











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introducción
Los mamuts fueron un componente importante de las faunas terrestres norteamericanas durante el Pleistoceno (2.58 millones de años [Ma] - 10000 años) y se extinguieron en este subcontinente hacia el final de esta época geológica (Kurtén y Anderson, 1980; Lange, 2002; Bell et al., 2004). Numerosas especies nominales se han considerado en el Género Mammuthus; sin embargo, recientemente se reconocen en el continente americano al menos cuatro especies de mamuts, las cuales incluyen Mammuthus meridionalis, M. columbi, M. exilis y M. primigenius (Agenbroad, 2005; Lister, 2017). La primera especie, Mammuthus meridionalis arribó a Norteamérica a través del estrecho de Bering, entre los 1.7 y 1.5 Ma (Agenbroad y Nelson, 2002; Lange, 2002; Bell et al., 2004; Rathbone, 2009) y después extendió su distribución geográfica hacia Canadá, Estados Unidos y el centro de México (Kurtén y Anderson, 1980; Agenbroad y Nelson, 2002; Arroyo-Cabrales et al., 2007).
La especie Mammuthus columbi se diferenció a partir de un inmigrante euroasiático perteneciente tal vez a M. meridionalis o M. trogontherii hace aproximadamente un millón de años (Lister y Bahn, 2007; Lister y Sher, 2015) y estuvo ampliamente distribuida desde las partes más templadas de Alaska y Canadá, a través de los Estados Unidos y México, alcanzando áreas de Centroamérica (Lange, 2002; Lister y Bahn, 2007; Lucas y Alvarado, 2010). En el caso de M. exilis, éste se diferenció a partir de algunos integrantes de M. columbi y se conoce únicamente de la Isla de Santa Rosa, California (Agenbroad, 2003, 2012). Las especies M. columbi (mamut de las praderas) y M. exilis (mamut pigmeo insular) fueron exclusivas del Continente Americano (Agenbroad, 2012).
Finalmente, Mammuthus primigenius llegó a Norteamérica durante el Pleistoceno tardío (≈ 100 Ka) (Lange, 2002; Lister y Bahn, 2007). Este mamut se conoce de regiones templadas y frías localizadas en latitudes altas. En Norteamérica se distribuyó en Alaska, Canadá y la parte más norteña de los Estados Unidos (Kurtén y Anderson, 1980; Lister y Bahn, 2007).
En los yacimientos pleistocénicos de México se conocen materiales fósiles de las especies Mammuthus meridionalis y M. columbi (Arroyo-Cabrales et al., 2007). El registro de M. meridionalis procede de tres localidades de mediados del Pleistoceno en el norte y centro del país, Arizpe, Sonora; El Mezquital, Baja California Sur y Culhuacán, Distrito Federal. Por su parte, la especie M. columbi tiene un amplio registro en la mayor parte del territorio nacional (excepto en Tabasco y la Península de Yucatán) y cuyo material procede de numerosas localidades del Pleistoceno tardío (Arroyo-Cabrales et al., 2007).
Por otra parte, desde hace aproximadamente un par de décadas, la caracterización de hábito alimentario en diversas especies de ungulados tanto fósiles como recientes, se ha convertido en una herramienta para conocer diversos aspectos de la ecología de los organismos, tales como la disponibilidad y repartición de recursos. De igual forma, proporciona evidencia de ciertas condiciones ambientales y, por lo tanto, de la dinámica de los ecosistemas (Retallack, 2007). Una de las técnicas que se utiliza para caracterizar el hábito alimentario es el análisis de microdesgaste, la cual permite conocer el tipo de recursos que consumieron los organismos antes de morir, mediante la evaluación microscópica del desgaste en el esmalte dental producido por el contacto diente - diente (atrición) y diente - alimento (abrasión) (Solounias y Semprebon, 2002).
Existen algunos trabajos en los que se ha aplicado la técnica de microdesgaste en elementos dentales de mamuts de Europa y Estados Unidos (Rivals et al., 2010, 2012, 2015; Semprebon et al., 2016). En particular, el estudio de Rivals et al. (2012) consideró muestras de M. columbi y encontraron un hábito comúnmente pacedor. Los estudios de hábito alimentario mediante isótopos estables, análisis de microdesgaste y de mesodesgaste en muestras referidas a M. columbi procedentes de diversas localidades mexicanas, indican que esta especie tuvo un amplio régimen trófico en México (Pérez-Crespo et al., 2012; Gutiérrez-Bedolla et al., 2016; Carbot-Chanona et al., 2017).
Durante la última década, el trabajo paleontológico en localidades de Hidalgo, Puebla y Oaxaca, permitió recuperar una importante muestra de ejemplares pertenecientes a mamuts. En el presente estudio se describen y caracterizan taxonómicamente los materiales dentales y óseos que integran el conjunto hasta ahora recuperado. Asimismo, se proporciona información del comportamiento alimentario de algunos de los individuos estudiados mediante un análisis de microdesgaste.
2. Área de estudio
El material fósil procede de 18 sitios en tres estados de la República Mexicana, a saber, Hidalgo y Puebla (región centro), así como Oaxaca (región sureste) (Figura 1). La presencia de material fósil perteneciente a Bison en todas las localidades es indicativo de una edad correspondiente al Rancholabreano en la cronología norteamericana de mamíferos (Bell et al., 2004). Los ejemplares de Hidalgo se recuperaron de ocho localidades en el sector suroriental del estado, las cuales incluyen: La Pinta de Vaquerías (HGO-10), 20º 22.32’ N - 98º 33.53’ O; Barranca El Berrendo (HGO-28), 20° 01.20’ N - 98° 37.37’ O; Barranca Jagüey Viejo (HGO-42), 20° 02.31’ N - 98° 35.88’ O; Las Cajas (HGO- 46), 20° 07.31’ N - 98° 58.07’ O; El Barrio (HGO-47), 20° 07.41’ N - 98° 56.02’ O; San Gabriel Azteca (HGO-49), 19° 52.43’ N - 98° 36.39’ O; Singuilucan (HGO-50), 20º 02.63’ N - 98º 32.33’ O; así como un molariforme aislado cuya procedencia se indica como Cd. Sahagún, municipio de Tepeapulco. En términos generales, la unidad portadora consiste en arcilla, limo, arena fina y esporádicos lentes conglomeráticos, formando abanicos aluviales y rellenos de valle (Bravo-Cuevas, 2002).
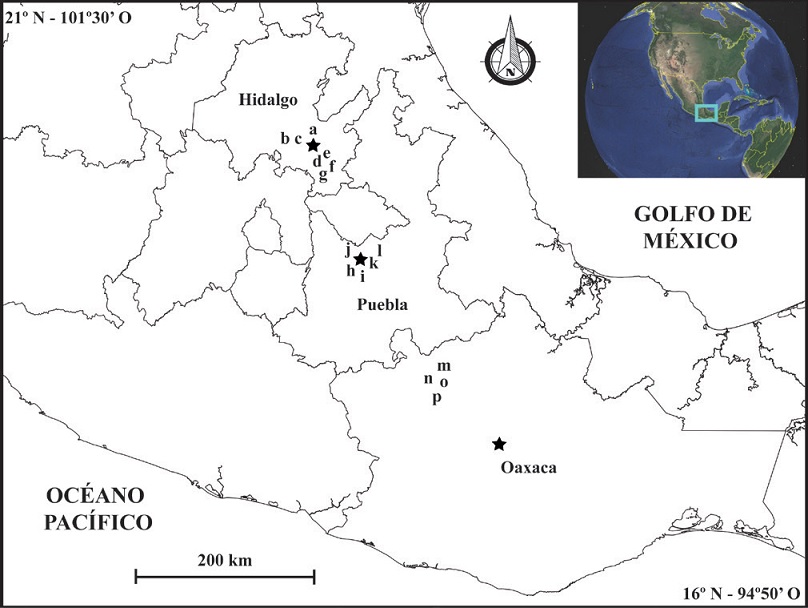
Figura 1 Mapa índice de la ubicación de las localidades en el |centro (Hidalgo y Puebla) y sureste (Oaxaca) de México de las cuales se recuperó el material fósil de Mammuthus columbi considerado en este estudio. Las capitales de cada uno de los estados se indican con una estrella: Pachuca de Soto, Hidalgo; Puebla de Zaragoza, Puebla; Oaxaca de Juárez, Oaxaca. Hidalgo: a. La Pinta de Vaquerías (HGO-10), b. Las Cajas (HGO- 46), c. El Barrio (HGO-47), d. Barranca El Berrendo (HGO-28), e. Barranca Jagüey Viejo (HGO-42), f. Singuilucan (HGO-50), g. San Gabriel Azteca (HGO-49). Puebla: h. Cuahutinchan (PUE-6), i. Barranca Policarpio (PUE-2), j. Dos Cruces (PUE-3), k. Santa Cata (PUE-4), l. Barranca Xocoa (PUE-7). Oaxaca: m. El Pedernal (Oax-2), n. Río Salado (Oax4), o. Llano de Hueso (Oax-5) y p. Río Tejupam (Oax-7).
El material de Puebla procede de las siguientes seis localidades: Barranca Policarpio (PUE-2), 18° 56.40’ N - 98° 07.33’ O; Dos Cruces (PUE-3) 18° 55.00’ N - 98° 10.00’ O; Santa Cata (PUE-4), 18° 55.54’ N - 98° 10’ 15.05” O; Cuahutinchan (PUE-6), 18° 55.54’ N - 98° 10.14’ O; Barranca Xocoa (PUE-7), 18° 57.39’ N - 97° 59.55’ O; así como un molariforme aislado cuya procedencia se indica como Axamilpa. Estos sitios están en los alrededores de la Cuenca de Valsequillo y los depósitos del Cuaternario consisten en sedimentos de origen fluviolacustre con un aporte variable de material volcánico (Ortega-Gutiérrez et al.,1992).
En el estado de Oaxaca, los sitios de donde se recuperó el material de mamuts, se ubican en la Mixteca Alta entre los 17º 35’ - 17º 55’ N y 97º 20’ - 97º 40’ O (Jiménez-Hidalgo et al., 2011), e incluyen a las localidades conocidas como El Pedernal (Oax-2), Río Salado (Oax-4), Llano de Hueso (Oax-5) y Río Tejupam (Oax-7). La unidad portadora consiste en una secuencia de toba o limo arcilloso en la base, mientras que hacia la cima consiste en arena limosa de grano fino a medio con intercalaciones de grava y presencia de rizolitos, así como nódulos de caliche (Jiménez-Hidalgo et al., 2011; Guerrero-Arenas et al., 2013).
3. Materiales y métodos
El material estudiado consistió en un total de 77 ejemplares que incluyen un fragmento de cráneo, tres fragmentos de mandíbula, 26 elementos dentales y 47 elementos del postcráneo. Los ejemplares procedentes de los estados de Hidalgo y Puebla están alojados en la Sección de Macrovertebrados del Museo de Paleontología de la Universidad Autónoma del Estado de Hidalgo (UAHMP), mientras que los ejemplares del estado de Oaxaca están alojados en el Laboratorio de Paleobiología de la Universidad del Mar, Campus Puerto Escondido, Oaxaca (UMPE). El método incluyó la descripción y caracterización taxonómica de los ejemplares, así como un análisis de microdesgaste para conocer el comportamiento alimentario de algunos de los individuos que representan la muestra considerada.
3.1. DESCRIPCIÓN Y CARACTERIZACIÓN TAXONÓMICA DE LOS ELEMENTOS DE MAMUTS
3.1.1. ESTUDIO ANATÓMICO COMPARADO DE LOS EJEMPLARES
La descripción de los materiales fósiles se realizó según el elemento anatómico. En este sentido, los incisivos se midieron de acuerdo con Smith y Fisher (2013). La nomenclatura dental de los molariformes se basa en los sistemas propuestos por Osborn (1942), Saunders (1970) y Madden (1981). En este estudio los elementos inferiores se indican con letras minúsculas y los superiores con mayúsculas (por ejemplo, dp2 = segundo premolar deciduo inferior, DP2 = segundo premolar deciduo superior). Los parámetros configuracionales y métricos evaluados en los elementos dentales son de Maglio (1973), Agenbroad y Brunelle (1992) y Agenbroad (1994).
Los rasgos cualitativos y cuantitativos de la mandíbula se basaron en el esquema de Álvarez-Lao y Méndez (2011). Los trabajos de Olsen (1972), Pasquini et al. (1989), Shoshani (1996) y Sisson y Grossman (2005) se usaron en la descripción de los elementos postcraneales. Por su parte, las características y medidas de las vértebras son de Harington y colaboradores (2012), en el caso de la pelvis se siguió el esquema de Lister (1996), mientras que para los huesos de las extremidades se siguió a Agenbroad (1994) y Hodgson et al. (2008).
Las medidas se tomaron con un calibrador digital (rango de 0-150 mm, resolución de 0.01 mm y precisión de 0.003 mm), un flexómetro de 1.83 m, una cinta métrica flexible de 3 m y una forcípula forestal, según fuese el caso. Todas las medidas están en milímetros (mm). En todos los casos, los elementos postcraneales (fragmentos y/o completos) están asociados a ejemplares cráneo-dentales diagnósticos de mamuts.
Los materiales se compararon con ejemplares seleccionados pertenecientes a las especies Mammuthus meridionalis, M. columbi, M. primigenius y M. trogontherii reportados en la literatura (Maglio, 1973; Dubrovo, 1977; Czyzewska y Wiszniowska, 1982; Pilgram y Western, 1986; Lucas y Effinger, 1991; Gillette y Madsen, 1993; Lucas y González-León, 1996; Morgan et al., 2001; McDaniel y Jefferson, 2003, 2006a, 2006b; García-Zepeda y Garduño-Monroy, 2006; Hodgson et al., 2008; Wei et al., 2010; Álvarez-Lao y Méndez, 2011; Agostini et al., 2012; Albayrak y Lister, 2012; Álvarez-Lao y García, 2012; Athanassiou, 2012; Baygusheva et al., 2012; Harington et al., 2012; Lister et al., 2012; Tong, 2012; Pawlowska et al., 2014). Asimismo, se compararon con elementos de Mammuthus procedentes de la localidad Ingleside, Condado de San Patricio, Pleistoceno tardío de Texas, alojados en el laboratorio de Paleontología de Vertebrados de la Universidad de Texas en Austin; así como con ejemplares alojados en la Universidad Michoacana de San Nicolás de Hidalgo (UM) y el Centro Regional Puebla del Instituto de Antropología e Historia (CRINAH).
3.1.2. ESTIMACIÓN DE EDAD Y SEXO DE LOS EJEMPLARES
La edad de los ejemplares se estimó con base en el estado de desgaste de los molariformes y escala de Craig para estimación de edad en años de elefante africano (Haynes, 1991). Aunado a esto, los segundos y terceros molares superiores (M2 - M3) e inferiores (m2 - m3) completos o fragmentados se clasificaron como pertenecientes a probables machos con base en los criterios propuestos por McDaniel y Jefferson (2006a, tabla 4, p. 174) , los cuales incluyen anchura, frecuencia lamelar (número de lamelas en 10 cm) y grosor del esmalte. Si dos de estos criterios se cumplían se consideró que probablemente el elemento pertenecía a un macho.
3.2. ANÁLISIS DE MICRODESGASTE
La caracterización del hábito alimentario de algunos de los individuos (UAHMP-4220, TCH056, UAHMP-4190, UAHMP-4002, UMPE022, UMPE-029-1, UMPE-029-2, UMPE-506, UMPE-618) que representan parte de la muestra de ejemplares pertenecientes a mamuts, se estableció mediante la evaluación del patrón de desgaste en el esmalte a nivel microscópico. Para tal efecto, se realizó un análisis de microdesgaste de baja magnificación (35X) en 18 bandas de esmalte que representan ocho individuos, de los cuales cinco son de Oaxaca, uno de Puebla y dos de Hidalgo. Se seleccionaron ejemplares que no tuvieran alteración tafonómica. Los moldes y duplicados se elaboraron utilizando material de impresión dental polivinilsiloxano de alta definición y resina epóxica cristalina respectivamente (Solounias y Semprebon, 2002). Las variables convencionales de microdesgaste, rasguños y fosas, se contabilizaron en fotografías preparadas siguiendo el método de Fraser et al. (2009). Las fotografías se tomaron con una cámara digital Nikon D200 acoplada a un microscopio estereoscópico Nikon SMZ1500, mediante el método de Proyección de Imagen de Alto Rango Dinámico (HDRI por sus siglas en inglés: High Dynamic Range Imaging); la resolución digital de las imágenes fue de 0.6 pixeles/μm. En cada foto se delimitaron una o dos áreas de 0.4 x 0.4 mm en las que estuviera representado el microdesgaste dental.
Los conteos de las variables fueron realizados por una sola persona (Elizabeth Ortiz-Caballero) para evitar el error inter-observador (Grine et al., 2002; Mihlbachler et al., 2012). Asimismo, se realizaron de manera aleatoria tres conteos en cada recuadro con el objetivo de minimizar el error de medición y asegurar la “ceguera” del observador (Mihlbachler et al., 2012). Con base en los conteos se obtuvieron los promedios de rayones y fosas, además se establecieron las frecuencias en la textura de los rayones, fosas pequeñas y fosas grandes, así como la presencia o ausencia de gubias.
La nomenclatura y sistema de registro de las variables de microdesgaste son de Solounias y Semprebon (2002) y Semprebon et al. (2004). En las fotografías, los rayones gruesos son menos refractivos y sobresalen como estriaciones oscuras, en tanto que los rayones finos se observan más refractivos. Por su parte, las fosas pequeñas son pequeños círculos oscuros, mientras que las fosas grandes se distinguen por la aparición de sombras oscuras en forma de media luna al lado de la fosa frente a la fuente de luz (Fraser et al., 2009). Las gubias son concavidades de contornos irregulares de dos a tres veces más largas y profundas que las fosas grandes (Solounias y Semprebon, 2002).
Los rasgos de microdesgaste en la muestra estudiada de mamuts se compararon con la base de datos de especies de ungulados con hábito alimentario típico de Solounias y Semprebon (2002), mediante un Análisis de Componentes Principales (ACP) sobre la matriz de correlación. Asimismo, se realizó un Análisis de Funciones Discriminantes sobre el conjunto de datos de Solounias y Semprebon (2002), asumiendo igualdad en las probabilidades de clasificación para todos los grupos, con la finalidad de clasificar la muestra estudiada en alguna de las siguientes categorías alimentarias: ramoneadores, ramoneadores frugívoros, dieta mixta alimento por alimento, dieta mixta estacional/regional y pacedores. Finalmente, se obtuvo una gráfica bivariante con el número promedio de fosas y el número promedio de rayones, con los datos de las especies de ungulados actuales de la base de datos de Solounias y Semprebon (2002) y de los mamuts considerados en el estudio.
Los análisis estadísticos se realizaron en los programas Past Versión 3.04 (Hammer et al., 2001) y STATISTICA 10.
4. Paleontología sistemática
Orden Proboscidea Illiger, 1811
Familia Elephantidae Gray, 1821
Género MammuthusBrookes, 1828
Mammuthus columbiFalconer, 1857

Figura 2 Mandíbula de Mammuthus columbi del Pleistoceno tardío de Hidalgo. UAHMP-4220, vistas lateral (A) y oclusal (B).

Figura 3 Incisivos de Mammuthus columbi del Pleistoceno tardío de Puebla y Oaxaca. UAHMP-924 (A), fragmento del extremo proximal de una defensa; UAHMP-4189 (B) y UMPE-526 (C) ápices de defensas.

Figura 4 Molariformes superiores de Mammuthus columbi del Pleistoceno tardío de Oaxaca. UMPE-022, vistas oclusal (A) y lateral (A1) de un M2/M3I; UMPE-029-02, vistas oclusal (B) y lateral (B1) de un M3D; UMPE-467, vistas oclusal (C) y lateral (C1) de un M3I.

Figura 5 Molariformes inferiores de Mammuthus columbi del Pleistoceno tardío de Hidalgo y Puebla. UMPE-618, vistas oclusal (A) y lateral (A1) de un dp4I; UMPE-506, vistas oclusal (B) y lateral (B1) de un dp4/m1D; UAHMP-4002, vistas oclusal (C) y lateral (C1) de un m2/m3I; UAHMP-4190, vistas oclusal (D) y lateral (D1) de un m3.

Figura 6 Vértebras de Mammuthus columbi del Pleistoceno tardío de Hidalgo, Puebla y Oaxaca. UAHMP-218, vistas craneal (A) y caudal (A1) de un atlas; UMPE-027-0, vistas craneal (B) y caudal (B1) del cuerpo de una vértebra cervical; UAHMP-4220, vistas craneal (C) y caudal (C1) de una vértebra torácica.

Figura 7 Elementos postcraneales de Mammuthus columbi del Pleistoceno tardío de Hidalgo, Puebla y Oaxaca. UAHMP-1107, vistas lateral (A) y ventral (A1) del fragmento de una escápula; UMPE-016, vistas anterior (B) y posterior (B1) del fragmento de un radio; UMPE519, vista lateral (C) de un fragmento de pelvis; UMPE-003, vistas proximal (D) y posterior (D1) del extremo proximal de una tibia; UAHMP-4172, vista ventral (E) de un astrágalo; UAHMP-571, vista proximal (F) de un calcáneo.
Tabla 1 Medidas (en mm) tomadas en mandíbulas de individuos referidos a Mammuthus columbi del Pleistoceno tardío de Hidalgo y Oaxaca. Abreviaturas: ACM, altura del cuerpo mandibular; LM, longitud máxima; ANCM, anchura del cuerpo mandibular; FM, número de forámenes mentales; FS, número de forámenes sublinguales; AAC, altura de la apófisis coronoides; AMH, anchura máxima entre las hemimandíbulas; LPM-AL, longitud entre el borde del proceso mental y los bordes anteriores del alvéolo; LCI, longitud de la cresta interalveolar; AS, altura de la sínfisis; LS, longitud de la sínfisis; ANS, anchura de la sínfisis; DH-AAL, distancia entre las hemimandíbulas al nivel del borde anterior del alvéolo; DH-PAL, distancia entre las hemimandíbulas al nivel del borde posterior del alvéolo.
| UMPE-617 | UMPE-614.1 | UMPE-614.2 | UAHMP-4220 | |
| ACM | 190 | 190 | 220 | |
| LM | 300 | 313.9 | 610 | |
| ANCM | 150 | 174.3 | 131 | 152.4 |
| FM | 4 c/lado (8 totales) | |||
| FS | 2 | |||
| AAC | 165.1 | |||
| AMH | 558.8 | |||
| LPM-AL | 152.4 | |||
| LCI | 254 | |||
| AS | 127 | |||
| LS | 120.65 | |||
| ANS | 114.3 | |||
| DH-AAL | 101.6 | |||
| DH-PAL | 203.2 |
Tabla 2 Medidas (en mm) de la longitud total y del diámetro (medido cada 10 cm del extremo distal al extremo proximal del elemento) de los incisivos de Mammuthus columbi del Pleistoceno tardío de Puebla y Oaxaca.
| UMPE-0076 | UMPE-526 | UMPE-619 | UAHMP-924 | UAHMP-4189 | |
| Longitud total del elemento | 2000 | 220 | 900 | 1050 | 300 |
| Extremo distal | 182 | 84.34 | 207 | 102.95 | 81.88 |
| 197 | 70.06 | 203.52 | 105.83 | 81.58 | |
| 198 | 28.85 | 200 | 104.18 | 68 | |
| 202 | 102.1 | 25.1 | |||
| 206 | 99.77 | ||||
| 208 | 94.4 | ||||
| 213 | 88.85 | ||||
| 210 | 81.5 | ||||
| 192 | 75.06 | ||||
| 180 | 72.11 | ||||
| 172 | 39.77 | ||||
| 170 | |||||
| 177 | |||||
| 166 | |||||
| Extremo proximal | 171 |
Tabla 3 Parámetros cuantitativos evaluados en molares de Mammuthus columbi del Pleistoceno tardío de los estados de Hidalgo, Puebla y Oaxaca. Abreviaturas: D, derecho; I, izquierdo; AEA, años de elefante africano; Fl, frecuencia lamelar (número de lamelas que ocurren en una longitud de 100 mm); En, grosor del esmalte; Pl, número de lamelas preservadas; W, amplitud de la lamela más ancha excluyendo el cemento marginal; Lg, longitud del ejemplar medida en línea recta lo más cerca posible a los ángulos rectos del eje general de la lamela excluyendo el cemento marginal; A, número de lamelas desgastadas; Al, altura de la lamela más profunda excluyendo la raíz; C, grosor del cemento en la superficie oclusal; R, cociente resultado de dividir la longitud (Lg) entre el número de lamelas preservadas (Pl); IH, índice de hipsodoncia (Ht /W); Ld, longitud máxima en un plano diagonal; Lo, longitud máxima en un plano oclusal; Ht, altura máxima del diente paralela al plano de orientación. *Elementos que probablemente pertenecen a individuos machos, con base en su anchura, frecuencia lamelar y/o grosor del esmalte (McDaniel y Jefferson, 2006a, tabla 4, p. 174). Las medidas están en mm.
| Estado | Localidad | Ejemplar | Posición | Edad (AEA) |
Fl | En | Pl | W | Lg | A | Al | C | R | IH | Ld | Lo | Ht |
| Oaxaca | OAX-4 | UMPE-029-1 | dp4 | 3 | 1.54 | 7 | 42.54 | 84.43 | 7 | 75.5 | 3 | 12.06 | 2.26 | 86.4 | 85.35 | 96.15 | |
| OAX-7 | UMPE-618 | dp4 I | 5 | 7 | 1.89 | 7 | 42.37 | 87.54 | 7 | 86.36 | 3.83 | 12.51 | 2.31 | 115.06 | 107.65 | 97.9 | |
| OAX-7 | UMPE-506 | dp4/m1 D | 3 - 6 | 6 | 2.32 | 7 | 74.67 | 96.73 | 6 | 91.55 | 5 | 13.82 | 1.65 | 135.96 | 127.01 | 123.34 | |
| OAX-7 | UMPE-525 | m1/m2 D | 16 | 2.64 | 3 | 72.79 | 57.43 | 3 | 95.55 | 5.61 | 19.14 | 1.44 | 61.93 | 56.38 | 105.37 | ||
| OAX-7 | *UMPE 614 | m2/m3 D | 37 - 38 | 5 | 2.75 | 84.59 | 154.72 | 7.5 | 6 | ||||||||
| OAX-7 | *UMPE 614 | m2/m3 I | 37 - 38 | 6 | 2.83 | 82.21 | 113.36 | 6.5 | 6 | ||||||||
| OAX-2 | *UMPE-022 | M2/M3 I | 15/28 - 30 | 6 | 2.49 | 9 | 68.6 | 130.19 | 8 | 122.09 | 5.93 | 16.27 | 2.91 | 260 | 160 | 200 | |
| OAX-7 | UMPE-467 | M3 I | 30 | 7 | 2.89 | 15 | 72.35 | 132.23 | 8 | 190 | 3.57 | 16.53 | 2.76 | 285 | 165 | 200 | |
| OAX-4 | UMPE-029-02 | M3 D | 30 | 6 | 2.83 | 13 | 87.17 | 108.76 | 7 | 257 | 4.39 | 15.54 | 2.63 | 280 | 170 | 230 | |
| OAX-7 | UMPE-524 | M3 | 30 - 32 | 6 | 2.7 | 6 | 57.15 | 126 | 0 | 194.49 | 21 | 207.01 | 98.96 | ||||
| Puebla | PUE-3 | UAHMP-228 | dp4 D | 3 | 5 | 3.52 | 5 | 58.31 | 119.34 | 4 | 48.68 | 10 | 23.87 | 2.04 | 170 | 150 | 119.52 |
| PUE-2 | UAHMP-229 | dp4/m1 D | 3 - 6 | x | 2.68 | 5 | 58.6 | 70.5 | 5 | 122.48 | 4 | 14.1 | 2.38 | 119.54 | 97.99 | 139.48 | |
| PUE-7 | *UAHMP-4002 | m2/m3 D | 16/35 | 6 | 2.84 | 7 | 71.38 | 131.34 | 7 | 150.43 | 6.61 | 18.76 | 2.36 | 190 | 180 | 170 | |
| Axamilpa | UAHMP-4191 | M3 | 40 | 6 | 3.26 | 10 | 77.12 | 153.7 | 6 | 210 | 15.37 | 2.72 | 300 | 210 | 210 | ||
| PUE-3 | UAHMP-227 | Indet. | 3 | 2.36 | 4 | 86.8 | 192 | 153.48 | 7 | 48 | |||||||
| Hidalgo | HGO-46 | TCH-056 | dp4 I | 5 | 8 | 1.94 | 8 | 46.56 | 102.44 | 8 | 48.93 | 4 | 12.81 | 1.46 | 109.42 | 109.65 | 68.09 |
| HGO-49 | UAHMP-3972 | dp4/m1 D | 3 - 6 | 6 | 2.48 | 8 | 40.2 | 115.25 | 8 | 120.84 | 4 | 14.41 | 3.58 | 150 | 142 | 144.11 | |
| Cd. Sahagún | UAHMP-4190 | m3 | 35 | 6 | 3.07 | 5 | 92.59 | 96.73 | 5 | 157.67 | 5 | 19.35 | 2.13 | 111 | 101 | 198 | |
| HGO-50 | *UAHMP-4220 | m3 D | 46 - 48 | 5 | 2.96 | 8 | 89.37 | 241.3 | 8 | 7.44 | 30.16 | 255 | |||||
| HGO-50 | *UAHMP-4220 | m3 I | 46 - 48 | 5 | 2.65 | 6 | 77.16 | 215.9 | 6 | 7.51 | 35.98 | 232 |
Tabla 4 Medidas (en mm) de las vértebras de Mammuthus columbi del Pleistoceno tardío de los estados de Hidalgo, Puebla y Oaxaca. Abreviaturas: DAPC, diámetro anteroposterior del cuerpo; AMC, anchura máxima del cuerpo; AlMC, altura máxima del cuerpo; LT, longitud total de la vértebra; LAE, longitud de la apófisis espinosa; AMCN, anchura máxima del canal neural; AlMCN, altura máxima del canal neural.
| Posición | Ejemplar | DAPC | AMC | AlMC | LT | LAE | AMCN | AlMCN |
| Cervicales | UMPE-027-0 | 33.29 | 122.48 | 109.79 | ||||
| UAHMP-963 | 81.99 | 180 | 150 | |||||
| Torácicas | UMPE-025 | 80.3 | 153.25 | 132.04 | ||||
| UMPE-0075 | 78 | 141.24 | 158.04 | |||||
| UMPE-503-1 | 77.01 | 120.91 | 138.23 | |||||
| UMPE-527 | 54.37 | 130.31 | 107.86 | |||||
| UAHMP-948 | 450 | |||||||
| UAHMP-4220 (a) | 64.52 | 126.24 | 160 | 710 | 450 | 75.75 | ||
| UAHMP-4220 (b) | 71.87 | 121.91 | 133.1 | 670 | 484 | 76.51 | 56.96 | |
| UAHMP-4220 (c) | 71.83 | 143.02 | 139.9 | 420 | 220 | 64.46 | 66.24 | |
| UAHMP-4220 (d) | 56.63 | 154.75 | 156.4 | 600 | 357 | 75.54 | 88.72 | |
| UAHMP-4220 (e) | 76.53 | 127.96 | 127.92 | 550 | 350 | 57.58 | 66.97 | |
| UAHMP-4220 (f) | 139.57 | 127.67 | 530 | 348 | 56.68 | 70.96 | ||
| UAHMP-4220 (g) | 74.85 | 129.71 | 127.83 | 557 | 354 | 60.11 | 65.46 | |
| UAHMP-4220 (h) | 73.66 | 134.66 | 124.95 | 550 | 346 | 64.09 | 63.5 | |
| UAHMP-4220 (i) | 402 | 210 | 62.24 | 59.71 | ||||
| UAHMP-4220 (j) | 76.28 | 134.71 | 125.63 | 360 | 176 | 60.75 | 67.81 | |
| UAHMP-4220 (k) | 190 | 65.76 | 62.79 | |||||
| UAHMP-4220 (l) | 180 | 71.44 | 63.96 | |||||
| UAHMP-4220 (m) | 196 | |||||||
| Lumbares | UMPE-028-1 | 72.85 | ||||||
| UMPE-028-3 | 59.5 | 140.62 | 108.74 | 31.89 | ||||
| UAHMP-187 | 67.48 | 136.52 | 105.29 | 334 | 143.78 | 83.82 | 48.13 | |
| UAHMP-236 | 139.35 | |||||||
| UAHMP-237 | 78.1 | 154 | 134.3 | |||||
| UAHMP-238 | 64.97 | 142.54 | 140.51 | |||||
| UAHMP-239 | 57.12 | 126.83 | 129.77 | 370 | 154 | 69.24 | 55.31 | |
| UAHMP-240 | 198 |
4.1. ALCANCE GEOCRONOLÓGICO Y DISTRIBUCIÓN GEOGRÁFICA
La especie Mammuthus columbi estuvo ampliamente distribuida en Norteamérica desde Alaska hasta Nicaragua, con un alcance biocronológico del Irvingtoniano medio al Rancholabreano (Kurtén y Anderson, 1980; Agenbroad, 2005). En México este proboscídeo procede de numerosos depósitos del Pleistoceno tardío ubicados a lo largo del territorio nacional, excepto en la Península de Yucatán y Tabasco (Kurtén y Anderson, 1980; Lange, 2002; Arroyo-Cabrales et al., 2003, 2007; Agenbroad, 2005; Lister y Bahn, 2007).
4.2. MATERIAL REFERIDO
Hidalgo. La Pinta de Vaquerías (HGO-10): UAHMP-187 fragmento de vértebra lumbar, UAHMP-153 fragmento del cuerpo de una vértebra; Barranca El Berrendo (HGO-28): UAHMP4219 fragmento de un premolar deciduo; Barranca Jagüey Viejo (HGO-42): UAHMP-908 fragmentos dentales; Las Cajas (HGO-46): TCH-056 dp4 izquierdo, UAHMP-948 vértebra torácica; El Barrio (HGO-47): UAHMP-1107 fragmento escapular, UAHMP-963 vértebra cervical; San Gabriel Azteca (HGO-49): UAHMP-3972 dp4/ m1 derecho, UAHMP-3983 fragmento de lamela; Singuilucan (HGO-50): UAHMP-4220 fragmento de cráneo, mandíbula rota con dos m3 y trece vértebras torácicas asociadas; Ciudad Sahagún, Municipio de Tepeapulco: UAHMP-4190 fragmento de m3 izquierdo.
Puebla. Barranca Policarpio (PUE-2): UAHMP226 fragmento del extremo distal de un incisivo, UAHMP-229 dp4/m1 derecho, UAHMP-218 atlas, UAHMP-236, UAHMP-237, UAHMP-238, UAHMP-239 y UAHMP-240 vértebras lumbares, UAHMP-571 fragmento de astrágalo; Dos Cruces (PUE-3): UAHMP-228 dp4 derecho, UAHMP227 fragmentos dentales; Santa Cata (PUE-4): UAHMP-533 fragmentos dentales; Cuahutinchan (PUE-6): UAHMP-573 fragmento de vértebra lumbar, UAHMP-574 fragmento de vértebra; Barranca Xocoa (PUE-7): UAHMP-4189 fragmento del extremo distal de un incisivo, UAHMP924 fragmento de incisivo, UAHMP-4002 m2/m3 derecho, UAHMP-4172 fragmento de calcáneo; Axamilpa: UAHMP-4191 fragmento de M3.
Oaxaca. El Pedernal (OAX-2): UMPE-022 M2/M3 izquierdo, UMPE-005 fragmento de escápula, UMPE-616 fragmento de diáfisis de fémur, UMPE-003 extremo distal de tibia; Río Salado (OAX-4): UMPE-029-2 M3 derecho, UMPE-029-1 dp4 lado indeterminado, UMPE0075 vértebra torácica, UMPE-016 fragmento de radio; Llano de Hueso (OAX-5): UMPE-617 fragmento de mandíbula, UMPE-0076 fragmento de incisivo; Río Tejupam (OAX-7): UMPE-614 mandíbula rota en dos fragmentos, con dos m3, UMPE-526 fragmento del extremo distal de un incisivo, UMPE-619 fragmento de maxila con parte del incisivo, UMPE-467 M3 lado indeterminado, UMPE-524 fragmento de M3, UMPE 506 dp4/m1 derecho, UMPE-618 dp4 izquierdo, UMPE- 027-0 vértebra cervical, UMPE-025, UMPE-503-1 y UMPE-527 vértebras torácicas, UMPE-028-1 y UMPE-028-3 vértebras lumbares, UMPE-027-1, UMPE-027-2, UMPE-027-3, UMPE-028-2 y UMPE-503-2 fragmentos de vértebra, UMPE-630 fragmento de escápula, UMPE-519 fragmento de pelvis.
4.3. ADESCRIPCIÓN DE LOS EJEMPLARES
4.3.1. CRÁNEO
El fragmento de cráneo del individuo catalogado como UAHMP-4220 está colapsado y solo se distingue una parte del arco cigomático.
4.3.2. MANDÍBULA
La descripción de este elemento se basa en la mandíbula casi completa del individuo UAHMP-4220 (Tabla 1, Figura 2). El cuerpo de la mandíbula y la rama mandibular son cortas y robustas; la apófisis coronoides es corta, está aplanada latero-medialmente y ubicada anteriormente con respecto al alvéolo del último molar; el ángulo mandibular es redondeado; cada hemimandíbula presenta cuatro forámenes mentonianos de diversas profundidades y diámetros, así como una foseta sublingual pequeña y profunda; la protuberancia mentoniana está bien desarrollada y se proyecta hacia abajo. El ejemplar UMPE-617 corresponde a la parte más posterior de una mandíbula derecha, al igual que en UAHMP-4220, la rama mandibular es corta, robusta y además cóncava en la cara lateral. Por su parte, el ejemplar UMPE-614 incluye la parte más posterior de los cuerpos mandibulares de ambos lados con los últimos molares (m3).
4.3.3. INCISIVOS
Incluyen un conjunto de cinco fragmentos de defensa que se caracterizan por ser gruesos hacia el extremo proximal y disminuyen en circunferencia hacia el extremo distal del elemento; el ejemplar UMPE-0076 es el de mayor longitud y corresponde a la porción media de una defensa (Tabla 2). Los ejemplares UMPE-526, UAHMP924 y UAHMP-4189 corresponden a fragmentos del extremo distal de una defensa, los cuales tienen forma cónica y están ligeramente curvos; la punta es roma en UMPE-52, mientras que es aguzada en UAHMP-924 y UAHMP-4189 (Figura 3). En algunos de los elementos es posible observar al menos cinco anillos de crecimiento, tal es el caso de UMPE-526 y UAHMP-226.
4.3.4. MOLARES
Los molares se describen de acuerdo a su posición dental. Las medidas tomadas en cada uno de los elementos dentales se indican en la Tabla 3.
Molariformes superiores (Figura 4). El ejemplar UMPE-022 corresponde a la porción anterior de un segundo molar o un tercer molar en cuya superficie oclusal se observan nueve lamelas de las cuales ocho están desgastadas, la frecuencia lamelar es de seis, tiene un grosor del esmalte de 2.5 mm y un índice de hipsodoncia cercano a tres. El elemento se encuentra en un estado de desgaste que sugiere una edad de 15 años si se tratase de un segundo molar y de 28 - 30 años si fuese un tercer molar. Con base en esto, la edad del individuo estaría alrededor de los 25 años.
Los ejemplares UMPE-029-02, UMPE-524, UMPE-467 y UAHMP-4191 corresponden a terceros molares, los cuales tienen una frecuencia lamelar que varía de seis a siete, un grosor del esmalte cercano a los 3 mm y un índice de hipsodoncia promedio de 2.7. Los ejemplares UMPE-029-2 y UMPE-467 están ligeramente fragmentados, muestran de 13 a 15 lamelas respectivamente de las cuales alrededor del 50% están desgastadas; ambos elementos están en un estado de desgaste que sugiere una edad de aproximadamente 30 años. El ejemplar UAHMP-4191 corresponde a la porción posterior del diente que muestra diez lamelas de las cuales seis están desgastadas, se encuentra en un estado de desgaste que sugiere una edad de alrededor de 40 años si se toma en cuenta que las últimas lamelas están ligeramente desgastadas. El elemento UMPE-524 corresponde a la sección media del diente en el que se observan seis lamelas sin desgastar y cuyo estado de desgaste es indicativo de una edad entre 30 y 32 años.
Molariformes inferiores (Figura 5). Los ejemplares UAHMP-228, TCH-56, UMPE-618 y UMPE029-01 corresponden a cuartos premolares deciduos. Los tres primeros están prácticamente completos, mientras que el otro consiste en la porción anterior del diente. La frecuencia lamelar de los ejemplares varía de cinco a siete, tienen un grosor del esmalte promedio de alrededor de 2 mm y un índice de hipsodoncia promedio de 2.0; en estos elementos todas las lamelas expuestas están desgastadas, excepto en UAHMP-228, en el que sólo el 80% están desgastadas. Con base en el estado de desgaste, los ejemplares TCH-56, UMPE-618 representarían una edad aproximada de cinco años, mientras que los ejemplares UAHMP-228 y UMPE-029-01 representarían una edad de alrededor de tres años.
Los ejemplares UMPE-506, UAHMP-3972 y UAHMP-229 son partes de un cuarto premolar deciduo o un primer molar, tienen un grosor del esmalte de alrededor 2.5 mm y un índice de hipsodoncia promedio de 2.5. Los ejemplares UMPE-506 y UAHMP-3972 corresponden a la sección anterior, tienen ocho lamelas desgastadas y una frecuencia lamelar de seis, mientras que el ejemplar UAHMP-229 corresponde a la sección media, presenta cinco lamelas desgastadas y cuya frecuencia lamelar se desconoce debido a que el fragmento tiene una longitud inferior a los 10 cm.
Con base en el estado de desgaste, los ejemplares representarían una edad entre tres y seis años. El ejemplar UMPE-525 es un fragmento de la sección central de un primer molar o un segundo molar, el cual tiene tres lamelas que están desgastadas, grosor del esmalte promedio cercano a 2.5 mm, así como una longitud inferior a los 10 cm y, por lo tanto, no es posible estimar la frecuencia lamelar; asimismo, tiene un índice de hipsodoncia de 1.44. El estado de desgaste del elemento sugiere una edad de ocho años si fuese un m1 y de unos 16 años si se tratase de un m2.
El ejemplar UAHMP-4002 corresponde a la sección central de un segundo molar o un tercer molar, tiene siete lamelas desgastadas, frecuencia lamelar de seis, grosor del esmalte promedio cercano a los 3 mm y un índice de hipsodoncia de 2.36; el desgaste del elemento correspondería a una edad mínima de 16 años si fuese un m2 y de 35 años si se tratase de un m3.
El ejemplar UAHMP-4190 corresponde a la sección central de un tercer molar, tiene cinco lamelas todas desgastadas, el desgaste es mayor hacia el borde lingual y la superficie oclusal está inclinada hacia ese borde, de tal forma que la altura de las lamelas es mayor hacia el borde labial; en este ejemplar la frecuencia lamelar es de seis, el grosor del esmalte promedio es de 3 mm, mientras que el índice de hipsodoncia es de 2.13; el estado de desgaste del elemento se asociaría a una edad de alrededor 35 años.
Los últimos molares engastados en la mandíbula catalogada como UMPE-614 están rotos de manera semejante y sólo se conserva la parte posterior de cada diente, en ambos molares se pueden contar diez lamelas de las cuales siete están desgastadas, la frecuencia lamelar es de 5.5 y el grosor promedio del esmalte es de 2.79 mm. El estado de desgaste de estos elementos se asociaría a una edad de 37 - 38 años.
En cada hemimandíbula del ejemplar UAHMP-4220 se conserva un último molar, los cuales se encuentran oblicuos al plano sagital del cuerpo de la mandíbula. El molar derecho tiene ocho lamelas preservadas y el molar izquierdo tiene seis, en ambos casos, cinco de las lamelas están desgatadas. La frecuencia lamelar de los dos molares es de cinco, mientras que el grosor promedio del esmalte es de 2.8 mm. El estado de desgaste general de ambos molares corresponde a una edad aproximada de 46 a 48 años.
En la muestra estudiada también se reconocen varios fragmentos de lamela y fragmentos dentales que debido a su estado de preservación son poco informativos.
4.3.5. POSTCRANEALES
Vértebras (Figura 6). Las vértebras son los elementos más abundantes y diversos de los elementos del postcráneo que integran la muestra estudiada (Tabla 4). El ejemplar UAHMP-218 es un atlas que se distingue por ser robusto, tiene forma subovalada, su diámetro transversal es de 419.1 mm y el diámetro medial es de 241.3 mm. Las apófisis transversas están rotas de los extremos, al igual que el tubérculo ventral; sin embargo, es posible observar algunos forámenes en la superficie de las apófisis. Por otra parte, el margen dorsal es ligeramente convexo tanto en vista anterior como posterior. El foramen vertebral tiene forma de “8”, en el que el círculo superior es de mayor anchura que el inferior. Las facetas articulares para los cóndilos occipitales son ovoidales, elongadas dorsoventralmente, cóncavas y su longitud es mayor a la del foramen vertebral, mientras que las facetas articulares para el axis son ovoidales, ligeramente cóncavas y su longitud es menor que la del foramen vertebral. Los surcos para la arteria vertebral son circulares, en tanto que los forámenes transversos son semicirculares.
Los ejemplares UMPE-027 y UAHMP-963 corresponden a vértebras cervicales que se distinguen por tener un cuerpo vertebral ligeramente más ancho que alto (1:0.86), el cual es de forma circular tanto en vista anterior como posterior y en vista lateral está aplanado antero-posteriormente. El cuerpo vertebral es cerca de 12 % y 20 % más aplanado en comparación con los cuerpos vertebrales de las vértebras torácicas y lumbares.
En las vértebras torácicas el cuerpo vertebral es tan ancho como alto (1:0.99), su cara anterior es circular, excepto en UMPE-503-1 y UMPE-527 en la que tiene forma subtriangular y en UMPE075 es cordiforme. La cara posterior del cuerpo vertebral es generalmente cordiforme, a excepción de UAHMP-4220(c) y UAHMP-4220(d) en donde es circular, mientras que en UMPE-527 y UAHMP-4220(b) es subtriangular. Estas vértebras tienen un cuerpo vertebral más grueso en comparación al de las vértebras cervicales (1:0.88) y las lumbares (1: 0.91). La apófisis espinosa es larga (307.93 mm en promedio), condición más evidente en las vértebras de la región anterior y va disminuyendo su longitud en las vértebras de la región posterior. Algo parecido ocurre con la robustez de la apófisis espinosa, de tal suerte que las espinas son más robustas en las vértebras de la región anterior. El surco que recorre la cara posterior de la apófisis espinosa es profundo, aunque en UAHMP-4220(a) es poco profundo. El canal neural es generalmente de forma circular; sin embargo, en UAHMP-4220(a), UAHMP-4220(c) y UAHMP-4220 (d) es triangular, mientras que en UAHMP-4220 (b) es ovoidal. La variación en la forma del canal neural podría relacionarse a la posición de la vértebra en la caja torácica.
Las facetas articulares craneales de las vértebras torácicas se ubican en la cara dorsal de la vértebra en la base de la apófisis espinosa. La forma de estas facetas varía de circular a ovoidal. Por su parte, las facetas articulares caudales también se encuentran en la base de la apófisis espinosa pero en la cara ventral de la misma, las cuales son ovoidales, excepto en UAHMP-4220(k) que son circulares. En las caras laterales del cuerpo de las vértebras torácicas se hallan las facetas costales, tanto las anteriores como las posteriores son de forma ovoidal. Conforme la vértebra está más cercana a la región pélvica, el número de facetas costales se reduce de cuatro a dos.
Las vértebras torácicas tienen un total de tres apófisis: dos transversas y una espinosa. Sin embargo, algunas como UAHMP-4220(a), UAHMP-4220(f) y UAHMP-4220(k) están incompletas y sólo presentan dos apófisis. En el ejemplar UAHMP-948 solamente está preservada la apófisis espinosa y en los ejemplares UMPE-025, UMPE0075, UMPE-503-1 y UMPE-527 únicamente el cuerpo vertebral.
Las vértebras lumbares tienen un cuerpo vertebral ligeramente más ancho que alto (1:0.91) que es cordiforme en vista anterior, salvo en UMPE-28-3 que tiene forma de media luna y en UAHMP-237 donde es subredondeado. En dicho cuerpo estas vértebras son ligeramente menos gruesas en comparación a las torácicas (1:0.99); sin embargo, son más gruesas que en las cervicales (1:0.88). La apófisis espinosa es corta (142 mm en promedio), en cuya base se observa una escotadura pronunciada en forma de “V” invertida. Las facetas articulares craneales como las caudales son de forma ovoidal y se encuentran en la base de la apófisis espinosa, las primeras en la cara dorsal y las segundas en la ventral. Ninguno de los ejemplares que corresponden a este tipo de vértebra está completo, por lo que tienen entre una y tres apófisis.
La muestra estudiada también incluye varios fragmentos de vértebra cuyo estado de preservación limita determinar la posición que tienen en la columna vertebral.
Escápula. El ejemplar UMPE-005 es un fragmento proximal de escapula derecha que conserva una parte de la cavidad glenoidea y de la base del acromio. En la superficie medial y la lateral del elemento se pueden observar algunos forámenes. La longitud de la cavidad glenoidea es de alrededor de 120 mm.
El ejemplar UMPE-630 es un fragmento de escápula derecha en el que se conserva la parte más proximal del elemento, la cavidad glenoidea completa y parte de las fosas supra e infraespinosa, así como la base del acromio. La cavidad glenoidea tiene forma rectangular, tiene una longitud de 203 mm y una anchura de 110 mm. El cuello de la escápula está completamente preservado y tiene una longitud de 225 mm. El acromio se sitúa aproximadamente a la mitad de la espina escapular. El ejemplar UAHMP-1107 es la parte proximal de una escápula izquierda, se conserva solamente una parte de la cavidad glenoidea y una pequeña porción de la base de la fosa infraespinosa. Aparentemente la cavidad glenoidea es alargada. El cuello de la escápula está constreñido, aún cuando no está completo. Es posible observar algunos forámenes en la superficie lateral del elemento (Figura 7A a 7A1).
Radio. El ejemplar UMPE-016 incluye la parte media de la diáfisis de un radio izquierdo, la cuál está rota en ambos extremos (Figura 7B a 7B1). La longitud máxima es de 620 mm y el diámetro mínimo es de 200 mm.
Pelvis. El elemento UMPE-519 es un fragmento de pelvis del lado derecho, el cual es masivo y robusto. En este ejemplar se conserva el acetábulo, una parte del eje del hueso iliaco y un fragmento del isquion (Figura 7C). La faceta articular es subovoidal (largo= 150 mm, ancho= 100 mm), y hacia el lado ventral de ésta se encuentra la fosa acetabular que tiene forma circular (40 x 40 mm); además la cresta iliaca presenta varios forámenes nutricios. Por el grado de preservación del ejemplar, únicamente es posible tomar la anchura mínima del eje del ilion que es de 210 mm aproximadamente.
Tibia. El ejemplar UMPE-003 corresponde al extremo proximal de una tibia izquierda, la longitud total del ejemplar es de alrededor de 300 mm y la anchura máxima de la epífisis proximal es de aproximadamente 200 mm (Figura 7D a 7D1). La cara anterior del elemento está rota, mientras que la cara posterior es cóncava y se aprecia la eminencia intercondilar. El cóndilo medial tiene forma circular mientras que el cóndilo lateral es de forma ovoidal.
Fémur. El ejemplar UMPE-616 es el fragmento de la diáfisis de un fémur, el cual tiene una longitud de 360 mm y anchura mínima de 390 mm. En la cara anterior se aprecia un fragmento de lo que aparentemente es la fosa supracondiloidea.
Astrágalo. El ejemplar UAHMP-4172 consiste en aproximadamente el 60% de un astrágalo (Figura 7E), el cual es aplanado dorsoventralmente y en vista ventral es posible observar algunas porciones de las facetas articulares para el calcáneo.
Calcáneo. El elemento UAHMP-571 es un fragmento de calcáneo cuya parte mejor preservada es el tuber calcis, el cual tiene la forma alargada típica del calcáneo en los mamuts y en los mamíferos en general (Figura 7F). En el resto del elemento es posible distinguir algunas partes de las facetas articulares.
4.4. EDAD Y SEXO DE LOS EJEMPLARES
Con base en la localidad, la posición dental y la edad estimada, el conjunto de molariformes considerados representaría de 15 a 16 individuos repartidos así, entre siete u ocho en el estado de Oaxaca, cuatro en el estado de Puebla y cuatro en el estado de Hidalgo. La categoría mejor representada es la de adultos maduros con una edad ≥ 30 años, seguida de los adolescentes con un intervalo de edad de 3 a 17 años; entre éstos, aquellos considerados en la categoría de 3- 6 años son más comunes. Con base en los criterios de McDaniel y Jefferson (2006a), entre los adultos maduros, los individuos representados por los ejemplares UMPE-614, UMPE-022, UAHMP- 4002 y UAHMP-4220, podrían ser machos, dado que presentan una frecuencia lamelar entre 5 y 6, así como un grosor del esmalte ≥ 2.5 mm (Tabla 3). Esto indica la presencia de al menos un macho adulto y sexualmente activo en cada uno de los conjuntos estudiados, asumiendo que los individuos de Mammuthus columbi hayan alcanzado tal estado sexual equiparable al que se observa en los elefantes asiáticos actuales (Elephas maximus) de alrededor de los 30 años (Macdonald, 2006).
5. Análisis de microdesgaste
En términos generales, las muestras de Hidalgo, Puebla y Oaxaca tienen un mayor número promedio de rayones que de fosas, lo cual se asociaría a un comportamiento alimentario con énfasis en recursos abrasivos; asimismo, se observa una frecuencia importante de rayones gruesos (> 60%) (Figura 8). Las muestras de Hidalgo y Puebla se distinguen por presentar una alta frecuencia de gubias; asimismo, en la muestra de Puebla los rayones cruzados son comunes (ca. 70%) (Tabla 5).

Figura 8 Microfotografías de bandas de esmalte de molares de Mammuthus columbi del Pleistoceno tardío de los estados de Hidalgo, Puebla y Oaxaca, regiones centro y sur de México. Los cuadros en líneas blancas indican el área de conteo estándar (0.4 X 0.4 mm) para la evaluación de las variables convencionales de microdesgaste (rayones y fosas). A, UAHMP-4220 (m3I); B, UAHMP-4002 (m2/m3D); C, UMPE-029-02 (M3D). Barras de escala de 0.4 mm.
Tabla 5 Variables de microdesgaste en muestras de Mammuthus columbi del Pleistoceno tardío de Hidalgo, Puebla y Oaxaca, centro y sur de México. Abreviaturas: PR, promedio de rayones; PF, promedio de fosas; %RC, porcentaje de rayones cruzados; %FG, porcentaje de fosas grandes; %G, porcentaje de gubias; %rf, porcentaje de rayones finos; %rg, porcentaje de rayones gruesos; %rm, porcentaje de rayones mixtos.
| Muestra | PR | PF | %RC | %FG | %G | %rf | %rg | %rm |
| Hidalgo | 22.43 | 15 | 12.5 | 25 | 87.5 | 25 | 75 | 0 |
| Puebla | 23.37 | 12.12 | 75 | 0 | 100 | 25 | 75 | 0 |
| Oaxaca | 21.85 | 13.35 | 28.5 | 21.4 | 50 | 35.7 | 64.2 | 0 |
En la gráfica de los dos primeros componentes que resultan del ACP de los datos de microdesgaste, la muestra de Oaxaca se ubica en el espacio multivariado de los pacedores típicos actuales cercana al bisonte Americano (Bison bison), la muestra de Puebla se ubica en el espacio multivariado de ungulados actuales de hábitos mixtos cercano al buey almizclero (Ovibos moschatus), mientras que la muestra de Hidalgo se coloca entre los ungulados pacedores y los de hábitos mixtos cercana al antílope acuático (Kobus ellipsiprymnus) (Figura 9).

Figura 9 Gráfica de los dos primeros componentes que resultan del Análisis de Componentes Principales (ACP) de los datos de microdesgaste en molares de Mammuthus columbi del Pleistoceno tardío de Hidalgo (1), Puebla (2) y Oaxaca (3), así como los de la base de datos de ungulados actuales de Solounias y Semprebon (2002). Abreviaturas: Ramoneadores folívoros BE, Boocercus eurycerus; TI, Tragelaphus imberbis; GC, Giraffa camelopardalis; CB, Camelus bactrianus; AA, Alces alces; LW, Litocranius walleri; TT, Tragelaphus strepsiceros; CL, Camelus dromedarius; DB, Diceros bicornis; AM, Antilocapra americana. Ramoneadores frugívoros: NI, Cephalophus niger; TERR, Tapirus terrestris; DR, Cephalophus dorsalis; SL, Cephalophus silvicultor; OJ, Okapia johnstoni; TRA, Tragulus sp.; MO, Moschus moschiferus; TBA, Tapirus bairdii; NA, Cephalophus natalensis. Hábitos mixtos estacionales: To, Taurotragus oryx; Ca, Capricornis sumatraensis; Bt, Budorcas taxicolor; Gg, Gazella granti; Ts, Tragelaphus scriptus; Vi, Lama vicugna; Oc, Ovis canadensis; Lg, Lama glama; Gt, Gazella thomsoni; Tr, Boselaphus tragocamelus; Ax, Axis axis; Mm, Muntiacus muntjak. Hábitos mixtos comidapor-comida: Cu, Cervus unicolor; Om, Ovibos moschatus; Cc, Cervus canadensis; Ci, Capra ibex. Pacedores: ct, Connochaetes taurinus; eb, Equus burchellii; hn, Hippotragus niger; bb, Bison bison; tq, Tetracerus quadricornis; eg, Equus grevyi; ke, Kobus ellipsiprymnus; ab, Alcelaphus buselaphus; cd, Cervus duvaucelii.
Los dos primeros componentes principales explican el 56.65% de la varianza observada entre las variables de microdesgaste consideradas. Con base en los factores de carga del ACP, las variables que contribuyen mayoritariamente al primer componente principal son los rayones gruesos (-0.53) y los rayones finos (0.46) y, en menor medida, las fosas grandes (0.39) (Tabla 6). Las muestras de Hidalgo, Puebla y Oaxaca tienen un registro positivo en el primer componente que se relaciona a que tienen valores relativamente altos de rayones gruesos y relativamente bajos en las otras dos variables.
Tabla 6 Eigenvalores y factores de carga de los componentes principales resultantes del Análisis de Componentes Principales de los datos de microdesgaste en muestras de Mammuthus columbi del Pleistoceno tardío de Hidalgo, Puebla y Oaxaca, centro y sur de México. Abreviaturas de las variables de microdesgaste en la Tabla 5.
| CP 1 | CP 2 | CP 3 | CP 4 | CP 5 | CP 6 | CP 7 | |
| Eigenvalor | 2.36672 | 1.3189 | 1.07894 | 0.901703 | 0.638129 | 0.44299 | 0.252627 |
| % varianza | 33.81 | 18.841 | 15.413 | 12.881 | 9.1161 | 6.3284 | 3.609 |
| Factores de carga | |||||||
| PR | 0.36743 | -0.57273 | 0.10318 | 0.075807 | 0.50448 | 0.090804 | 0.50778 |
| PF | 0.29064 | 0.67547 | -0.25796 | -0.13331 | 0.11441 | -0.2375 | 0.55269 |
| RC | 0.18795 | 0.39446 | 0.49539 | 0.63731 | 0.31591 | 0.099662 | -0.21857 |
| FG | 0.39115 | 0.041201 | -0.65842 | 0.10222 | 0.14008 | 0.51021 | -0.34845 |
| G | 0.30337 | 0.14843 | 0.4284 | -0.73778 | 0.19777 | 0.22083 | -0.26498 |
| rg | -0.53415 | 0.1844 | 0.070369 | -0.036933 | 0.081778 | 0.7262 | 0.3746 |
| rf | 0.46353 | -0.048906 | 0.2354 | 0.11912 | -0.75312 | 0.29829 | 0.23869 |
En el segundo componente, las variables que contribuyen mayoritariamente son el promedio de fosas (0.67), el promedio de rayones (-0.57) y en menor medida los rayones cruzados (0.39) (Tabla 6). La muestra de Puebla está más cercana al origen del segundo componente, lo cual se relaciona a que tiene un menor número promedio de fosas y un mayor número promedio de rayones en comparación a las muestras de Hidalgo y Oaxaca (Tabla 6).
Por su parte, el análisis de funciones discriminantes clasifica correctamente a las muestras de Hidalgo y Oaxaca con los ungulados de hábitos mixtos (estacional-regional) con una probabilidad del 75 y 73% respectivamente. La muestra de Puebla se clasifica en ese mismo grupo dietario con una probabilidad del 57% y en los pacedores con una probabilidad del 35%.
La proporción del número promedio de fosas y número promedio de rayones (PF/PR) de las muestras de Hidalgo (PF/PR = 0.67) y de Oaxaca (PF/PR = 0.61) está en el límite inferior del morfoespacio de los pacedores actuales, mientras que la muestra de Puebla está incluida en ese morfoespacio (PF/PR = 0.51). La especie pacedora actual más cercana a las tres muestras de Mammuthus columbi consideradas en este estudio es la cebra de las planicies (Equus quagga = E. burchelli) (Figura 10).

Figura 10 Gráfico del número promedio de rayones y número promedio de fosas en muestras de Mammuthus columbi del Pleistoceno tardío de Hidalgo (1), Puebla (2) y Oaxaca (3), muestras procedentes de Carolina del Sur (4), Texas (5), Nebraska (6) (Rivals et al., 2012) y centro-occidente de México (Michoacán-Guanajuato) (7) (Gutiérrez-Bedolla, 2014), así como de ungulados actuales con hábitos alimentarios típicos (Solounias y Semprebon, 2002). Abreviaturas de las especies actuales como en la Figura 9. Ramoneadores folívoros, círculos negros; ramoneadores frugívoros, círculos blancos; hábitos mixtos estacionales, rombos blancos; hábitos mixtos comidapor-comida, cuadros negros; pacedores, triángulos blancos.
6. Discusión
6.1. CARACTERIZACIÓN TAXONÓMICA
Los elementos mandibulares, dentales y postcraneales estudiados presentan la siguiente combinación de caracteres que permiten referirlos a Mammuthus: (1) mandíbula con proceso mentoniano bien desarrollado; (2) molariformes formados por múltiples láminas de esmalte, gruesas (comúnmente con un grosor ≥ 2 mm), paralelas entre sí y unidas por cemento (= lamelas); (3) el margen dorsal del atlas está considerablemente aplanado; (4) el cuerpo vertebral de las vértebras cervicales está comprimido antero-posteriormente; (5) las vértebras torácicas tienen apófisis espinosas altas y robustas, apófisis transversas extendidas, así como cuerpo vertebral masivo; (6) el margen ventral del cuerpo vertebral de las vértebras lumbares es redondeado; (7) el proceso espinoso medio de la escápula está ubicado a la mitad y a lo largo de la espina escapular, mientras que el cuello escapular es notablemente estrecho y la cavidad glenoidea es rectangular; (8) el astrágalo en vista anterior, está considerablemente comprimido dorsoventralmente (Olsen, 1972; Haynes, 1991; Hodgson et al., 2008).
La diferenciación entre las especies de Mammuthus se ha basado comúnmente en la configuración de los últimos molares (M3/m3), dado que estos elementos se usan durante más tiempo en comparación a los molariformes precedentes y son los que frecuentemente se recuperan en las muestras fósiles (Maglio, 1973; McDaniel y Jefferson, 2006a; Hodgson et al., 2008). Cabe señalar que la configuración de los elementos mandibulares de Hidalgo (UAHMP-4220) y Oaxaca (UMPE-617) es semejante a la que se observa en mandíbulas de Mammuthus columbi con las siguientes características: (1) cuerpo de la mandíbula robusto; (2) proceso mentoniano proyectado hacia abajo; (3) rama ascendente corta y robusta; así como (4) proceso coronoides ubicado anterior al margen posterior de los molariformes inferiores (Gillette y Madsen, 1993; McDaniel y Jefferson, 2003).
Los molares superiores (M3) e inferiores (m3) terceros de la muestra estudiada presentan las siguientes características típicas de Mammuthus columbi: (1) molariformes grandes, relativamente angostos, con una anchura entre 75 a 108.5 mm (anchura promedio de los elementos estudiados de alrededor 80 mm); (2) dientes hipsodontes (índice de hipsodoncia comúnmente mayor a 2); (3) grosor del esmalte promedio cercano a 3 mm (grosor del esmalte promedio de la muestra estudiada de 2.8 mm); (4) esmalte fuertemente crenulado; y (5) frecuencia lamelar de 5 a 7 (Arroyo-Cabrales et al., 2003; McDaniel y Jefferson, 2006a). En este sentido, los terceros molares (M3/m3) de M. meridionalis se distinguen por ser menos hipsodontes (índice de hipsodoncia ≤ 1), grosor del esmalte que suele ser ≥ 3 mm y tener una menor frecuencia lamelar (4 - 5) (Arroyo-Cabrales et al., 2003; McDaniel y Jefferson, 2006a), mientras que los pertenecientes a M. primigenius tienen un grosor del esmalte comúnmente ≤ 2 mm y una mayor frecuencia lamelar (≈ 9) (Harington et al., 2012).
La frecuencia lamelar e índice de hipsodoncia de UMPE-022 (M2/M3) (Tabla 3) son cercanos a los del fragmento de un M2 de Mammuthus columbi recuperado de la localidad CRISOBA industrial CEPAMISA al sur de la ciudad de Morelia, Michoacán; sin embargo, este último ejemplar tiene un mayor grosor del esmalte de 3.5 mm (García-Zepeda y Garduño-Monroy, 2006: tabla 1, p. 10). Los M3 catalogados como UMPE-029-2 y UMPE-467 son proporcionalmente más grandes que los molares terceros de CRINAH-2274 (fragmento de cráneo con M2-M3 izquierdos y M3D) de M. columbi de Sierra de Amozoc, Pleistoceno tardío de Puebla (Carbot-Chanona et al., 2017). Sin embargo, la frecuencia lamelar (6 - 7) y grosor del esmalte promedio (≈ 3 mm) de los elementos oaxaqueños y los del ejemplar poblano son similares.
La frecuencia lamelar (5 - 6) e índice de hipsodoncia promedio de los m1 (2.5) y m2 (2.0) de Hidalgo, Puebla y Oaxaca son semejantes a la de los molares UM148 (m1D) y UM149 (fragmento de m1), así como UM186 (fragmento de un m2) pertenecientes a Mammuthus columbi de la Cuenca de Cuitzeo, Pleistoceno tardío de Michoacán; sin embargo, los elementos michoacanos tienen un mayor grosor del esmalte (> 3 mm) (García-Zepeda y Garduño-Monroy, 2006: tabla 1, p. 10). La frecuencia lamelar (≈ 6), grosor del esmalte (2 - 3 mm) e índice de hipsodoncia (> 2 mm) de los m3 de Hidalgo, Puebla y Oaxaca son similares a la de los molares del ejemplar CRINAH-2275 (m2 y m3 derechos e izquierdos de un mismo individuo), perteneciente a Mammuthus columbi de Barranca El Morillo, Pleistoceno tardío de Puebla (Carbot-Chanona et al., 2017). Los m3 engastados en la mandíbula catalogada como UAHMP-4220 de Hidalgo son un 22% más grandes en longitud oclusal promedio (243.5 mm) en comparación a la de los molares terceros de CRINAH-2275 (186 mm).
La diferencia en tamaño, entre los M3 de Oaxaca y los m3 de Hidalgo con los M3/m3 de los ejemplares de Puebla, probablemente se explique por variación intraespecífica relacionada con la edad y/o sexo de los individuos que representan tales ejemplares, así como por la ponderación temporal de las muestras, dado que no son isócronas.
La configuración de los elementos mandibulares y de los terceros molares de Hidalgo, Puebla y Oaxaca es la que distingue a los de Mammuthus columbi. Los otros molariformes (superiores e inferiores) y elementos postcraneales asociados se refieren de manera parsimoniosa a la misma especie.
6.2. HÁBITO ALIMENTARIO DE ALGUNOS INDIVIDUOS DE MAMMUTHUS COLUMBI DEL CENTRO Y SUR DE MÉXICO
El patrón de microdesgaste de los individuos de Mammuthus columbi del Pleistoceno tardío de Hidalgo, Puebla y Oaxaca se caracteriza por presentar una importante representación de rasguños en comparación a fosas (PF/PR ≈ 0.6) (Figura 8). Esta condición se relaciona con un efecto en el que la abrasión (contacto diente - alimento) domina sobre la atrición (contacto diente - diente), lo cual a su vez sugiere un hábito alimentario pacedor (Solounias y Semprebon, 2002). Aunado a esto, los individuos muestreados exhiben una importante frecuencia de rayones gruesos (> 60%), condición que suele ser bastante común en algunos ramoneadores actuales, tal es el caso de los antílopes americanos (Antilocapra americana) y los camellos (Camelus dromedarius y C. bactrianus) (Solounias y Semprebon, 2002).
En particular, el individuo de Puebla (UAHMP4002) tiene un patrón de microdesgaste muy parecido al que caracteriza al elefante africano (Loxodonta africana), el cual se distingue por una alta frecuencia de rayones gruesos, rayones cruzados y gubias; sin embargo, en esta especie es común también una alta frecuencia de fosas grandes (Solounias y Semprebon, 2002). Los elefantes africanos tienen hábitos mixtos con una dieta que consiste en proporciones variables de follaje, plantas leñosas, arbustos y/o pastos dependiendo de la época del año (Codron et al., 2006; Owen-Smith y Chafota, 2012).
La información disponible sugiere que los individuos de Mammuthus columbi del Pleistoceno tardío de Hidalgo, Puebla y Oaxaca incorporaron en su dieta recursos suculentos (v. gr. hojas, brotes y/o ramas) y abrasivos (v. gr. pastos y/o partículas sedimentarias). Con base en esto, se propone un hábito alimentario mixto con énfasis en recursos abrasivos para los individuos estudiados. Esta interpretación es consistente con los resultados derivados del ACP y el de funciones discriminantes.
El estudio de contenido estomacal, contenido en coprolitos, microdesgaste y análisis de isótopos estables de carbono ha revelado que M. columbi tuvo una amplia flexibilidad alimentaria (Lister y Bahn, 2007). Con relación a esto, el análisis de microdesgaste en ejemplares de Mammuthus columbi del Pleistoceno de Nebraska, Carolina del Sur y Texas demostró que esta especie de mamut tuvo un comportamiento dietario variable, incluyendo individuos de hábitos mixtos, ramoneadores y pacedores (Rivals et al., 2012). La proporción del número promedio de fosas y número promedio de rayones en estas poblaciones estadounidenses (PF/ PR = 1.40) es mayor a la observada en las muestras estudiadas y se asocia a un efecto diferencial de atrición y abrasión común en organismos de hábitos mixtos. En este mismo sentido, la proporción de los ejemplares de Hidalgo, Puebla y Oaxaca es semejante a la que se observa en muestras de Mammuthus columbi de localidades ubicadas entre Michoacán y Guanajuato en el centro-occidente de México (PF/PR = 0.54) (Gutiérrez-Bedolla, 2014) (Figura 10). Esto sugiere que varias poblaciones de M. columbi del centro y sur de México tuvieron un comportamiento dietario con mayor énfasis en materiales abrasivos en comparación a poblaciones de la misma especie de las Grandes Planicies y la Planicie Costera del Golfo y el este de los Estados Unidos. Esto a su vez podría relacionarse a que los conjuntos mexicanos hayan frecuentado sitios más áridos y abiertos.
Durante el siglo pasado y principios de este siglo, los mamuts del Pleistoceno tardío de México se consideraron herbívoros con una dieta que consistía principalmente en el consumo de pastos; esta interpretación alimentaria se relacionaba con la morfología típica de los molares en estos proboscídeos (Ferrusquía-Villafranca, 1978; Morett et al., 2003). Recientemente, la caracterización del hábito alimentario en numerosas poblaciones de Mammuthus columbi del Pleistoceno tardío de México, con base en el análisis de isótopos estables de carbono en esmalte dental, ha demostrado que esta especie tuvo un amplio espectro trófico que incluía en su dieta una proporción variable de plantas tipo C3 y C4 (Pérez-Crespo et al., 2009, 2012, 2014, 2016; Bravo-Cuevas et al., 2017). Por su parte, el estudio de microdesgaste y mesodesgate en ejemplares de M. columbi del Pleistoceno tardío de Michoacán-Guanajuato y Puebla respectivamente, condujo a considerar una dieta dominada por la abrasión asociada a un hábito pacedor (Gutiérrez-Bedolla et al., 2016; Carbot-Chanona et al., 2017).
La información sobre el comportamiento dietario de Mammuthus columbi en Norteamérica derivada de evidencia directa, análisis de las relaciones isotópicas de carbono y evaluación de patrones de desgaste dental, indica que esta especie de mamut tuvo una condición alimentaria generalista, comparable a la que se observa en el actual elefante africano Loxodonta africana. Esta condición les permitió explotar una amplia gama de ambientes, lo cual a su vez se asociaría a su amplia distribución geográfica a lo largo del subcontinente norteamericano, alcanzado áreas de Centroamérica (Lange, 2002).
Los resultados de microdesgaste en los individuos de Mammuthus columbi considerados en este estudio corroboran la amplia adaptabilidad dietaria de esta especie de mamut. Asimismo, la condición alimentaria mixta evidencia de manera indirecta la presencia de sitios abiertos (tal vez cubiertos por pastos y arbustos) y zonas de vegetación cerradas (áreas arboladas) en lo que ahora es parte del sureste de Hidalgo, la Cuenca de Valsequillo en Puebla, así como en la Mixteca Alta Oaxaqueña durante la segunda mitad del Pleistoceno. Estudios paleoecológicos en asociaciones de mamíferos cuaternarios del centro y sur de México, muestran que ambientes mixtos de vegetación caracterizados por zonas arboladas y pastizales fueron comunes en esas regiones del país en el Pleistoceno tardío (Bravo-Cuevas y Jiménez-Hidalgo, 2018).
7. Conclusiones
El estudio de elementos craneales y postcraneales de mamut procedentes de los estados de Hidalgo, Puebla y Oaxaca permitió identificarlos taxonómicamente como pertenecientes a Mammuthus columbi. Con base en los elementos dentales se estimó que la muestra representa entre 15 y 16 individuos; siete u ocho en Oaxaca, cuatro en Hidalgo y cuatro individuos en Puebla. La categoría de edad mejor representada es la de más de 30 años, seguida de los adolescentes con un intervalo de edad entre tres y diecisiete años. Al menos cuatro de los ejemplares podrían ser machos, lo que indica la presencia de al menos un macho adulto y sexualmente activo en cada una de las muestras estudiadas, si se considera que M. columbi alcanzó dicho estado sexual a una edad similar a la de Elephas maximus.
El análisis de microdesgaste de algunos individuos mostró un mayor número de rayones que de fosas, así como una frecuencia elevada de rayones gruesos. Dicho patrón de microdesgaste refleja un hábito alimentario mixto con una ingesta elevada de recursos abrasivos. Este hábito alimentario mixto sugiere la presencia de zonas con vegetación abierta y vegetación de tipo cerrado en lo que ahora son los estados de Hidalgo, Puebla y Oaxaca durante el Pleistoceno tardío.

