed controversially in the past. A very shallow depositional model was suggested by Kott (1989); however, subsequently many arguments, especially the total absence of a typical shallow-water fauna (hermatypic corals, thick-shelled bivalves, large gastropods), have been put forward in favour of greater water depths within a range of ca. 50 to 150 metres, certainly below the storm-weather wave basis (e.g., Leinfelder et al., 1994, 1996; Schmid, 1996; Schmid et al., 2005). The occurring decapod crustacean fauna consists of small-sized crabs (see below), is relatively low-diverse and does not comprise any species otherwise restricted to Upper Jurassic coral reefs (cf. Schweitzer et al., 2018).

Figure 1 Locality of the holotype. Arrow indicates location of the Petersbuch quarry at Markt Titting-Petersbuch, Bavaria, southern Germany. Map based on OpenStreetMap (http://www.openstreetmap.org).
Although numerous amateur collectors are hunting for fossils in the Franconian Treuchtlingen Formation these tiny fossils are not in their focus and they are mostly ignored and overlooked. Here is a list of brachyurans from the Treuchtlingen Formation (compiled after Wehner, 1988, Schweigert and Kuschel, 2018, and own observations; taxonomy modified after Schweitzer et al., 2018):
Eodromites grandis (von Meyer, 1857)
Laeviprosopon laeve (von Meyer, 1857)
Nodoprosopon ornatum (von Meyer, 1842)
Petersbuchia thauckei n. gen., n. sp. (herein)
Pilidromia thiedeae (Schweigert and Koppka, 2011)
Planoprosopon heydeni (von Meyer, 1857)
Tanidromites insignis (von Meyer, 1857)
Tanidromites scheffneraeSchweigert and Koppka, 2011.
Verrucarcinus torosus (von Meyer, 1857)
3. Systematic palaeontology
The classification follows Schweitzer et al. (2010).
Order Decapoda Latreille, 1802
Infraorder Brachyura Linnaeus, 1758
Section Dromiacea De Haan, 1833
Superfamily Homolodromioidea Alcock, 1900
Family Prosopidae von Meyer, 1860
Genus Petersbuchia nov.
Type species: Petersbuchia thauckei n. gen., n. sp., monotypic. Gender is feminine.
Diagnosis: See diagnosis of the type species (monotypic).
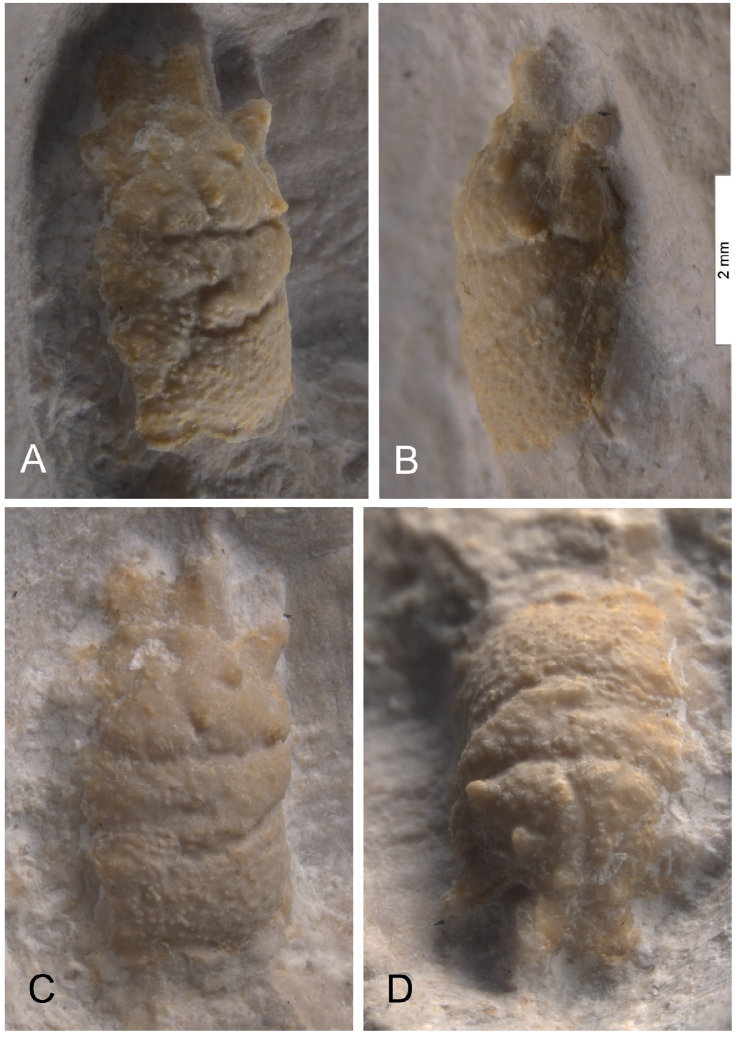
Figure 2 Petersbuchia thauckei n. gen. n. sp., holotype, SMNS 70622, Upper Jurassic, Treuchtlingen Formation, Petersbuch Member; late Kimmeridgian, Acanthicum Zone, Markt Titting-Petersbuch, Bavaria, southern Germany. Dorsal (A, C), lateral (B) and frontal (D) views. A with artificially enhanced contrast. Scale bar = 2 mm.

Figure 3 Petersbuchia thauckei n. gen. n. sp., holotype, SMNS 70622, with a fragmentary chela possibly belonging to the same individual (inset, close-up view). Scale bar = 2 mm.
Etymology: Named after Thomas Haucke (Freystadt-Mörsdorf), who collected and donated the sole specimen.
Holotype: Specimen SMNS 70622 (Figure 2).
Type locality: Quarry area of Petersbuch/ Erkertshofen, Markt Titting, Franconian Alb, Germany (Figure 1).
Type horizon: Treuchtlingen Formation, Petersbuch Member (Upper Jurassic, late Kimmeridgian, Acanthicum Zone; Niebuhr and Pürner, 2014). Exact bed not traceable, because the rock with the fossil was found ex-situ on top of a dump close to a quarry.
Studied material: Holotype only.
Occurrence: At present only known from the type locality.
Diagnosis: Carapace subquadrate, with long protrusive outer orbital spine; Surface coarsely pustulose, anterior part with a few prominent tubercles. Rostrum trifid, with spatulate margins. Hepatic and gastric areas not separated by grooves.
Description: The small-sized holotype SMNS 70622 (Figure 2) is represented by the three-dimensionally preserved inner cast of the carapace. The left side of the carapace is partly lost prior to burial due to breakage. Surface of carapace densely pustulose in all areas. Outline of carapace subquadrate, with broad trifid rostrum. Total length of carapace excluding rostrum 3.7 mm, including rostrum 4.6 mm; reconstructed width by mirroring in symmetry plane 3.6 mm, maximum height 2.1 mm. Trifid rostrum with spatulate upturned margins and downturned tip in the centre. Rounded orbital margin connects base of rostrum and protrusive outer orbital spine. Frontal aspects of orbital area hidden. Hepatic, protogastric and epigastric areas not separated from each other. Mesogastric area well separated by lateral grooves, with two coarser tubercles in its symmetry plane. Posterior gastric muscle scars with subparallel, spring-like ornamentation. In addition to the general pustulose ornamentation, four strong and prominent tubercles are developed in the anterior part of the carapace, one close to the mesogastric groove, another one in the outer third of the hepatic region, close to the cervical groove, the third one near the centre of the joint hepatic-protogastric region and the last one in the epigastric region, at the base of the rostrum. Subhepatic region not separated from hepatic region but posteriorwards forming a lobe. Cervical groove and branchiocardiac groove well developed. Epi- and mesobranchial regions fused. Meta- and urogastric regions fused as well. Cardiac region triangular, but hardly separated from adjacent branchial region. Outer lateral margin of branchial region parallel to longitudinal axis of carapace. Posterior margin of carapace forming a smooth rim, with concavities placed laterally from the intestinal area. Sternal region not preserved. Close to the specimen, there is an incompletely preserved chela possibly belonging to the same individual (Figure 3). The propodus is twice as long as broad and widens distally. Surface of propodus with similar pustulose ornamentation as in the carapace. Fixed finger thick, smooth, distally broken, but slightly curved imprint points to a sickle-like outline. Free finger lost.
Remarks: It is impossible to state whether the sole small-sized specimen of Petersbuchia thauckei n. gen., n. sp. was an adult or a juvenile.
4. Discussion
At present, Petersbuchia thauckei n. gen. n., sp. from the late Kimmeridgian of Franconia represents the only species assigned to this new genus. In first view, the number and arrangement of prominent tubercles in the hepatic region of Petersbuchia thauckei n. gen., n. sp. is strikingly reminiscent to Laeviprosopon sublaeve (von Meyer, 1857), another very rare taxon reported from Upper Jurassic beds of similar age and lithology.
Some species of Laeviprosopon including the type species exhibit a similar trifid rostrum as in Petersbuchia n. gen. However, Laeviprosopon sublaeve lacks a pustulose ornamentation of the carapace as well as an outer orbital spine. It must be stressed that ornamentation is usually less developed in inner casts than on the cuticle. Therefore, if the cuticle of Petersbuchia n. gen. was preserved it would have been even stronger ornamented. Moreover, in Laeviprosopon sublaeve the anterior part of the carapace exhibits a trapezoidal outline.
The posterior margin of the carapace showing laterally placed concavities is shared with a few other genera safely assigned to Prosopidae (Prosopon von Meyer, 1835 s. str.; EuroprosoponKlompmaker, Starzyk, Fraaije and Schweigert, 2020; ProtuberosaSchweitzer and Feldmann, 2009a), but this character does not occur within species of Laeviprosopon (see Klompmaker et al., 2020). Hence, Petersbuchia n. gen. is well distinct from Laeviprosopon and shows only some superficially convergent characters with this co-occurring genus.

