











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink1. Introduction
During the mid-Cretaceous, particularly in the Albian through Cenomanian, widespread carbonate platform development took place throughout the tropical-subtropical shallow-water environments of the Tethys (Schroeder and Neumann, 1985, p. 5). Benthic foraminifers and algae are the most important groups that inhabited this milieu. In Mexico, during the Albian/Cenomanian, the Valles-San Luis Potosí Platform (VSLPP) was part of a wide carbonate platform system that bordered the ancestral Gulf of Mexico (Salvador, 1991). This platform experienced the evolution of a rich and diverse benthic foraminiferal assemblage now contained in the El Abra Limestone.
In this study, we analyzed calcareous samples derived from the Cenomanian El Abra Formation containing an abundant and well-preserved benthic foraminiferal assemblage that also includes algae, corals, echinoderms, and annelids.
The El Abra Formation is renowned for its invertebrate fossils, primarily Albian rudists, which have been analyzed by numerous authors ( e.g.Adkins, 1930; Coogan; 1973; Alencáster, 1998; Alencáster and García-Barrera, 2008).
The Albian benthic foraminifers from the El Abra Formation were documented by Bonet (1956), and Tavitas and Solano (1984) from the eastern part VSLPP. They reported an assemblage composed of Orbitolina (Mesorbitolina) texana, Dictyoconus wal-nutensis (Carsey), Coskinolina sunnilandensis (Maync), Pseudonummoloculina heimi (Bonet), and Dicyclina schlumbergeri (Munier-Chalmas).
We studied the benthic foraminiferal community from the western part of the VSLPP. This association has been dated as middle-late Cenomanian and is very different from the well-known foraminiferal association of the eastern part composed mostly of orbitolinids (Omaña et al., 2016).
The upper part of the El Abra Formation in the area of study represents the last shallow-water deposit preceding the VSLPP, which was drowning at the Cenomanian/Turonian boundary interval (Basañez et al., 1993; Omaña et al., 2012). It is overlain by a succession of bioclastic packstone and pelagic wackestone bearing planktic foramini-fers and pithonellids (Soyatal Formation).
The drowned episode in the Cenomanian/Turonian (C/T) interval at the global level is linked with rising sea level (Haq et al., 1987; Haq, 2014; Hallam, 1992; Miller et al., 2005; Gale et al., 2008), and widespread flooding of Mediterranean platforms (Drzewiecki and Simó, 1977; Caus et al., 1997; Hart et al., 2005; El-Sabbagh et al., 2011; Korbar et al., 2012).
The start of the demise of the carbonate platforms occurred during the latest Cenomanian (Whitei-nella archaeocretacea Zone), with a major biological change that affected benthic organisms such as rudists and larger benthic foraminifera.
Philip and Airaud-Crumière (1991, p. 117) documented that the benthic foraminifera were affected in two steps; the genera Praealveolina, Chrysalidina, Pseudocyclammina, and Pseudolituonella disappeared at the top of Geslinianum Zone. During the late Cenomanian (Juddii Zone) the benthic forami nifera were scarce and poorly diversified miliolids, textularids, and trochamminids. At second step occurs when a major extinction affected the benthic foraminifera at the C/T boundary.
Parente et al. (2008) also reported a two-step pa ttern of extinction of larger foraminifers in the upper Cenomanian in shallow-water carbonates of the southern Apennines (Italy). They stated that “the first step eliminated the alveolinids, the most extreme k strategists, reducing the diversity of the larger foraminifera to less complex forms such as Chrysalidina, Pseudolituonella and Pseudoraphydionina; when the high productivity persisted only r-opportunist planktic foraminifers predominated while the rise of the mesotrophic threshold triggered the second step extinction of larger foraminifera.” However, in the present study, the C/T boundary interval is characterized by the disappearance first of larger forms such as Pseudolituonella, Daxia, Merlingina, Hemicyclammina, Dicyclina, and Cuneolina. Later, only forms such as Nezzazatinella picardi, Prae-chrysalidina, rotalids, miliolids, and algae remained, this interval is overlain by a deposit with abundant pithonellids, and r and r-k strategist planktic foraminifers with simple morphologies, such as Hedber-gella, Heterohelix, Globigerinelloides, and Whiteinella as a result of flooding of the platform.
The objectives of the work are a) to describe and illustrate the benthic foraminifers identified from the upper part of the El Abra Formation as a contribution to the knowledge of the faunal assemblage of this unit in Mexico and of the western part of the Tethys Realm; b) to provide information about the distribution of the larger foraminifers before their demise, caused by the flooding of the VSLPP; c) to constrain the age of the El Abra Formation using the stratigraphic value of the benthic foraminifers, comparing the biostratigraphic standards based on them in the peri-Tethyan area; moreover, supporting it from the planktic age of the overlying unit; and d) to integrate the paleoecological data of the larger foraminifers and the microfacies study to infer information about the paleoenvironment.
2. Geological setting
The samples were collected east of the city of San Luis Potosí (Figure 1). This area belongs to the western part of the VSLPP, which formed in association with the opening of the Gulf of Mexico. According to Salvador (1991), the Early Cretaceous was a time of significant tectonic stability in the Gulf of Mexico Basin. During this period, the influx of terrigenous sediment decreased and the stable shelves, ramps, and platforms bordering the deep central part of the basin became the site of widespread carbonate deposition, particularly during the Albian, when carbonate deposition extended from the Bahamas along the northern rim of the Gulf, across Mexico to the Yucatan Peninsula, and to various Caribbean localities (Scott, 1990). Along the western flank of the basin in east-central Mexico, carbonate platforms were more locally restricted, such as the VSLPP (Figure 2), which remained active until the Cenomanian. Wilson and Ward (1993, p. 42) stated that the VSLPP is one of “the largest isolated carbonate platforms (200 by 300 km) located on the western side of the Gulf of Mexico. It initiated in the Early Cretaceous, but reached its maximum area in the Albian, when it evolved a rimmed shelf margin. The platform apparently formed along the south-western flank of a north-northwest-trending tectonic ridge of Precambrian gneiss and Upper Paleozoic schist that is exposed at Miquihuana, 70 km southwest of Ciudad Victoria on the state line between Nuevo León and Tamaulipas”.
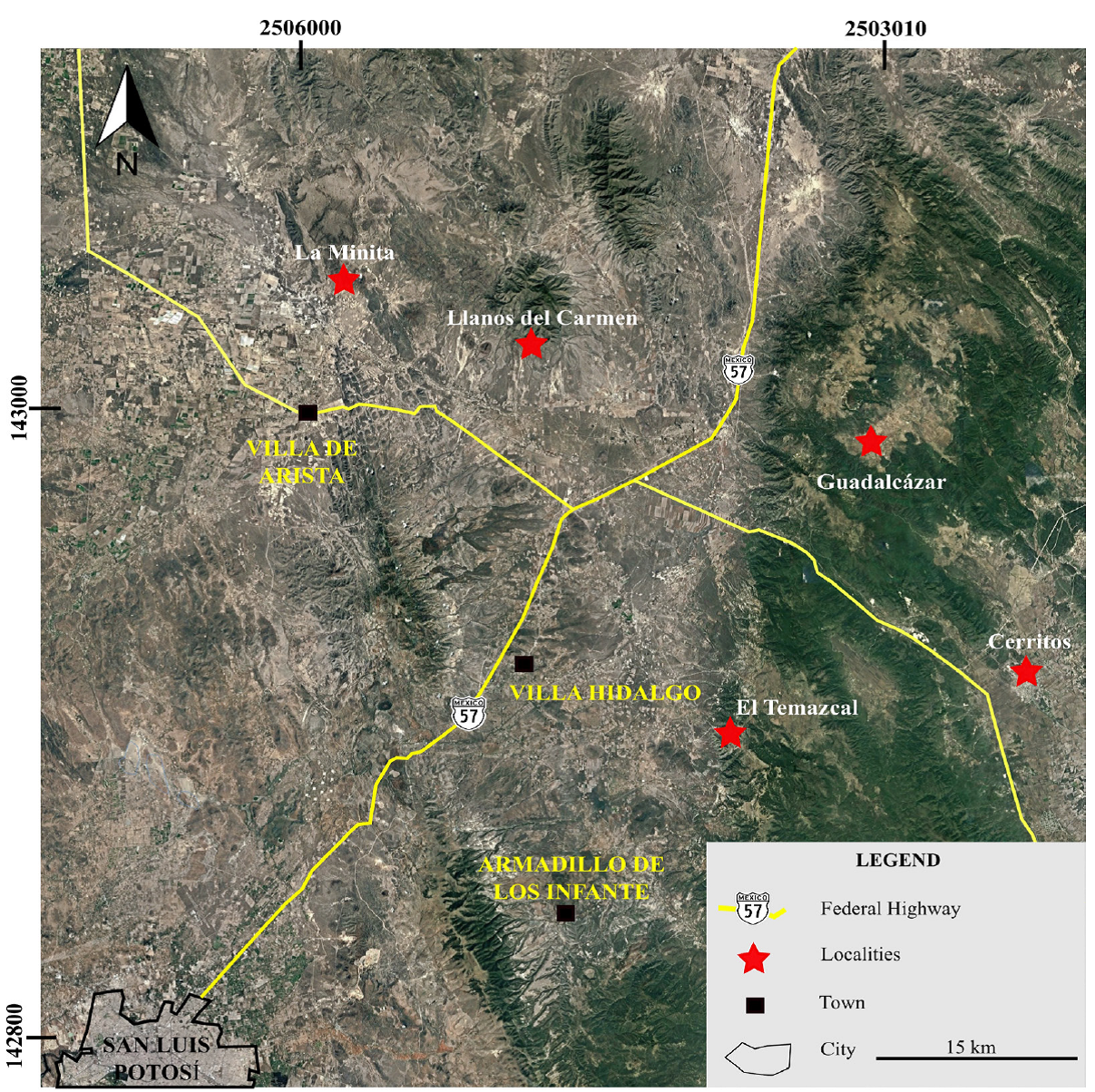

Figure 2 Geological map showing the El Abra and Soyatal Formations (modified from Labarthe et al., 1982; López-Doncel, 2003).
Recently, Venegas-Rodríguez et al. (2009) stated that the VSLPP overlies Toarcian rocks dated by detrital zircon from the El Alamito locality at W Río Verde, San Luis Potosí. It is interpreted by these authors as a deposit from the paleo-Pacific margin occuring at same time as the opening of the Gulf of Mexico.
The VSLPP now lies within the Sierra Madre Oriental fold-thrust belt (Suter, 1984; 1987). Its eastern shelf margin is exposed in quarries along the Cuesta de El Abra near Valles and Antiguo Morelos (km 541-546 of the San Luis Potosí-Tampico railway). Part of its western side can be seen along road cuts 30 km east of the city of San Luis Potosí. The stratigraphic framework for the VSLPP has been described by Cserna and Bello-Barradas (1963), Carrillo-Bravo (1971), Suter (1990), Basañez (1993), Wilson and Ward (1993), and Torres-Hernández (1994). The depositional process has been recorded by Enos and Stephens (1993), Aguayo (1998), and López-Doncel (2003).
3. Materials and Methods
The studied material comes from the upper part of the El Abra Limestone (Figure 2). It was collected from four sections that ranged from 40 to 100 m thick located at the western part of the VSLPP: Llano del Carmen (LLC), Guadalcázar (G), Cerritos (C), El Temazcal (ET), as well as in the La Minita outcrop (LM).
The samples were collected at an average interval of 5 to 10 m. The 150 thin sections were prepared both parallel and perpendicular to the stratification for foraminiferal and microfacies examination. The faunal preservation is good, which permitted precise identification; the biostratigraphic scheme is based on the benthic and planktic foraminiferal stratigraphic ranges. In addition, a microfacies study was carried out to reconstruct the paleoenvironment.
4. Results and Discussion
4.1. Lithofacies and microfacies
The El Abra Formation displays two facies, the Taninul and El Abra, which are stratigraphically correlated (Aguayo, 1998). The Taninul facies is a thick to massive skeletal bioclastic limestone mostly composed of rudists, foraminifers and algae (Figure 3a to 3c). The El Abra facies is a light gray limestone rich in miliolids and ostreids (Figure 3e and 3f).

Figure 3 Microfacies of the upper part of the El Abra Formation. Scale bars 100 μm. a: Packstone-grainstone (Sample ET-2); b: Grainstone poorly classified with miliolids (Sample LLC-4); c: Worn algal packstone (Sample C-9); d: Calcisphere-rich packstone-wackestone (Sample C-10); e: Wackestone-packstone with a great abundance of miliolids and thin-shelled ostreids oriented in a parallel arrangement (Sample -LM-1); f: Benthic foraminiferal wackstone-packstone (Sample LM-5).
4.1.2. THE taninul facies
The following sections were measured, sampled, and described from the Taninul facies.
4.1.2.1. Llano del Carmen Section (LLC)
The Llano del Carmen section (LLC) currently crops out north of the city of San Luis Potosí (coordinates 22º45ʹN, 100°40ʹW). It consists of massive dark-gray limestone of the El Abra For-mation and is ~100 m thick (Figure 4).

The petrographic analysis indicates a peloidal packstone-grainstone with high foraminiferal species richness that includes Pseudolituonella reicheli, Daxia cenomana, Hemicyclammina sigali, Merlingina cretacea, Pseudocyclammina sp., Dicyclina schlumbergeri, Cuneolina pavonia, Cuneolina parva, Nezzazata gyra, N. simplex, Nezzazatinella picardi, Spiroloculina cenomana, Peneroplis parvus, and Minouxia inflata. The algae Bouenia sp. and encrusting Lithocodium aggregatum are present. The components of more than 60% of the content can be recognized. They are embedded in microsparitic cement, densely packed in a matrix-supported fabric. Most of the components are coated with micrite, suggesting moderate transport into a relatively high-energy environment. The lithology and faunal-algal assemblage indicates a deposit in a normal-marine open platform not subjected to significant restriction in water circulation. It is important to note that rudist Hippurites resectus is also found in this association.
Other microfacies consist of a coated, poorly sorted foraminiferal grainstone consisting of miliolids and other forms such as Peneroplis parvus, and algae; the grains and microfossils are strongly micritized (Figure 3b).
4.1.2.2. The guadalcázar section (G)
The Guadalcázar section (G) is located NE of the city of San Luis Potosí (22°64ʹN, 100°38ʹW). The studied succession belongs to the upper part of the El Abra Formation that crops out in the Aguaje de los García stream. It consists of a gray calcareous deposit ~50 m thick of the Taninul Facies (Figure 5). The lithofacies is dominated by packstone and subordinate grainstone with the same foraminiferal assemblage as the Llano del Carmen site but containing algae, such as Salpingoporella hasi and Clypeina sp.

The presence of sparitic cement represents an increase in energy in practically the same microfacies as at the Llano del Carmen site. This microfacies is interpreted as Standard Microfacies Type (SMF) 18 (Flügel, 2004) of Facies Zone 7 to 8 (Wilson, 1975) in the platform (open marine). The El Abra limestone is unconformably overlain by the Soyatal Formation, which here is 15 m thick.
4.1.2.3. The cerritos section (C)
The Cerritos section (C) was measured on the San Luis Potosí-Cerritos road (22°43ʹN, 100°28ʹW). It consists of 45 m of massive gray limestone from the upper part of the El Abra Formation of the Taninul Facies, and it is conformably overlain by the Soyatal Formation (Figure 6).

In the El Abra Formation we recognized the following 2 microfacies:
Microfacies 1-Benthic foraminiferal packstone- grainstone rich in pellets and pseudopellets. The percentage of components locally exceeds 50%, forming a grain-supported fabric. The ground-mass, mostly sparry cement, shows small remains of an original micritic matrix. The microfacies type could be comparable to SMF 18 “bioclastic grainstones and packstones with abundant benthic foraminifera or calcareous green algae” (Flügel, 2004).
The depositional environment of this microfacies suggests a shallow marine environment above the normal wave base, within the euphotic zone in the open marine interior platform, corresponding to Facies Zone 7 of Wilson (1975). The fossil assemblage indicates precipitation in shallow subtidal zones with normal salinity, stable temperature conditions, and good oxygenation of the seawater.
Microfacies 2-Worn algal packstone, within a fine-grained matrix containing a larger embedded amount of algae (Figure 3c). The foraminiferal community is reduced to small forms of Nezazzatinella picardi, miliolids, and rotalids. This change could be related to an increase in nutrient supply. This microfacies is similar to SMF 10, suggesting an open sea shelf environment of Facies Zone 7 of (Wilson 1975). The main components have been transported from high-energy to low-energy environments, and a variation in the water energy can be inferred from the increase in size of the bio clasts and groundmass grains (microsparite). This microfacies corresponds to Facies Zone 7 (interior platform, open marine environment).
Up-section, the lithology changes to marly lime-stone of the hemipelagic-pelagic Soyatal Formation represented by the next microfacies:
Microfacies 3-Pithonellid-rich packstone-wackestone, this microfacies contains a bloom of pithonellids embedded in a micritic matrix that varies locally in microsparitic cement (Figure 3d). The population of pithonellids can be up to more than 50% of the components in a mud to grain-supported fabric. In addition to the pithonellids, r and r-k strategist planktic foraminifera are also present, including Muricohedbergella delrioensis, M. planispira, Heterohelix moremani, Heterohelix reussi, Macroglobigerinelloides caseyi, Whiteinella archaeocretacea, W. cf. W. aprica, W. baltica, W. brittonensis, and W. paradubia. The r-strategists are small to medium-sized species with unornamented tests and simple morphologies that have high reproductive potential and are adapted to eutrophic or unstable environments, and morphotypes as Whiteinella that connect the r and k morphogroups and are called r/k-intermediate species (Gebhardt et al., 2010). Small benthic foraminifera Dorothia sp., Gavelinella sp., Lingulogavelinella sp., and roveacrinids were identified. No microlamination or orientation of the components is recognizable; rather, it resem bles an unsorted accumulation of microfauna in the micritic groundmass. This microfacies could be similar to types SMF 2 and 3 Flügel (2004) (wackestone to packstone with pithonellids and foraminifers) and a Facies Zone 2 to 3 of Wilson (1975) (deep shelf), but its depositional environ ment is certainly evidence of an initial drowning event.
4.1.2.4. The el temazcal section (ET)
The analyzed section was measured in the Cerro Quemado locality (22°38ʹN, 100°50ʹW). It is composed of 40m of gray, massive limestone corresponding to the El Abra Formation, and is unconformably capped by a reddish calcareous shale unit.
The limestone in this locality has packstone-grainstone texture and locally the component exceeds 60%, forming a typical grain-supported fabric. The foraminiferal association is like that of the other sections, but the algae Neomeris cretacea is present. The standard microfacies type and the facies zone are similar to those at the other localities, which include SMF 8 to 10 in the Facies Zone 7 (Flügel, 2004; Wilson,1975).
4.1.3.1. The la minita outcrop (LM)
At this locality (22°78 5ʹN, 100°85ʹW), we collected 8 samples of cream to light-gray limestone of the El Abra facies (Figure 7).

The La Minita outcrop displays several examples of cream limestone of the El Abra facies (40 m thick). Two microfacies are defined: 1) a wacke-stone-packstone with reduced diversity but a great abundance of miliolids and thin-shelled ostreids, which are well-oriented in a parallel arrangement (Figure 3e). The presence of benthic foraminifera suggests shallow-water settings and the arrangement points to some control by the current. The components are embedded in a micritic matrix with a mud-supported fabric. These characteristics are comparable to SMF 8 (Flügel, 2004) of Facies Zone 7 in the interior platform; and 2) another that shows a benthic foraminifer-rich wackstone-packstone with a higher diversity of species including Pseudonummoloculina heimi, Ovalve-olina maccagnoae, Dicyclina schlumbergeri, and Cuneolina pavonia (Figure 3f). The ground mass varies from micritic up to microsparitic and the fabric is much denser (grain-supported) in comparison with the first microfacies. This microfacies can be classified as SMF 10 to 11 of Facies Zone 7 in the interior platform with moderate energy.
4.2. Age
Cretaceous larger benthic foraminifera are stratigraphically significant in shallow-water environments, where they are the most prominent marker fossils when other fossils such as the ammonites and planktic foraminifera are absent (Schroeder and Neumann, 1985).
4.2.2. Nezzazatinella picardi
Interval Zone, which overlies the Pseudolituonella reicheli Assemblage Zone which, is characterized by a change in the benthic foraminiferal assemblage in which the larger foraminifera Pseudolituonella, Pseudocyclammina, Hemicyclammina, Daxia, and Merlingina disappear, reducing the foraminiferal community to forms of Nezzazatinella picardi, Rotorbinella mesogeensis, Praechrysalidina sp., and miliolids. A proliferation of gymnocodaceans, such as Permocalculus, is observed. This association later also disappeared when the platform was drowned.
The most important foraminifer in the studied material is Pseudolituonella reicheli, which has a short range and is considered to be of mid-late Cenomanian age.
Schroeder and Neumann (1985) indicated that P. reicheli is a species mainly of Cenomanian and possibly Turonian age; however Saint-Marc (1974) assigned the taxon a long stratigraphic range spanning from the mid-Albian to Turonian in Lebanon. In Italy, Luperto-Sinni (1966) assigned a late Cenomanian age to P. reicheli; Sartoni and Crescenti (1962) also found the species at the top of the Cenomanian (Cisalaveolina fraasi Zone). Chiocchini et al. (1979, 1984) stated that this foraminifer is limited to the upper Cenomanian rocks; this author used P. reicheli as a zonal marker. In France, P. reicheli is reported from the Cenomanian in W Aquitaine (Neumann 1962, 1965; Saint-Marc, 1966). Bilotte (1984) considered this species typical of the Cenomanian. Husinec et al. (2000) in Croatia andDecrouez (1977) in Greece reported P. reicheli from the mid-late Cenomanian. In Israel, it is recorded in the late Cenomanian and Turonian (Reiss, 1957). In Iran, Bozorgnia and Banafti (1964), and Sampó (1969) recorded this species in the Cenomanian. Schulze et al. (2004) indicated that P. reicheli only occurs in the Cenomanian in west central Jordan. In Turkey, Sari et al. (2009) used this microfossil as a late Cenomanian zonal marker. Recently, Frijia et al. (2015, p. 129) indicated that this species disappears at levels that can be correlated by carbon isotope stratigraphy to latest Cenomanian (Neocardioceras juddi ammonite zone); however, in the studied material we recorded the complete flooding of the platform in this time with deposits of opportunist planktic foraminifers and pithonellids (Whiteinella archaeocretacea Zone).
In Mexico, Aguilera-Franco (2003) situated Pseudo-lituonella reicheli in the Pseudorhapydionina dubia Zone in the Guerrero-Morelos Platform in upper middle-upper Cenomanian. In the Chiapas region, Michaud (1987) regarded this species as middle- late Cenomanian.
Another important species found in the material examined is Daxia cenomana, which determines a mid-late Cenomanian age. This species was described by Cuvillier and Szakall (1949) from Audignon in France; they indicated that this genus appears to be restricted to the Cenomanian. It is considered a mid-Cenomanian species by Neumann (1965) and Moreau (1996) in the Charente Maritime region. In Spain, Barroso (2004) found Daxia cenomana from the Burgos Province, dating it as mid-Cenomanian. Gräfe (2005) recorded D. cenomana in the Cenomanian and Turonian from the Basque-Cantabrian Basin. Caus et al. (2009) regarded this species as early-mid Cenomanian age in the Iberian Range. Ismail et al. (2009) document the occurrence of D. cenomana from the Cenomanian in Egypt.
Another form identified in the examined samples is Merlingina cretacea, which confirms the mid-late Cenomanian age. It was described from Negev, Israel by Hamaoui and Saint-Marc (1970) as from the Cenomanian. Schroeder and Neumann (1985) regarded the species as restricted to the middle part of the Cenomanian and the base of the Turonian. Caus et al. (2009) found this species in mid-upper Cenomanian rocks in the Iberian Peninsula.
Other conspicuous form in the studied samples is Hemicyclammina sigali, which is also considered mid-late Cenomanian, taking into account that it is associated with Pseudolituonella reicheli.
Hemicyclammina sigali was described for the first time by Maync (1953). He assigned it to the middle Cenomanian age. Berthou (1984a) recorded the occurrence of H. sigali restricted to the Cenomanian in the western Portuguese Basin. Later, Hart et al. (2005) studied a carbonate succession to outcrops along the valley of the Rio Mondego in the Lusitanian Basin, they stated that H. sigali is a particularly useful Cenomanian marker in many Tethyan areas. Bignot and Poisson (1974) recorded this species in the Cenomanian in the Katran, Dağ area, Turkey. Shirazi (2009) documented the presence of H. sigali in Iran in the mid-late Cenomanian. It has also been recorded in Syria (Mouty et al., 2003). Berthou and Bengtson (2007) considered H. sigali to be a reliable Cenomanian marker from the Sergipe Basin in Brazil.
An abundant form in the studied samples is Dicyclina schlumbergeriMunier-Chalmas (1887), associated with the mid-late Cenomanian species.
In Mexico, D. schlumbergeri is very common in Albian and Cenomanian strata; it has been reported from east of the VSLPP (Bonet, 1956; Tavitas et al., 1984) as well as in the Morelos Formation in the Guerrero-Morelos Platform (Aguilera-Franco and Allison, 2005), and in the Sierra Madre Formation in the Chiapas Range (Michaud, 1987). This species has been found in the Orizaba Formation in the Veracruz Basin (Sohl et al., 1991) and the Tepalcatepec Formation in Jalisco and Colima (Ponce de León, 1983; Ornelas, 1984).
The usual occurrence of Pseudonummoloculina heimi was recorded in the material examined. The species was described by Bonet (1956) for the first time from the El Abra Canyon in the VSLPP. He assigned it an Albian/Cenomanian age. In Mexico, this species has a similar geographical and stratigraphical distribution to that of Dicyclina schlumbergeri. The species is recorded in Texas, Florida, and Louisiana in USA (Conkin and Conkin, 1958). It has also been recorded in Italy (Luperto-Sinni 1966 and Spalluto, 2012), in Slovenia by Koch et al. (1998), and in Croatia by Husinec et al. (2000); it is also registered in Turkey (Sari et al., 2009) and Syria (Ghanem et al., 2012) in middle-upper Cenomanian strata.
A common species found in the analyzed samples is Cuneolina pavonia d’Orbigny. This species is known from Albian to Cenomanian in Tethys strata (Arnaud-Vanneau and Premoli-Silva, 1995). Peneroplis parvus is a common species in the samples analyzed. According to Schroeder and Neumann (1985), this form ranges from the latest Albian to mid-Cenomanian. According to Fleury (1980) this species is late Albian to early Cenomanian age from the Varassova Mountains in Greece. In Croatia, P. parvus was determined to be in the late Cenomanian (Velić and Vlahović, 1994).
We assigned a mid-late Cenomanian age to Peneroplis parvus in the material studied and thus its stratigraphic distribution could be extended from that given in other publications, in which the species is considered as mid-Cenomanian age (De Castro, 1965; Schroeder and Neumann, 1985).
The extinction of all the benthic foraminifera occurred in the Cenomanian/Turonian boundary interval by eutrophication of shallow marine environments and the terrigenous supply, as has been recognized in our material.
The El Abra Cenomanian shallow-water carbonates are overlain by the hemipelagic Soyatal Formation with a Pithonella acme and the r and r-k strategist planktic foraminifers Muricohedbergella delrioensis, M. planispira, Heterohelix moremani, H. reussi, Whiteinella archaeocretacea, W. aprica, W. brittonensis, W. baltica, W. paradubia,W praehelvetica, and other small benthic foraminifera; for example, Lingulogavellinella, Gavellinella and textularids. This interval is recognized as the Whiteinella archaeocretacea Partial Range Zone, which has been regarded as latest Cenomanian/earliest Turonian, which is impor tant because it constrains the age assigned to the El Abra Limestone.
4.3. Paleoecology and paleoenvironment
In modern environments, benthic foraminiferal distribution patterns are controlled by temperature, light intensity, nutrient content, substrate, and hydrodynamic energy. It is inferred that they had a similar distribution in the Mesozoic and Cenozoic (Hohenegger, 2000, 2004; BouDagher-Fadel, 2008).
The larger symbiotic-bearing foraminifera are indicative of warm waters (Murray, 1991). Their depth distribution is related to light, which is the main factor because it is required by the symbionts. The influence of the symbionts on the shape of the foraminiferal test as well as the hydrodynamic factor provides an important tool for paleoenvironmental interpretation. Thus, detached life forms with robust spherical tests indicate current-swept reef environments while thin fragile tests with maximum surface-to-volume ratios will be found in quiet low-light environments (Haynes, 1965). This is supported by the modern assemblage study. Recent larger foraminifera may occur as encrusters, periphyton (free-living on seagrass and calcareous algae), or living on or in the sediment (epi-faunal, infaunal). In higher energy environments, encrusters and periphyton are the most common, while algal-film and sediment dwellers predominate under lower energy conditions (Hallock and Glenn, 1986; Murray, 2006).
Recent Peneroplis and Spiroloculina are known as epi-phytic forms (Langer, 1993) or living in sea grass (Hallock and Glenn, 1986).
In this study, benthic foraminifera, together with other biogenic components such as algae, as well as lithologic characteristics were used to reconstruct the paleoenvironmental conditions that existed during the El Abra deposit.
The El Abra Formation (Taninul Facies) in the studied successions, Llanos del Carmen, Cerritos and El Temazcal sections, is characterized by an abundant and diversified benthic foraminiferal assemblage, composed of Pseudolituonella reicheli, Hemicyclammina sigali, Daxia cenomana, Merlingina cretacea, Cuneolina pavonia, Cuneolina pavonia parva, Pseudocyclammina sp., Dicyclina schlumbergeri, Nezzazata gyra, N. simplex, Nezzazatinella picardi, and Peneroplis parvus. The algae recorded are Bouenia pyigmaea and Neomeris cretacea, as well as the encrusters Lithocodium aggregatum and Thaumatoporella parvovesiculifera.
The great variety and diversity of the benthic foraminifers, together with algae and invertebrates such as rudists, gastropods, echinoderms, and calcareous worm tubes, as well as the peloidal bioclastic packstone-grainstone texture of the rock, suggest a shallow-water environment within the euphotic zone on the open marine platform during stable salinity and temperature conditions, permitting this oligotrophic association to develop and flourish. The occurrence of large-sized discoidal and conical benthic foraminifers such as Dicyclina, Cuneolina, and the encrusting habit of Thaumatoporella parvovesiculifera and Lithocodium, indicate medium hydrodynamic energy.
There is a conspicuous change in the top of the El Abra Formation. The foraminiferal community is reduced to a few benthic foraminifers such as Nezzazatinella picardi, miliolids, and rotalids, and the algae Permocalculus irenae and P. budaensis. The abundance of algae as well as decreased diversity of benthic foraminifera indicates a marine deposit associated with changes in the nutrient supply to the platform that caused the larger sized for-aminifers adapted to oligotrophic conditions to disappear, reducing the assemblage to small forms, indicating a decrease of benthic-factory carbonate production (Schlager, 1981), which preceded the drowning of the platform. The Soyatal Formation overlies the El Abra limestone, which consists of a foraminiferal packstone-wackestone with an overabundance of pithonellids, r and r-k strategist planktic, and different benthic foraminifers such as gavellinelids, which indicate a deeper and eutrophic environment. These significant faunal and lithological changes are related to an early phase of the transgression that flooded the VSLPP in the latest Cenomanian (Whiteinella archaeocretacea Partial Range Zone).
A lateral change of facies of the El Abra Formation, termed the El Abra facies, is recorded in the La Minita outcrop, which is characterized by a cream limestone represented by a wackestone with abundant miliolids and ostreids adapted to survive within environments with varying salinities, these characteristics suggest a restricted shallow environment. Other microfacies is identified showing a larger diversity of species including Pseudonummo-loculina heimi and Ovalveolina maccagnoae, other alve-olinids, Dicyclina schlumbergeri and Cuneolina pavonia allow us to infer an intermediate semi-restricted shallow-water environment.
4.4. Systematic paleontology
The benthic foraminifers contained in the lime-stone samples of the El Abra Formation are abundant and well preserved. The new suprageneric agglutinated foraminiferal assignment is based on the classification proposed by Kaminski (2014) for the agglutinated foraminifera, and Pawlowski et al. (2013 partim).
The thin sections are housed in the Paleontology Collection of the Institute of Geology (Universidad Nacional Autónoma de México).
Class FORAMINIFERA d’Orbigny, 1826
Subclass Globothalamea Pawlowski Holzmann and Tyszka, 2013
Order Lituolida Lankester, 1885
Suborder Nezzazatina Kaminski, 2004
Superfamily Nezzazatoidea Hamaoui and Saint-Marc, 1970
Family Nautiloculinidae Loeblich and Tappan, 1985
Genus BiconcavaHamaoui, 1965
Type species. Biconcava bentoriHamaoui, 1965
Biconcava bentoriHamaoui, 1965

Figure 8 Mid-late Cenomanian foraminifera from the El Abra Formation (scale bar 200 μm). a: Biconcava bentori tangential section; b: Charentia sp. equatorial section (Sample G-2) (Sample LLC-1); c: Daxia cenomana equatorial section (Sample LLC-1); d: Pseudocyclammina sp. subaxial section (Sample LLC-2).
Biconcava bentoriHamaoui 1965, p. 14; Hamaoui and Saint-Marc, 1970, p. 304, pl. 18-21; Loeblich and Tappan, 1988, p. 71, pl. 55, figs. 1-4.
Description: Test planispirally coiled with three whorls, showing a rounded proloculus, the chambers growing slowly; curved sutures, but slightly curved in. The last whorl has 18 chambers.
Remarks: Loeblich and Tappan (1988) transferred Biconcava to Maycinidae due the absence of a rudimentary internal tooth plate described by Hamaoui and Saint-Marc (1970, p. 301). The former authors stated, “this genus is closest in appearance to Daxia but differs in being more evolute and in the aperture located in a vertical groove in the apertural face.”
Genus DaxiaCuvillier and Szakall, 1949
Type species: Daxia cenomanaCuvillier and Szakall, 1949
Daxia cenomanaCuvillier and Szakall, 1949

Figure 9 Mid-late Cenomanian foraminifera from the El Abra Formation (All scale bars 200 μm except f, g scale bar 500 μm. a: Daxia cenomana equatorial section (Sample ET-4); b: Nezzazatinella picardi equatorial section on the spiral and the umbilical side (Sample ET-4); c: Merlingina cretacea oblique section (Sample LLC-5); d: Nezzazata gyra oblique subequatorial section (Sample ET-4); e: Hemicyclammina sigali equatorial section (Sample LLC-1); f, g: Nezzazata simplex equatorial section on spiral and umbilical side (Sample LLC-1).
Daxia cenomanaCuvillier and Szakall, 1949, p. 8, pl. 2, figs. 4-6; Maync, 1952, p. 55, pl. 11, fig. 3; Schroeder and Neumann, 1985, p. 15, pl. 1, figs. 1-8.1.
Description: Test planispirally coiled; with a rounded proloculus followed by rhomboid-shaped chambers; the latest chambers are low and elongated with curved sutures. Some specimens in our material possess 12-16 chambers in the last whorl. The measured height varies from 500 to 700 µm; the diameter of the proloculus is 30 mµ.
Remarks: Daxia cenomana differs from Daxia minimaLaug and Peybernès (1979) of the Aptian (Pyrénées Basco-Béarnaises), in its larger size, thinner septa and sutures less depressed in equatorial section.
Occurrence: Daxia cenomana is considered a typical Tethysian species (Schroeder and Neumann, 1985), and has been recorded in France (Cuvillier and Szakall, 1949; Neumann, 1965; Bilotte, 1984), Spain (Calonge and Caus, 1996; Gräfe, 2005; Barroso, 2004; Caus et al., 2009), Portugal (Berthou, 1984b; Berthou and Lauverjat, 1979), and Egypt (Ismail et al., 2009; Anan et al., 2013).
VSLPP, it was recorded in the Llanos del Carmen, Cerritos, and Temazcal sections.
Family Nezzazatidae Hamaoui and Saint-Marc,1970
Subfamily Nezzazatinae Hamaoui and Saint-Marc, 1970
Genus MerlinginaHamaoui, 1965
Type species: Merlingina cretaceaHamaoui, 1965
Merlingina cretaceaHamaoui, 1965
Merlingina cretaceaHamaoui, 1965, p. 7, pl. 9, fig. 6; pl. 14, fig. 14; Hamaoui and Saint-Marc, 1970, p. 315, pl. 22-27; Schroeder and Neumann, 1985, pl. 1, figs. 1-8, p. 37, pl. 15, figs. 1-9; Michaud, 1987, pl. 12, figs. 1-12.

Figure 10 Mid-late Cenomanian foraminifera from the El Abra Formation. a: Cuneolina pavonia axial section (Sample G-5); b: Cuneolina pavonia axial section (Sample ET-1); c: Cuneolina parva axial section (Sample LLC-5); d: Cuneolina parva axial section (Sample ET-1); e: Cuneolina parva axial section (Sample LLC-5); f: Cuneolina parva axial section (Sample LLC-5); g: Dicyclina schlumbergeri axial section equatorial section showing the great proloculus (Sample G-5); h: Dicyclina schlumbergeri axial section (Sample LLC-5); i: Dicyclina schlumbergeri axial section subequatorial section (Sample LLC-2); j: Dicyclina schlumbergeri subaxial section (Sample G-5).

Figure 11 Mid-late Cenomanian foraminifera from the El Abra Formation (all scale bars 200 μm). a: Pseudolituonellareicheli axial section (Sample LLC-1); b: Minouxia inflata axial section (Sample G-1); c: Peneroplis parvus axial section (Sample ET-1); d: Archaecyclus cenomanianus (Sample LLC-5); e: Ovalveolina maccagnoae equatorial section (Sample LM-5); f: Pseudonummoloculina heimi subequatorial section (Sample LM-5); g: Spiroloculina cenomana axial section (Sample C-3); h: Ovalveolina maccagnoae subaxial section (Sample LM-5).
Description: The specimen illustrated is a triangular section with initial part planispiral, later uncoiled and finally flaring; broad and low chambers, subdivided by tooth plate slightly curved inward from the lateral borders of the apertural face, having two or three basal digitations attaching to the septum of the preceding chamber; sutures sinuous.
Remarks: Merlingina differs from Nezzazata and Biconcava by having an uncoiled stage in adult specimens, with rapid enlarging of spire, and sutures more complex as well as the aperture with a more complex tooth plate (Hamaoui and Saint-Marc, 1970).
Occurrence: Merlingina cretacea was described from a subsurface sample from Negev, Israel (Hamaoui and Saint-Marc, 1970). It has been recorded in Italy (Chiochini et al., 1984), Spain (Caus et al., 2009), Turkey (Sari et al., 2009), Oman (Philip et al., 1995), and Syria (Ghanem et al., 2012). In the Americas, this species was identified in Mexico from the Chiapas Platform (Michaud, 1987) and the Morelos Guerrero Platform (Aguilera-Franco, 2003), and in Peru (Jaillard and Arnaud-Vanneau, 1993).
VSLPP in the Llanos del Carmen section.
Genus NezzazataOmara, 1956
Type species: Nezzazata simplexOmara, 1956
Nezzazata simplexOmara, 1956
Nezzazata simplexOmara, 1956, p. 887, pl. 102, figs. 7-13; Hamaoui and Saint-Marc, 1970,p. 348, pl. 40 fig. 13; Schroeder and Neumann, 1985, p.33, pl. 11, figs. 1-11; Ettachfini and Andreu, 2004, p. 286, figs 8 b, c.
Description: Test sub-circular, trochospiral, ventral side convex, dorsal area moderately convex; the chambers increasing slightly in size as added; ventral sutures radial, slightly depressed with a small indentation in the septal suture, which points backwards away from the apertural face, umbilical area closed. The aperture extending from near the umbilical region at the base of the apertural face bending below the dorsal wall parallel to the periphery; the bend nearly forms an L-shaped orifice, with a flattened tooth-like outgrowth (Loeblich and Tappan, 1988, p. 83).
Remarks: Nezzazata conica (Smout, 1956) differs from N. simplex by possessing a very convex umbilical side and a smaller umbilicus. N. simplex differs from N. convexa (Smout, 1956) by possessing a flat to slightly convex spiral side rather than a strongly convex planispiral side; and is distinct from N. con-cava (Smout, 1956) because it has a strongly con-cave spiral view. Hauterivian Nezzazata germanicaOmara and Strauch (1965) is poorly described and could be incorrectly assigned to Nezzazata because it does not have the tooth plate (Arnaud-Vanneau and Sliter, 1995).
Occurrence: Nezzazata simplex was recorded in the lower part of the Cenomanian of Gebel Nezzazat, west central Sinai by Omara (1956). Omara and Strauch (1965) extended the stratigraphic range to the early Turonian. Ismail et al. (2009) recorded it in the late Cenomanian. This species has been recorded from the Cenomanian in Italy (Simone et al., 2012), Croatia (Husinec et al., 2000), and Turkey (Sari et al., 2009); Syria (Ghanem et al., 2012). In Mexico, it has been found in the Chiapas Platform (Michaud, 1987) and the Guerrero Platform (Aguilera-Franco, 2003). It has also been found in Honduras by Scott and Finch (1999).
VSLPP in the Llano del Carmen and Guadalcazar sections.
Nezzazata gyra (Smout, 1956) (Figure 9d)
Beggia gr. gyra Smout, 1956, p. 340, pl. 1, figs. 1-9; Nezzazata gyra (Smout) Schroeder and Neumann, 1985, p. 34, pl. 12, fig. 9.
Description: Test plano-convex with three whorls on the spiral side; umbilicus closed, chambers with a narrow internal plate extending back from one septum to the previous septum resulting in a kink or indentation in the septal suture, typical aperture with an apertural tooth.
Remarks: Nezzazata gyra differs from N. simplex by possesing a false umbilicus and from the Aptian- early Albian N. isabellae (Arnaud-Vanneau and Sliter, 1995) by having larger size and a well-developed tooth plate which extends to the previous aperture.
Occurrence: Nezzazata gyra has been reported from the Cenomanian in Italy (Luperto-Sinni, 1966), southern Croatia (Husinec et al., 2009), Syria (Ghanem et al., 2012), Egypt (Ismail et al., 2009), the subsurface of Iraq (Bernaus and Masse, 2006), and Turkey (Farinacci and Yeniay, 1986). VSLPP in the Llano del Carmen, Guadalcazar and Cerritos sections.
Genus Nezzatinella Darmoian, 1976
Type species: Nezzazatinella adhami Darmoian, 1976, p. 492
Nezzazatinella picardi (Henson, 1948a)
Valvulammina picardiHenson, 1948a, p. 613, pl. 15, figs. 1,3, pl. 18, figs.3-6; Nezzazatinella picardi (Henson) Aguilera-Franco et al., 2001, p. 248, fig. 9 i; Sari et al., 2009, p. 60, pl. 2, figs. 12, 13; Rahim-pour-Bonab et al., 2012, p. 28, fig. 8 p.
Description: Test trochospiral, dorsal side flat or slightly convex; with chambers arranged in 1-2 whorls; they are narrow, elongated, with the last chamber flaring backwards, 11 chambers in the last whorl, sutures radial, depressed; the apertural area is wide with a large curved slit, may have secondary apertures in a curved row or scattered over the umbilicus region (Henson, 1948a; Loeblich and Tappan, 1988).
Remarks: Nezzazatinella picardi differs from pre- Coniacian N. adhami Darmoian (1976) by having fewer and differently shaped chambers and a larger aperture with secondary apertures in a curved row or scattered over the apertural face. This species differs from Turonian N. aegyptiaca (Said and Kenawy, 1957) by having more chambers in the last whorl, and more depressed sutures in the umbilical side.
Nezzazatinella picardi is distinguished from Barremian/Aptian N. macoveiiNeagu (1979) by possesing a smaller test, a rounded periphery, chambers less elongated, and radial sutures in the umbilical side in place of a test in later species that measures more than 1 mm, with a lobate and angled margin, and strongly curved sutures in the umbilical view.
Occurrence: N. picardi occurs widely in Upper Cretaceous rocks. This species was described for the first time as Valvulammina picardi from the Santonian beds of Egypt (Henson, 1948a). Forms comparable to it were later described and drawn from the Albian and Cenomanian of Israel, Lebanon, Iran, Slovenia, Croatia, Turkey, and Portugal (Reiss, 1961; Bozorgnia and Banafti, 1964; Dozet and Šribar, 1998; Hamaoui, 1966; Hamaoui and Saint-Marc, 1970; Moro and Jelaska, 1994; Koch et al., 1998; Sari et al., 2009, Berthou and Lauvejat, 1979).
VSLPP in the Llanos del Carmen, Cerritos, and Temazcal sections.
Order Loftusiida Kaminski and Mikhalevich, in Kaminski, 2004
Suborder Loftusiina Kaminski and Mikhalevich, in Kaminski, 2004
Family Cyclamminidae Marie, 1941
Subfamily Hemicyclammininae Banner, 1966
Genus HemicyclamminaMaync, 1953
Type species: Hemicyclammina sigaliMaync, 1953
Hemicyclammina sigaliMaync, 1953
Hemicyclammina sigaliMaync, 1953, p. 148, figs. 1-5; Saint-Marc, 1970, p. 232, pl. 3, figs. 4, 5; Shirazi et al., 2011, p. 783, pl. 2, fig. 11.
Description: Test planispiral, shows a rounded to subacute periphery, with a small proloculus, and a last chamber incompletely subdivided by a discontinuous straight or slightly curved septum; wall agglutinated, outer layer imperforate, and sub-epidermal layer uniformly alveolar; the lower part of the septa coalesces as an imperforate basal layer against the previous whorl, in equatorial section appearing very short and originally termed semi-septa (Maync, 1953; Loeblich and Tappan, 1988).
Remarks: Hemicyclammina sigali differs from Albian H. whitei (Henson, 1948b) because the latter species possesses the septa in contact with the test periphery at right angles to form chambers that are almost quadrangular (in equatorial section). In axial section H. sigali is more compressed.
The Coniacian/Santonian Hemicyclammina chal-masi (Schlumberger, 1899) differs from H. sigali by having a thick and coarsely agglutinated wall and large test with chambers increasing in height, producing a peneropliform test.
Occurrence: Hemicyclammina sigali was described by Maync (1953) from the middle Cenomanian of the Constantine Province in Algeria. This species is recorded in Portugal by Hart et al. (2005), Italy (Simone et al., 2012), Turkey (Bignot and Poisson, 1974), Iran (Shirazi, 2009), Syria (Mouty et al., 2003), Jordan (Weidich and Al-Harithi, 1990), and Brazil (Berthou and Bengtson, 2007). VSLPP in the Llanos del Carmen
Subfamily Choffatellinae Maync, 1958
Genus Pseudocyclammina Yabe and Hanzawa, 1926
Type species: Cyclammina lituus Yokohama, 1890
Pseudocyclammina sp.
Description: Test planispiral with a rounded periphery; composed of three whorled chambers growing rapidly; sutures oblique, wall coarsely agglutinated with a thick subepidermical network with numerous alveoles closely spaced in the agglutinated wall and septa (Loeblich and Tappan, 1988).
Occcurrence: VSLPP in the Llanos del Carmen, Cerritos, El Temazcal sections.
Suborder Biokovinina Kaminski, 2004
Superfamily Biokovinacea Gŭsić, 1977
Family Charentididae Loeblich and Tappan, 1985
Genus Charentia Neumann, 1965
Type species: Charentia cuvillieri Neumann, 1965
Charentia sp.
Description: Test planispiral later partly uncoiled with ten chambers in last whorl; the periphery is rounded; imperforate wall pierced by cylindrical canaliculi resembling keriotheka or pseudoalveolar structure; wall of the early chambers and apertural surface solid, noncanaliculate; the portion of the apertural surface beneath the aperture distinctly thickened to form a triangular chomata-like projection.
Remarks: Charentia sp. differs from Charentia cuvil-lieriNeumann (1965) by having fewer whorls and chambers.
Occurrence: VSLPP in the El Temazcal section.
Suborder Ataxophragmioidea Schwager, 1877
Superfamily Ataxophragmiacea Schwager, 1877
Family Cuneolinidae Saidova, 1981
Subfamily Cuneolininae Saidova, 1981
Genus Cuneolina d’Orbigny, 1839
Type species: Cuneolina pavonia d’Orbigny, 1846
Cuneolina pavonia d’Orbigny, 1846
Cuneolina pavonia d’Orbigny, 1846, p. 253, pl. 21, figs. 50-52; Chiocchini et al., 1984 p. 181, pl. 5, fig. 1; Tasli et al., 2006, p. 529, fig. 6 L; Sari et al., 2009, p. 423, pl. 3-7.
Description: Test compressed conical to flabel-liform, early trochospire followed by very broad, low biserially arranged chambers, commonly compressed parallel to the plane of biseriality; interior subdivided into nearly rectangular chamberlets by radial partitions positioned perpendicular to the outer wall and projecting inward toward the plane of biseriality, and by horizontal partitions paralleling the septa; wall agglutinated, imperforate, with reticulate subepidermal layer (cf. Loeblich and Tappan, 1988, p. 48).
Remarks: The two species reported most often are Cuneolina pavonia d’Orbigny and C. parva Henson. C. pavonia is flabelliform and tends to be slightly larger and have a greater apical angle than C. parva, which is conical and elongate (Arnaud-Vanneau and Sliter, 1995; Arnaud-Vanneau and Premoli-Silva, 1995).
Cuneolina pavonia d’Orbigny (1846) differs from Santonian Cuneolina conica d’Orbigny (1850) by having a broader test. C. pavonia differs from Aptian/Albian Cuneolina sliteriArnaud-Vanneau and Sliter (1995) by having a larger size and flabel-liform test instead of a triangular test with a short biserial stage with 6 or 7 chambers in later species.
Occurrence: VSLPP in the Llanos del Carmen, Cerritos, Temazcal sections.
Cuneolina parva Henson, 1948a
Cuneolina pavonia parvaHenson, 1948a, p. 624-627, pl.14, figs. 1-6, pl. 17, figs. 7-12, pl. 18 figs. 12-14; Cuneolina parva Arnaud-Vanneau and Sliter, 1995, pl. 4, figs. 6-9; Filkorn and Scott, 2011, p. 184, figs. 4, 11-12.
Description: Test compressed, conical. Embry-onic apparatus composed of a large, oval proto-conch covered by a deuteroconch followed by a broad, low biserial stage. Chambers compressed parallel to the plane of biseriality and subdivided by radial beams, and one or two transverse rafters that produce a subepidermal rectangular network.
Remarks: Cuneolina parva was described by Henson (1948a) from the Santonian of Egypt. This species differs from C. pavonia d’Orbigny (1846) by possessing a smaller test with a large megalosphere and smaller biserial stage as well as having a smaller apical angle than C. pavonia has two to three transverse rafters in the chambers compared to one to two in C. parva (Arnaud-Vanneau and Sliter, 1995; Arnaud-Vanneau and Premoli-Silva, 1995). Cuneolina parva is reported from Albian- Cenomanian strata and C. pavonia from uppermost Albian to Santonian (Filkorn and Scott, 2011).
C. parva differs from Barremian/Aptian C. hensoniDalbiez (1958) by having a dense network of radial partitions and more common occurrence of tranverse rafters. C. parva is similar in cross section to the Maastrichtian Iraquian C. cylindricaHenson (1948a). The latter possesses a remarkable triangular test and almost cylindrical early stage.
Occurrence: The species is mostly known from the southern margin of the Tethys in the Middle East, Egypt, Greece, Italy, Albania, Yugoslavia, and North Africa, and from the northern margin in France and Spain (Arnaud-Vanneau and Premoli-Silva, 1995), and in Mexico: as Cuneolina walteri (in Scott and González-León 1991, p. 58; Filkorn and Scott, 2011); in Florida from subsur-face samples (Cushman and Applin, 1947).
VSLPP in the Llanos del Carmen, Cerritos, Guadalcazar, and the El Temazcal sections.
Family Dicyclinidae Loeblich and Tappan, 1964
Genus Dicyclina Munier-Chalmas, 1887
Type species Dicyclina schlumbergeri Munier Chalmas, 1887
Dicyclina schlumbergeri Munier Chalmas, 1887
Dicyclina schlumbergeri Munier Chalmas, p. xxx-xxxi; Schlumberger and Choffat, 1904, p. 362, fig. 1, p. 363, fig. 2; Bonet, 1956, p. 406, pls. 5-7; Sari et al., 2009, p. 61, pl. 3, figs. 8, 9.
Description: Test discoidal, flattened, slightly undulating, initial stage slightly inflated; the megalospheric generation consists solely of the proloculus measuring 700 µm. Later annular chambers added alternating on either side of the test, interior subdivided by numerous thin radial partitions, perpendicular to the outer wall and aligned from chamber to chamber. Wall agglutinated with an imperforate epidermis. The exoskeleton comprises beams and rafters organized in a subepidermic network (Loeblich and Tappan, 1988).
Remarks: Cherchi and Schroeder (1990) stated that Dicyclina was until recently assigned solely to Dicyclina schlumbergeri, a taxon ranging from the late Albian to Maastrichtian (Bilotte, 1984); however, these authors showed that Dicyclina comprises at least two species the mid-Cenomanian D. simplex and the Coniacian/Santonian D. schlumbergeri. These taxa were differentiated based on the different degree of development of the embryonic megalospheric apparatus. We assigned our specimens to D. schlumbergeri, which we consider to be valid because the Mexican specimens have a large, complex embryon (Figure 10g). D. schlumbergeri differs from the Iranian Cenomanian D. sampoi by not having a special layer of the secondary chamberlets in the uppermost part of the supraembryonic zone and the lowermost part of subembryonic zone (Cherchi and Schroeder, 1990).
Occurrence: D. schlumbergeri is widely distributed in the Tethys; it has been reported in Italy (Checconi et al., 2008), Portugal (Berhou and Lauverjat, 1979), Slovenia (Koch et al., 1998), Spain (Ullastre and Masriera, 2006), the Vardar region, Macedonia (Polavder, 2003), Turkey (Farinacci and Yeniay, 1986; Tasli, 2006; Robertson, et al., 2013), Iran (Afghah et al., 2014), and Morocco (Ettachfini et al., 2005; Lézin et al., 2012).
VSLPP in the Llanos del Carmen, Cerritos, Guadalcazar, and El Temazcal sections.
Suborder Orbitolinina Kaminski, 2004
Superfamily Coskinolinoidea Moullade, 1965
Family Coskinolinidae Moullade, 1965
Genus PseudolituonellaMarie, 1954
Type species: Pseudolituonella reicheliMarie, 1954
Pseudolituonella reicheliMarie, 1954
Pseudolituonella reicheliMarie 1954, p. 117-119, text-fig. 2a-b; Luperto-Sinni, 1966, p. 149, pl. 10, fig.1; Chiochini et al., 1984, p. 181, pl. 5, fig. 15; Schroeder and Neumann, 1985, p. 27, pl.8, figs. 1-9; Michaud, 1987, pl. 11, fig. 2; Loeblich and Tappan, 1988, p. 155, pl. 166, figs. 7-11; Aguilera-Franco et al., 2001,p. 248, fig. 9d; Sari et al., 2009, p. 61, pl. 3, fig. 10.
Description: Follows Loeblich and Tappan (1988, p. 155): Test elongate, conical, early portion with a short trochospiral stage, later with broad, low uniserial chambers divided by depressed sutures; interior chamber with tubular pillars projecting upward from the margins of the circular apertures but not completely crossing the chambers; wall microgranular, imperforate. The aperture is cribate with numerous pores in the center of the apertural face.
Remarks: Pseudolituonella reicheli differs from lower Aptian P. conicaLuperto-Sinni and Masse (1993) because the latter species has the initial trochospiral stage in an eccentric position, and a thinner wall.
Barremian Pseudolituonella gavonensis Foury (1968) is different from P. reicheli by having a streptospiral early stage. P. reicheli differs from upper Santonian P. mariaeGendrot (1968) by having a larger test and reduced trochospiral stage, and more developed uniserial stage compared to the latter species, possessing only two or three chambers in the uniserial portion.
Occurrence: Pseudolituonella reicheli is a typical Tethysian species. Its geographical and stratigraphical distribution has already been discussed above.
VSLPP in the Llanos del Carmen and Cerritos sections.
Order Textulariida Delage and Hérouard 1896,
emended Kaminski, 2004
Superfamily Eggerelloidea Cushman, 1937
Family Eggerellidae Cushman, 1937
Subfamily Minouxinae Loeblich and Tappan, 1986
Genre MinouxiaMarie, 1954
Type species: Minouxia guembelitrioidesMarie, 1954, p. 119
Minouxia inflataTronchetti and Zijlstra-Bessais, 2002
Minouxia inflataTronchetti and Zijlstra-Bessais, 2002, p. 541, pl. 1; figs. 1-7; pl. 2, figs. 1-8; pl. 3, figs. 1-4; pl. 4, figs. 3-5
Description: Test conical, elongate, triserial with globular and inflated chambers, the last chamber larger, sutures depressed; wall thin, agglutinated, finely canaliculate, aperture multiple that consists in circular openings in the latest chamber on a trematophore-like plate that covers the umbilical region (Loeblich and Tappan, 1988; Tronchetti and Zijlstra-Bessais, 2002).
Remarks: Minouxia inflata was described from Cenomanian beds of central Tunisia by Tronchetti and Zijlstra-Bessais (2002).
Minouxia differs from Tetraminoxia by having a triserial arrangement instead of quadriserial chambers arranged throughout as in the latter genus. Minouxia inflata is different from Maastrichtian M. guembelitrioidesMarie (1954) by having fewer chambers, the last chamber covering the preceding chambers.
Minouxia inflata differs from Santonian M. conicaGendrot (1963) by having a larger test and globular chambers in place of chambers longer than wide in a conical test.
Minouxia inflata shows globular chambers in axial sections like Santonian M. lobataGendrot (1963).
Occurrence: VSLPP in the Llanos del Carmen, Cerritos, Guadalcazar, and Temazcal sections.
Subclass Tubothalamea Pawlowski, Holzmann, Tyszka, 2013
Order Miliolida Delage and Hérouard, 1896
Superfamily Miliolacea Ehrenberg, 1839
Family Spiroloculinidae Wiesner, 1920
Genus Spiroloculina d’Orbigny, 1826
Type species: Spiroloculina depressa d’Orbigny, 1826
Spiroloculina cenomanaChiocchini, 2008
Spiroloculina cenomanaChiocchini, 2008, p. 187, pl. II, fig. 1.
Description: Test elliptical in outline, with strongly carenate extremities; last chambers changing plane of coiling and showing a sigmoid arrangement, and truncate periphery; sphaeric proloculus followed by a tubular second chamber one whorl in length, the last part with four pairs of chambers added in a single plane; aperture at open end of the final chamber consists of simple or bifid tooth commonly on a short neck. Wall is calcareous, imperforate, porcelanaceous (Chiocchini, 2008).
Remarks: Spiroloculina cenomana differs from S. cretacea due to the former having a sigmoidal chamber arrangement and a truncate periphery.
Occurrence: VSLPP in the Llanos del Carmen, Cerritos, Guadalcazar, and the El Temazcal sections.
Family Hauerenidae Schwager, 1876
Pseudonummoloculina Calvez, 1988
Type species: Pseudonummoloculina aurigerica Calvez, 1988
Pseudonummoloculina heimi (Bonet, 1956)
Nummoloculina heimi Bonet, 1956, p. 402, pl. 3, 4; Conkin and Conkin, 1958, p. 156, pl. 1, figs. 1-10; Michaud, 1987, p. 268, pl. 3, fig. 15; Pseudonum-moloculina heimi (Bonet) Scott and González-León, 1991, p. 61, pl. 5L; Sari et al., 2009, p. 62, pl. 4, figs. 6, 7
Description: Test discoidal, streptospirally involute, with a megalosphere provided with a miliolid bottle-neck (goulot) and at least in adult growth stages, with more than two chambers per whorl; wall calcareous porcelanaceous.
Remarks: The genus Pseudonummoloculina is represented by four species: P. aurigerica Calvez from the early to middle Albian of the Pyrenees, France; P. heimi (Bonet, 1956) from the Albian to Ceno-manian of Mexico; P. robusta (Torre, 1966) from the Senonian of Italy; and P. irregularis (Decrouez and Radoičić, 1977) from the Santonian-Campanian of W Serbia. Pseudonummoloculina heimi differs from early to middle Albian ovoidal P. aurigerica by having a discoidal test and a more reduced early stage but a more developed final stage (Mancinelli and Chiocchini, 2006). Senonian Pseudonummoloculina robusta differs from P. heimi by possessing an inflated test and a very thick wall. The Santonian/ Campanian P. irregularis differs from P. heimi by having more chambers per whorl and a plane of coiling which changes irregularly.
Occurrence: P. heimi is widely distributed in the Albian/Cenomanian of the Gulf Coast and other localities from the Mediterranean area. VSLPP, the La Minita locality.
Superfamily Alveolinacea Ehrenberg, 1839
Family Alveolinidae Ehrenberg, 1839
Ovalveolina Reichel 1936
Type species: Alveolina ovum d’Orbigny, 1850
Ovalveolina maccagnoae De Castro, 1966
Ovalveolina maccagnoaeDe Castro, 1966; Schroeder and Neumann, 1985, p. 106-109, pl. 50, figs. 5-7.
Description: The test of megalospheric form is planispiral involute, spherical proloculus followed by a flexostyle; the chambers short and numerous; in equatorial section the chamberlets can be elongated, elliptical from the first whorls, later appearing rectangular, preseptal passage elon-gated occupies a third part of the chamber; in the subaxial section the chamberlets have a pyriform appearance in the last whorl; wall porcelaneous.
Remarks: The mid-Cretaceous alveolinids are poorly known from the New World but Streptalveolina mexicanaFourcade, Tardy and Vila (1974) was described from the Mexican Cenomanian strata (Aurora Formation). This species is considered endemic to the Caribbean Province, but appears to be related to species in the Mediterranean area such as S. peybernesi (De Castro, 1983). Streptalveolina mexicana has been also recorded in some localities of Gulf Coast in the Albian-Cenomanian (Scott, 2002; Mancini and Scott, 2006). Omaña et al. (2012) reported Ovalveolina maccagnoaeDe Castro, 1966 from the Cenomanian deposits of the Manantlan Range in Colima State, Mexico.
Further alveolinids Sellialveolina sp. and S. cf. drorimensis were reported from the Cenomanian in northern Peru by Jaillard and Arnaud-Vanneau (1993, p. 603).
Occurrence: VSLPP in the La Minita outcrop.
Family Peneroplidae Schultze, 1854
Peneroplis de Montfort, 1808
Type species: Peneroplis planatus Fichtel and Moll, 1798, p. 91
Peneroplis parvus De Castro, 1965
Peneroplis parvusDe Castro, 1965, p. 345-348, pl. 1, 14-15; Schroeder and Neumann, 1985, p. 86, pl. 39, figs. 1-17; Tasli et al., 2006, p.529, pl. 6, fig. O.
Description: Test with a rounded proloculus, early stage planispirally coiled, later with or
without uncoiling stage; the chambers quickly increasing in width, resulting in a flaring peneropliform test. Sutures slightly depressed. Wall is calcareous imperforate porcelaneous; the surface with numerous grooves alternating with fine ribs perpendicular to the sutures.
Remarks: Peneroplis parvus is like the Recent P. planatus but differs by having a smaller size and more numerous chambers. It differs from P. turon-icus (Said and Kenawy, 1957) because the latter species lacks radial striae on the outside surface. By this character P. turonicus could be assigned to Lae-vipeneroplis, according to the foraminiferal generic classification of Loeblich and Tappan (1988).
Peneroplis was placed in the family Soritidae by Schroeder and Neumann (1985); later Loeblich and Tappan (1988) settled it in the family Pen-eroplidae because this form has chambers with a simple interior, while the family Soritidae has chambers subdivided by interseptal pillars or septula.
Occurrence: Peneroplis parvus was described for the first time by De Castro (1966) from the mid-Cenomanian from the Campania region (Italy); this species has been recorded from Spain (Calonge and Caus, 1996), Croatia (Velić and Vlahović, 1994), Turkey (Tasli, 2006), and Syria (Ghanem and Kuss, 2013).
VSLPP in the Llanos del Carmen, Cerritos, Guadalcazar, and Temazcal sections.
Subclass Globothalamea Pawlowski, Holzmann and Tyszka 2013
Order Rotaliina Delage and Hérouard, 1896
Superfamily Planorbulinacea Schwager, 1877
Family Cymbaloporidae Cushman, 1927
Archaecyclus Silvestri. 1908
Type species: Planorbulina cenomaniana Seguenza, 1882
Archaecyclus Silvestri, 1908, p. 134
Archaecyclus sp.
Test large, convex planispirally coiled, five chambers in the first whorls, later forming an annular arrangement; the sutures are oblique, the wall calcareous, perforated, having a dark thin layer in middle position. The aperture is interiomarginal in the early stage; after with stolon-like openings at the lateral margins of each chamber (Loeblich and Tappan, 1988).
Occurrence: Archaecyclus sp. has been recorded by Jaillard and Arnaud-Vanneau (1993) from Peru. VSLPP has been only observed in the Llanos del Carmen section.
5. Conclusions
The deposits of the upper part of the El Abra Formation preserved a rich and diverse association mostly composed of larger benthic foraminifera, which are described and illustrated for the studied succession. The species of benthic foraminifera have a significant age-diagnostic value in the El Abra shallow-water deposits, dating the succession studied here as mid-late Cenomanian.
The foraminiferal and algal assemblages are age-equivalent to the Mediterranean associations. Two zones are proposed for the sections from the El Abra Formation: the Pseudolituonella reicheli Assemblage Zone and the Nezzazatinella picardi Interval Zone.
The Pseudolituonella reicheli Assemblage Zone contains a rich and varied benthic foraminiferal association which was reduced to a few foraminifers that survived. The Nezzazatinella picardi Interval Zone later also disappeared when the hemipelagic Soyatal deposit took place (Whiteinella archaeocretacea Partial Range Zone) due to the large scale flooding and climate perturbations.
The integration of the data provided by the benthic foraminiferal association and the lithology enable a more reliable paleoenvironmental interpretation.
We consider that there was a first episode (mid- late Cenomanian) in the Taninul facies of the El Abra Formation consisting of a warm shallow-water carbonate platform with open circulation and medium-high hydrodynamic energy. The maximum diversity of benthic foraminiferal assem-blages coincides with this environment. Later, a decrease of benthic foraminiferal species diversity is recorded, with a few foraminiferal species surviving (Nezzazatinella picardi, miliolids, rotalids, and textularids) in association with algae. This change could be related to increasing nutrient availability in mesotrophic conditions.
The drowning event of the platform with a hemi-pelagic deposit (Soyatal Formation) occurred in the latest Cenomanian. It is marked by the bloom of pithonellids and r and r-k strategist planktic foraminifera and benthics such as gavellinellids, which are present in a deeper and eutrophic environment.
An environment with restricted circulation from the El Abra facies is distinguished by a shallow-water association of reduced diversity but commonly with numerous individuals of typical miliolid for-aminifera. Another intermediate semi-restricted environment with a greater variety of foraminifera is also defined.
The foraminiferal and algae community have a marked Tethysian affinity.

