











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink1. Introducción
El extinto lobo gigante,Canis dirusLeidy, 1858, se distingue por ser la especie de mayor talla y con mayor grado de hipercarnivoría dentro del género (Merriam, 1911;Nowak, 1979; Berta, 1988; Tedford et al., 2009). Esto se ve reflejado en una masa corporal estimada entre los 60 y 68 kg (Anyonge y Roman, 2006) y en un aumento del contacto de cizallamiento entre los dientes superiores e inferiores (Anyonge y Baker, 2006; Wang y Tedford, 2010).
Su distribución geográfica se extiende desde Alberta, Canadá, hasta Tarija, Bolivia (Nowak, 1979; Berta, 1988; Dundas, 1999). Los registros de esta especie están concentrados principalmente en Estados Unidos con 123 localidades, luego en México con 15 localidades, Canadá y Venezuela con dos localidades cada uno y finalmente Perú y Bolivia con una localidad cada país (Dundas, 1999; Prevosti y Rincón, 2007;Carbot-Chanona et al., 2016; el presente trabajo). Cronológicamente esta especie abarcó desde el Irvingtoniano tardío hasta el Rancholabreano (NALMA) en Norteamérica (Tedford et al., 2009) y el Lujanense (SALMA) en Suramérica (Berta, 1988). En ambos casos, la biocronología va desde el Pleistoceno tardío al Holoceno (Dundas, 1999; Berta, 1988), aunque Wang y Tedford (2010) la restringen solamente al Pleistoceno tardío.
Se ha propuesto que, debido a la amplia distribución geográfica de este lobo durante el Pleistoceno, existieron varias poblaciones morfológicamente distintas en Norteamérica.Frick (1930) fue el primero que nombró dos subespecies deAenocyon(=Canis)dirus:A. dirus alaskensispara Alaska yA. dirus nebrascensispara Nebraska, sin embargo, éstas no son válidas debido a que no contaron con una descripción formal (Nowak, 1979). Más adelante, Kurtén (1984) evidenció ligeras diferencias a nivel de la talla de los huesos de las extremidades y el largo del segundo premolar superior (P2), concluyendo que los individuos colectados desde noreste de Estados Unidos hasta Canadá eran corporalmente más grandes y tenían un P2 más corto, contrariamente a los individuos colectados en México y la costa occidental de Estados Unidos. Bajo este argumento, este autor dividió a esta especie enC. dirus dirusLeidy, 1858 yC. dirus guildayiKurtén, 1984 (Figura 1).

Kurtén (1984) también señaló biocrones para ambas subespecies. En la diagnosis, describe queC. dirus dirusvivió durante el Rancholabreano (240000-11000 años AP) mientras queC. dirus guildayidurante el interglaciar Sangamoniano (125000-75000 años AP) y la glaciación Wisconsiniana (85000-11000 años AP). Sin embargo, existe una confusión al respecto de los biocrones, porque en otra sección del mismo artículo se menciona queC. dirus dirusyC. dirus guildayihabitaron durante el mismo período. Posteriormente, Wang (1990) publicó un estudio donde compara miembros deC. dirus dirusde la glaciación Wisconsiniana con respecto a los deC. dirus guildayidel Rancholabreano, destacando que hay diferencias en cuanto a la talla de los dientes entre los dos grupos de cánido, apoyando así la propuesta de Kurtén (1984). Casi dos décadas después,Tedford et al. (2009) realizaron la última revisión de la especie en Norteamérica, pero no hicieron referencia a las subespecies anteriormente mencionadas. En cambio, sí realizaron una distinción entre las poblaciones que vivieron durante el Irvingtoniano tardío con respecto a las del Rancholabreano en Estados Unidos, sugiriendo que existen ligeras diferencias a nivel de la talla de los dientes, sin atribuirlas a ninguna de las subespecies descritas previamente.
En el caso de México, a la fecha no se ha realizado ninguna revisión de los restos deC. dirusdescubiertos, y se sigue la asignación taxonómica propuesta por Kurtén (1984) (i.e.,C.dirus guildayi). México cuenta con 15 localidades donde se han registrado restos fósiles deC. dirus(Figura 2, Tabla 1). Pese a esto, Kurtén (1984) se basó, para su propuesta biogeográfica y taxonómica, solamente en los individuos de la Cueva de San Josecito, Nuevo León, y un polémico ejemplar de El Tajo de Tequixquiac, Estado de México (el cual será discutido más adelante).
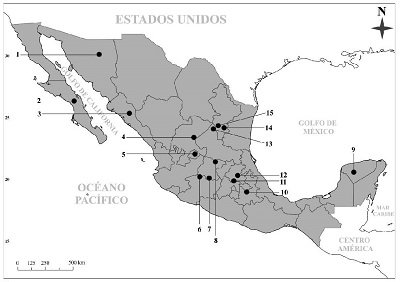
Figura 2 Localidades referidas paraCanis dirusen México. 1-Térapa, Sonora; 2-Comondú, Baja California Sur; 3-Potrecito, Sinaloa; 4-Chupaderos, Zacatecas; 5-Cedazo, Aguascalientes; 6-La Cinta-Portalitos, Michoacán; 7-La Piedad-Santa Ana, Michoacán; 8-Cuitzeo, Michoacán; 9-Gruta de Loltún, Yucatán; 10-Valsequillo, Puebla; 11-El Tajo de Tequixquiac, Estado de México; 12-Pachuca-Tulancingo/San Agustín Tlaxiaca, Hidalgo; 13-El Cedral, San Luis Potosí; 14-Cueva de San Josecito, Nuevo León; 15-Minas, Nuevo León.
Cabe mencionar que el material mexicano en el que se basó Kurtén (1984) es el más completo y mejor preservado hasta la fecha. El resto del material asignado aC. dirusen México consiste en elementos dentales y postcraneales aislados, fragmentados y muy desgastados.
En este trabajo se presenta una actualización del estado del conocimiento de esta especie para México, y se evalúa si es posible apoyar la asignación a nivel de subespecie propuesta por Kurtén (1984). Para esto nos basamos en un mayor número de especímenes mexicanos que los utilizados por este autor, donde además incluimos ejemplares ya publicados y otros depositados en colecciones (Instituto Nacional de Antropología e Historia, INAH, en México y la del Natural History Museum, Los Angeles County, LACM, en Los Ángeles, California, Estados Unidos).
Tabla 1 Registros fósiles de Canisdirus en México.
| Localidad | Edad | Referencia |
|---|---|---|
| Térapa, Sonora | Pleistoceno tardío (Rancholabreano) | Hodnett et al . (2009) |
| Comondú, Baja California Sur | Pleistoceno tardío | Berta, 1988; Dundas (1999) |
| Potrecito, Sinaloa | Pleistoceno tardío (Rancholabreano) | Kurtén (1984) |
| Chupaderos, Zacatecas | Pleistoceno tardío | Barrón-Ortiz et al . (2009); Ferrusquía- Villafranca et al . (2017) |
| Cedazo, Aguascalientes | Pleistoceno (Irvingtoniano- Rancholabreano) | Mooser y Dalquest (1975); Montellano-Ballesterios (1990) |
| La Piedad-Santa Ana, Michoacán | Cuaternario | Cervantes (2015) |
| La Cinta-Portalitos, Michoacán | Pleistoceno tardío | Cervantes (2015) |
| Cuitzeo, Michoacán | Pleistoceno | Este trabajo. |
| Gruta de Loltún, Yucatán | Pleistoceno tardío- Holoceno (G. Wisconsin) | Álvarez y Polaco (1982); Dundas (1999) |
| Valsequillo, Puebla | Pleistoceno tardío | Thenius (1970) Carbot-Chanona (2016) |
| Tequixquiac/El Tajo, Estado de México | Pleistoceno tardío | Freudenberg (1910); Maldonado- Koerdell (1953); Kurtén (1984) |
| Pachuca-Tulancingo/San Agustín Tlaxiaca, Hidalgo | Pleistoceno | Bravo-Cuevas et al . (2009) |
| Rancho “La Amapola” (El Cedral), San Luis Potosí | Pleistoceno tardío- Holoceno | Álvarez et al . (2012) |
| Cueva San Josecito, Nuevo León. | Pleistoceno tardío | Arroyo-Cabrales et al . (1995) |
| Minas, Nuevo León | Pleistoceno tardío (Rancholabreano) | Ferrusquía-Villafranca et al . (2017) |
2. Materiales y métodos
2.1.1. Generales
AP: Antes del presente.
Ca: California.
Ce: El Cedral.
CP: Componente principal.
Fl: Florida.
IrvT: Irvingtoniano tardío.
Ma: Millones de años.
MC: Maricopa.
MK: McKittrick.
MX: México.
NALMA: North American Land Mammal Ages (Edades Mamíferos de Norteamérica).
Ne: Nebraska.
Ran: Rancholabreano.
RLB y RB: Rancho La Brea.
SALMA: South American Land Mammal Ages (Edades Mamíferos de Suramérica).
SJ: Cueva de San Josecito.
So: Sonora.
USA: Estados Unidos.
Wis: Glaciación Wisconsiniana.
2.2. Análisis Morfométricos
Se tomaron datos morfométricos de los ejemplares depositados en las siguientes colecciones: Colección Nacional de Paleontología, Instituto de Geología (IGM), Ciudad de México, México; Florida Museum of Natural History (UF), Gainsville, Florida, Estados Unidos; Instituto Nacional de Antropología e Historia, Departamento de Prehistoria (INAH, DP), Ciudad de México, México; La Brea Tar Pit & Museum, Los Ángeles, California, Estados Unidos; Natural History Museum, Los Angeles County (LACM), Los Ángeles, California, Estados Unidos; Royal Ontario Museum (ROM), Toronto, Canadá (ver Apéndice donde se detallan los ejemplares revisados).
Las variables dentales consideradas fueron el largo (mesio-distal) y ancho (buco-lingual) del P3, P4, M1 (largo tomado en la cara bucal), M2, p4, m1 (ancho tomado a la altura del trigónido) y m2. Las medidas fueron tomadas con un vernier analógico (± 0.1 mm, máx. 150 mm) y fueron representadas en milímetros (mm). Adicionalmente se consideraron las medidas reportadas por Tedford et al. (2009) para especímenes de Estados Unidos; por Wang (1990) para un espécimen de Kansas; porMaldonado-Koerdell (1953) para el espécimen de Tequixquiac y porHodnett et al. (2009) para el espécimen de Térapa, Sonora.
La base de este estudio se fundamentó en evaluar si existían diferencias morfométricas entre grupos definidos deCanis diruspara Norteamérica: (1)C. dirus diruscon respectoC. dirus guildayi, (2)C. dirusdel Irvingtoniano con respecto a los del Rancholabreano y finalmente (3) miembros recolectados en México con respecto a los miembros de Estados Unidos.
Por otro lado, se realizaron análisis estadísticos para evaluar la propuesta taxonómica de Kurtén (1984). Para esto se aplicó una prueba de T (p= 0.05) mediante el uso del software libre PAST (v. 3.17) (Hammer et al., 2001). Con el fin de analizar la existencia de agrupaciones basadas en las variables morfométricas descritas anteriormente, se aplicó un análisis multivariado de ordenación y clasificación llamado Análisis de Componentes Principales (ACP) con el mismo software PAST. Los valores analizados no fueron estandarizados, esto con el objetivo de evidenciar con mayor exactitud la existencia de diferencias en las magnitudes de las estructuras dentales. Análisis y gráficas adicionales fueron realizadas mediante el uso del software Excel, Microsoft Office 365.
2.3. Estimación de masa corporal
La masa corporal fue estimada mediante la regresión propuesta por Van Valkenburgh (1990) basada en el largo del m1 (log10Y = ENT#091;K * log10XENT#093; + log10b), donde “Y” es la masa corporal (en kg), “X” es el largo mesiodistal del m1 (en mm), “K (= 1,82)” es un coeficiente alométrico y “b (= -1,22)” es la intercepción con el eje “y” (r= 0.87).
3. Resultados
3.1. Taxonomía
Kurtén (1984) definió aCanis dirus diruspor tener el P2 más corto queC. dirus guildayi. Para este argumento, el autor realizó una prueba estadística bajo la cual reportó que existían diferencias significativas entre ambos grupos. Retomamos los datos que este autor utilizó, y bajo una prueba de T encontramos que no existen diferencias significativas entre los individuos de Rancho La Brea y Cueva de San Josecito (C. dirus guildayi) con respecto a los individuos reportados del noreste de Estados Unidos para el interglaciar Sangamoniano (C. dirus dirus). Pero sí se encontraron diferencias significativas entre los miembros deC. dirus guildayicon respecto a losC. dirus dirusde la glaciación Wisconsiniana (Tabla 2).
Tabla 2 Valores de p obtenidos de una prueba de T (p = 0.05) utilizando los datos reportados por Kurtén (1984) para el largo del P2 de dos subespecies de Canis dirus.
| Canis dirus dirus Sangamoniano | Canis dirus dirus Wisconsiniano | |
|---|---|---|
| Canis dirus guildayi | T = - 0.511; p = 0.616 | T = 2.369; p = 0.029 |
| Rancho La Brea | ||
| Canis dirus guildayi | T = 0.811; p = 0.436 | T = 3.658; p = 0.003 |
| Cueva San Josecito |
Para evaluar si existían diferencias bajo la perspectiva cronológica de Tedford et al. (2009), se tomaron los valores del P2 reportados por estos autores para miembros del Irvingtoniano tardío de Nebraska y California y se compararon con individuos del Rancholabreano de California. La prueba T (p= 0.5705) indica que tampoco hay diferencias significativas bajo este criterio cronológico (Tabla 3).
Tabla 3 Prueba de T para el largo del P2 de individuos de Canis dirus del Irvingtoniano tardío con respecto a individuos del Rancholabreano.
| N° Catálogo | Localidad/Edad | Largo P2 (mm) |
|---|---|---|
| F:AM 25511 | Nebraska/Irvingtoniano tardío | 14.8 |
| UCMP 156048 | California/Irvingtoniano tardío | 14.8 |
| AMNH 14339 | California (Rancho La Brea)/Rancholabreano | 15.7 |
| AMNH 15867 | California (Rancho La Brea)/Rancholabreano | 14.7 |
Otro carácter que describió Kurtén (1984) para diferenciar entre las subespecies deC. dirus, es un alargamiento mayor de las extremidades enC. dirus diruscon respecto a su contraparteC. dirus guildayi. Bajo esta perspectiva los miembros mexicanos y californianos deberían ser ligeramente más pequeños que sus contrapartes del noreste de Norteamérica. Sin embargo, como se mencionó anteriormente, el material mexicano es escaso para realizar esta comparación. A la falta de esta información se evaluó el tamaño utilizando una estimación de masa corporal (Figura 3) mediante la ecuación de Van Valkenburgh (1990) (véase apartado 2 del presente trabajo, Materiales y Métodos).
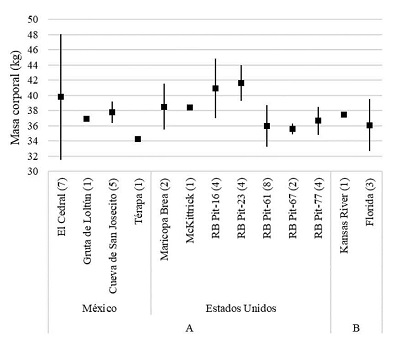
Figura 3 Masa corporal en kg estimada paraCanis dirusen localidades de Norteamérica. Se representa el promedio con un cuadrado (■) acompañado de barras que indican la Desviación Estándar. Los valores entre paréntesis significan el número de individuos. A-Canis dirus guildayi; B-C. dirusdirus.
La masa corporal paraC. dirustiene un rango que va desde 31.52 hasta 48.06 kg. Curiosamente este rango se evidenció para la población de El Cedral, Rancho “La Amapola”, San Luis Potosí. En las localidades restantes, tanto de México como de Estados Unidos, los individuos tuvieron rangos más restringidos, y se incluyeron dentro del espectro de los de El Cedral.
Descartando esta localidad mexicana, no se evidencia una distinción entre la masa corporal de miembros deC. dirus guildayicon respecto a los deC. dirus dirus. Sólo se evidencia un máximo entre las masas corporales de los individuos de Rancho La Brea Pit 16 y Pit 23, donde las medias son 40.89 y 41.59 kg respectivamente.
3.2. Otras consideraciones morfométricas
Tanto Wang (1990) comoHodnett et al. (2009) se basaron en variables de los dientes inferiores para asignar algunos especímenes deCanis dirusa alguna de las subespecies propuestas por Kurtén (1984). De acuerdo con Hodnett et al. (2009), el ejemplar TERA-154 (Sonora) mostró un valor dental similar (largo del p4) acorde aC. dirus gulidayipropuesto por Kurtén (1984). Replicando la prueba de T para este carácter encontramos que no hay diferencias significativas entre los miembros descritos paraC. dirus guildayicon respecto aC. dirus dirusen cuanto al largo del p4 (Tabla 4). Sólo existe una ligera diferencia entre los miembros rancholabreanos de Rancho La Brea con respecto a los miembros de la glaciación Wisconsiniana del noreste de Estados Unidos.
Tabla 4 Valores de p obtenidos de una prueba de T (p= 0,05) utilizando los datos reportados por Kurtén (1984) para el largo del p4 de dos subespecies deCanis dirus.
| Canis dirus dirus Sangamoniano | Canis dirus dirus Wisconsiniano | |
|---|---|---|
| Canis dirus guildayi | T = - 0.749; p = 0.459 | T = 2.530; p = 0.015 |
| Rancho La Brea | ||
| Canis dirus guildayi | T = -1.684; p = 0.101 | T = 1.571; p = 0.123 |
| Cueva San Josecito |
Los valores reportados por Kurtén (1984) para el largo del p4 tienen un rango que va de 15.0 a 21.8 mm paraC. dirus guildayi, mientras que paraC. dirus diruslos valores van de 13.5 a 15.5 mm. Sin embargo, hay individuos identificados comoC. dirus diruscon valores que pueden llegar a 20.5 mm (UNSM-21432 de Nebraska), lo que amplía el rango de medidas (tamaño) para este grupo de cánidos, haciendo que el valor de TERA-154 (19.5 mm) quede dentro del rango de ambas subespecies, incluso dentro de los grupos cronológicos sugeridos por Tedford et al. (2009) (Figura 4). Esto no sólo se evidencia en el ejemplar de Sonora, sino que también ocurre en individuos de El Cedral y la Cueva de San Josecito en México.
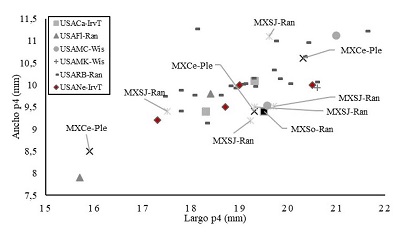
Figura 4 Distribución de la talla del p4 en distintas asociaciones deCanis dirusen Norteamérica. Los símbolos en gris-negro equivalen a los miembros deC. dirus guildayi, mientras que los símbolos en rombo de color rojo equivalen aC. dirus dirussegún la propuesta deKurtén (1984).
También se aplicó un análisis de componentes principales para considerar la relación entre distintas variables dentales, tanto de dientes superiores como de inferiores (véase el apartado 2, Materiales y Métodos, para la descripción de las variables consideradas). Pese a que el número de individuos asignados aC. dirus diruses considerablemente menor con respecto a los deC. dirus guildayi, no se encontraron claras diferencias entre los miembros de ambas subespecies. Esto ocurre tanto en dientes inferiores (Figura 5A) como dientes superiores (Figura 5B). En ambos casos se observaron solapamientos entre los taxones que Kurtén (1984) diferenció (véanse las áreas demarcadas en Figura 5). Tampoco se observa una distinción entre los individuos del Irvingtoniano con respecto a los del Rancholabreano o glaciación Wisconsiniana.
Finalmente, en ambos ACP no se encontraron agrupamientos diferenciales entre los individuos de México y los de Estados Unidos. Lo que indica que no hay diferencias evidentes entre estos grupos bajo los caracteres dentales utilizados.

Figura 5 Análisis de componentes principales (ACP) para variables dentales deCanis dirusde Norteamérica (variables definidas en Materiales y Métodos). Los puntos representan ejemplares deC. dirus guildayidel Rancholabreano, los cuadrados equivalen a miembros del Irvingtoniano tardío, los rombos equivalen a ejemplares deC. dirus dirus. Se muestra el área que ocupa cada subespecie; notar que en algunas regiones estas áreas se solapan. A-Dientes inferiores (CP 1 = 76,45 % de Varianza; CP 2 = 8,84 % de Varianza). B-Dientes superiores (CP 1 = 48,87 % de Varianza; CP 2 = 22,92 % de Varianza).
4. Discusión
4.1. Taxonomía
Si bien la propuesta taxonómica de Kurtén (1984) es actualmente aceptada, pocos autores la han seguido. Wang (1990) mencionó que, aunque las diferencias entreCanis dirus dirusyC. dirus guildayison muy pequeñas, esta subdivisión se justifica bajo la consideración de que el actual lobo gris americano (C. lupus) tiene múltiples subespecies reconocidas.
Posteriormente, Anyonge y Roman (2006) realizaron estimaciones de masa corporal donde concluyeron queC. dirus dirusera más grande queC. dirus guildayi,con una masa promedio entre 68 y 60 kg respectivamente. Nuestras estimaciones, utilizando la ecuación de Van Valkenburgh (1990), no se aproximan a los valores de Anyonge y Roman (2006), esto es probablemente debido a diferencias metodológicas que consideran distintas estructuras óseas, lo que modifica las ecuaciones de regresión para esta estimación. Nuestros resultados obtenidos indican que las masas corporales promedio fueron muy similares en ambas subespecies:C. dirus dirus(36.97 kg) yC. dirus guildayi(37.64 kg).
Algunos estudios han medido la evolución del tamaño corporal a lo largo del tiempo en el mismo yacimiento como en Rancho La Brea, California. Los análisis realizados para las poblaciones deC. dirusapuntan que durante las épocas más frías (Pit 16; 68000 a 11503 años AP) las poblaciones de este lobo eran más pequeñas con respecto a las épocas más calientes (Pit 61, 67 y 77; época interglaciar, 4450 a 40570 años AP) (O’Keefe et al., 2009,2014;O’Keefeet al., 2014;Brannick et al., 2015), lo cual es congruente con nuestros resultados para Rancho La Brea (véase Figura 3).
De manera peculiar, la masa corporal estimada para los individuos de los pozos o Pit 61, 67 y 77 es parecida a la de los individuos de la Cueva de San Josecito, la cual está fechada entre 27000 y 45000 años AP y correspondería a una época menos fría (Arroyo-Cabrales et al., 1995).
Por otro lado,Hodnett et al. (2009) identificaron el ejemplar de Térapa, Sonora (TERA 154), comoC. dirus guildayi, basándose en la talla de los dientes de la mandíbula. Sin embargo, nuestros resultados indican que este ejemplar no presenta información suficiente que lo permita asignar aC. dirus dirusoC. dirus guildayi, más allá de su ubicación geográfica. Adicionalmente, los rangos morfométricos reportados por Kurtén (1984) difieren cuando se consideran otros individuos de la misma especie (ej. UNSM-21432 de Nebraska), de lo que se desprende que los rangos morfométricos no son parámetros diagnósticos para separar las subespecies.
Aunque Tedford et al. (2009) no consideraron las diferencias biogeográficas y morfométricas propuestas por Kurtén (1984), reportaron que las tallas difieren entre los individuos del Irvingtoniano y del Rancholabreano de Norteamérica. Pese a que no todos los ejemplares fósiles estudiados por nosotros provienen de localidades con un fechamiento exacto, no encontramos diferencias en el tamaño o talla de los dientes de individuos de estos períodos.
4.2. Canis dirusen México
Anteriormente se mencionó que México cuenta con 15 registros fósiles deC. dirus. Algunos de estos registros provienen de listas faunísticas publicadas en las que no se incluyen detalles del material, tales como descripciones, figuras, números de catálogo o medidas. Este es el caso del registro de Comondú en Baja California Sur (Berta, 1988). Una situación similar se repite con los reportes de Potrecito (Sinaloa), Minas (Nuevo León) y Chupaderos (Zacatecas) (Barrón-Ortiz et al., 2009; Ferrusquía-Villafranca et al., 2017).
La situación es contraria en localidades como Térapa (Sonora), El Cedral (San Luis Potosí), Cedazo (Aguascalientes), La Piedad-Santa Ana y La Cinta-Portalitos (Michoacán), San Agustín Tlaxiaca (Hidalgo), El Tajo/Tequixquiac (Estado de México), Valsequillo (Puebla), Gruta de Loltún (Yucatán) y la Cueva de San Josecito (Nuevo León).
Los registros de C. dirus en México están principalmente concentrados al centro del país. Sin embargo, algunos reportes como el de Hodnett et al. (2009), donde descubrieron una mandíbula y un húmero de C. dirus, se localizan en región norte del país (Térapa, Sonora). Por otro lado, la Cueva de San Josecito es la localidad con mayor abundancia de elementos esqueléticos asignados a C. dirus, más de 90 elementos fósiles (según la base de datos de la Colección Paleontológica de Vertebrados del Natural History Museum Los Angeles County, https://collections.nhm.org/vertebrate-paleontology), y con el mejor estado de preservación (Figura 6A).
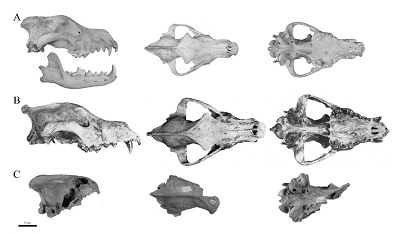
Figura 6 Canisdirus. A-LACM 9795 (cráneo) y LACM 9229 (mandíbula), Cueva de San Josecito, Nuevo León; B-Sin número de catálogo, tomado y modificado de Maldonado-Koerdell (1953), Tequixquiac, Estado de México; C-IGM 5239, Tequixquiac, Estado de México.
Cercana a esta cueva está la localidad de El Cedral, en “El Rancho La Amapola”, San Luis Potosí. Aunque Álvarez et al. (2012) mencionaron que el tamaño de los ejemplares de esta localidad es similar a los de Rancho La Brea en California, los siete individuos examinados en el presente estudio indican una variabilidad mayor en la masa corporal que la descrita para los ejemplares de Estados Unidos. Hacia el centro de México se ubica la localidad de Cedazo, en el Estado de Aguascalientes. Aunque las referencias son escasas,Mooser y Dalquest (1975) reportaron restos mandibulares y postcraneales de un cánido que tiene una talla similar aC. dirusde Rancho La Brea.
En la colección del INAH hay un ejemplar DP 5100 que corresponde a un primer molar superior (M1) proveniente de la región de Cuitzeo, Michoacán, el cual no está citado en la literatura. El ejemplar presenta un carácter diagnóstico deC. dirus(i.e., hipocono reducido), por lo cual se incluyó en este trabajo. En la frontera de Michoacán y Guanajuato, Cervantes (2015) reportó dos localidades (La Piedad-Santa Ana y La Cinta-Portalitos) donde fueron descubiertos elementos muy deteriorados de cánidos, los cuales presentan una talla similar a laC. dirus. Para el Estado de Hidalgo, Bravo-Cuevas et al. (2009) reportaron un radio al cual asignaron aC. dirus, aunque no lo describieron.
Después de la Cueva de San Josecito, Tequixquiac, en el Estado de México, representa la localidad con los ejemplares fósiles más completos asignados aC. dirusen México. Un espécimen interesante es un cráneo completo descrito por Maldonado-Koerdell (1953) (Figura 6B). Sin embargo, se desconoce la ubicación actual de este fósil, ya que este autor no reporta número de catálogo. Dundas (1999) marca como distintas a Tequixquiac y a El Tajo. Para El Tajo hace referencia aFurlong (1925), pero este autor nunca mencionó la presencia de restos deC. dirus, en cambio mencionó la presencia del géneroCanispara los estratos pleistocénicos de la localidad de Tequixquiac. Este mismo autor, cuando hizo referencia a este taxa, se refirió a “possibly n. sp. cf.ocrupus”. Esta nueva especie nunca fue formalmente propuesta, por lo cual es inválida. Por otro lado, también nombró a un cánido (sin asignación taxonómica específica) para las cercanías de la cantera El Tajo. La confusión ocurre cuando Kurtén (1984) hace referencia a un cráneo y mandíbulas (UCMP 26650) provenientes de esta región, pero lamentablemente no ha sido posible confirmar si el cráneo del que hace referencia este autor es el mismo reportado por Maldonado-Koerdell (1953). Otro fósil proveniente de Tequixquiac es un basicráneo (IGM 5239) asignado aC. dirus(Figura 6C). Históricamente, este fósil representa el primer reporte deC. diruspara México, aunque su autor,Freudenberg (1910), lo asignó originalmente aC. indianensis, y un año despuésMerriam (1911)lo nombróC. dirus, e indicó que no existen diferencias entre este ejemplar y el material de Rancho La Brea, California.
Las localidades más al sur de México y, por ende, de Norteamérica, son las de Valsequillo, en Puebla y Gruta de Loltún, en Yucatán. Para Valsequillo fueThenius (1970) quien realizó uno de los primeros reportes deC. dirusen la localidad y recientementeCarbot-Chanona et al. (2016) reportaron nuevos elementos mandibulares. Por su lado, Álvarez y Polaco (1982) fueron los primeros en mencionar la presencia deC. dirusen la Gruta de Loltún, reportando diversos elementos craneales y postcraneales. Estos autores basaron su identificación en la talla de los elementos: su clasificación queda confirmada con los caracteres diagnósticos presentes en un primer molar inferior (m1) y un cuarto premolar superior (P4) asociados (DP 269) (i.e., presencia de cresta entre el metacónido y el hipocónido del m1 y el protocono del P4 está muy reducido).
El registro fósil de México es aún mayor del que se enumera en este trabajo. En la colección paleontológica del INAH, están almacenados dos fragmentos mandibulares (DP 2907 y 1924), los cuales sólo tienen como referencia “La Paz”. Debido a que no hay mayor información de la localidad, no es posible distinguir si proviene del Estado de México o de Baja California Sur.
Finalmente, en múltiples ocasiones se ha mencionado la presencia deC. dirusen Chapala, Jalisco (Nowak, 1979; Berta, 1988; Dundas, 1999, Ferrusquía-Villafranca et al., 2017), sin embargo,Rufolo (1998) identificó un M2 izquierdo, anteriormente asignado aC. dirus, como posiblemente perteneciente aC. latrans.
Luego de la aceptación de este trabajo se descubrió dentro de la Colección Geográfica de Referencia del Instituto de Geología de la UNAM un basi cráneo fósil (IGM 8871) correspondiente aCanis dirus, proveniente de la caverna La Cieneguilla en el Municipio Indé en el Estado de Durango, el cual fue donado por el Sr. Abel Chávez Ramírez en 1974.
5. Conclusiones
México es el segundo país con registros fósiles deCanis dirus, esto representa casi el 10 % de la distribución total de este lobo en América. Aunque anteriormente todos estos registros habían sido clasificados bajo la subespecieC. dirus guildayi, nuestros resultados no apoyan la separación de esta especie en subespecies, como lo propuso Kurtén (1984). Infortunadamente, el material deC. dirusen América no es muy abundante, y no se descarta la idea de Wang (1990) de que dentro de la amplia distribución geográfica de esta especie pudieran haber existido variaciones morfológicas o morfométricas. Sin embargo, en nuestros análisis no encontramos diferencias significativas que permitan algún tipo de fragmentación o subdivisión taxonómica de esta especie en Norteamérica. Es probable que exista mayor claridad en el tema si se realizan nuevos estudios, considerando mayor cantidad de ejemplares completos provenientes del noreste de Estados Unidos.
Después de haber realizado esta revisión del material mexicano hay varios puntos que sobresalen y que plantean nuevas interrogantes, como por qué el material de El Cedral tiene el rango de talla más grande que cualquier otra muestra.

