











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introducción
La Formación Tuxpan comprende una sucesión de rocas terrígenas con sedimentos que se acumularon durante el Mioceno temprano-medio en diferentes subambientes litorales (Gío-Argáez, 1982; Vega et al., 1999). Sus múltiples afloramientos se encuentran a lo largo de la Planicie Costera del Golfo de México, desde la región de Tampico en Tamaulipas, hasta la región de Misantla en Veracruz. La mayoría de esos afloramientos contienen abundantes fósiles, principalmente bivalvos y gasterópodos (Perrilliat, 1994), asociados a restos de equinoideos, así como, diferentes tipos de anélidos, briozoos, crustáceos (Vega et al., 1999) y otros grupos de macro-invertebrados, así como restos de peces cartilaginosos y de mamíferos acuáticos, posiblemente sirenios. Dentro de esta diversidad, los equinoideos están representados por diferentes especies, tanto de formas exocíclicas como endocíclicas. Buitrón-Sánchez y Solís-Marín (1993) enlistan diversas especies de equinoideos que se han encontrado en la Formación Tuxpan y otras unidades litoestratigráficas del Paleógeno-Neógeno de Veracruz, sin precisar la edad y la localidad de origen. En particular para el género Plagiobrissus, no existen referencias previas que señalen su presencia en sedimentos de la Formación Tuxpan, ni en otras unidades del Cenozoico de México. Se puede considerar como una excepción la mención que hacen Dickerson y Kew (1917) sobre el hallazgo de “un espécimen de Metalia cumminsi en una localidad del terciario medio del estado de Tamaulipas”, acompañada de una descripción muy superficial que no permite reconocer los rasgos diagnósticos del género Metalia. Dicho ejemplar, asignado tentativamente al Mioceno de México y que se encuentra depositado como el Holotipo CASG # 371 en la colección de la California Academy of Sciences en San Francisco, California, fue analizado por Israelsky (1924) y reasignado al género Plagiobrissus, conservando el nombre específico. El mismo ejemplar fue analizado nuevamente por Lambert y Sánchez-Roig (Durham, 1961) y asignado al género Lajanaster. Considerando estos antecedentes, el informar acerca del hallazgo en rocas de la Formación Tuxpan de un espécimen de Plagiobrissus grandis, así como presentar su descripción, establecer con certeza la edad, localidad en que se encuentra y analizar las condiciones paleoambientales en que vivió, aporta información relevante sobre la historia biogeográfica y paleoambiental del género y permite explicar el patrón de distribución que presenta en la actualidad.
2. Área de estudio
2.1. localidad
El ejemplar estudiado se encontró en un afloramiento de la Formación Tuxpan, ubicado en el kilómetro 50.5 de la carretera 180 Poza Rica-Veracruz, en el tramo que une el poblado de Papantla con el de Gutiérrez Zamora, Municipio de Papantla, Veracruz, en las coordenadas 20° 26’ latitud N y 97° 06’ longitud W. Esta localidad es denominada La Pioja por encontrarse actualmente en las inmediaciones de la gasolinera del mismo nombre (Figura 1).
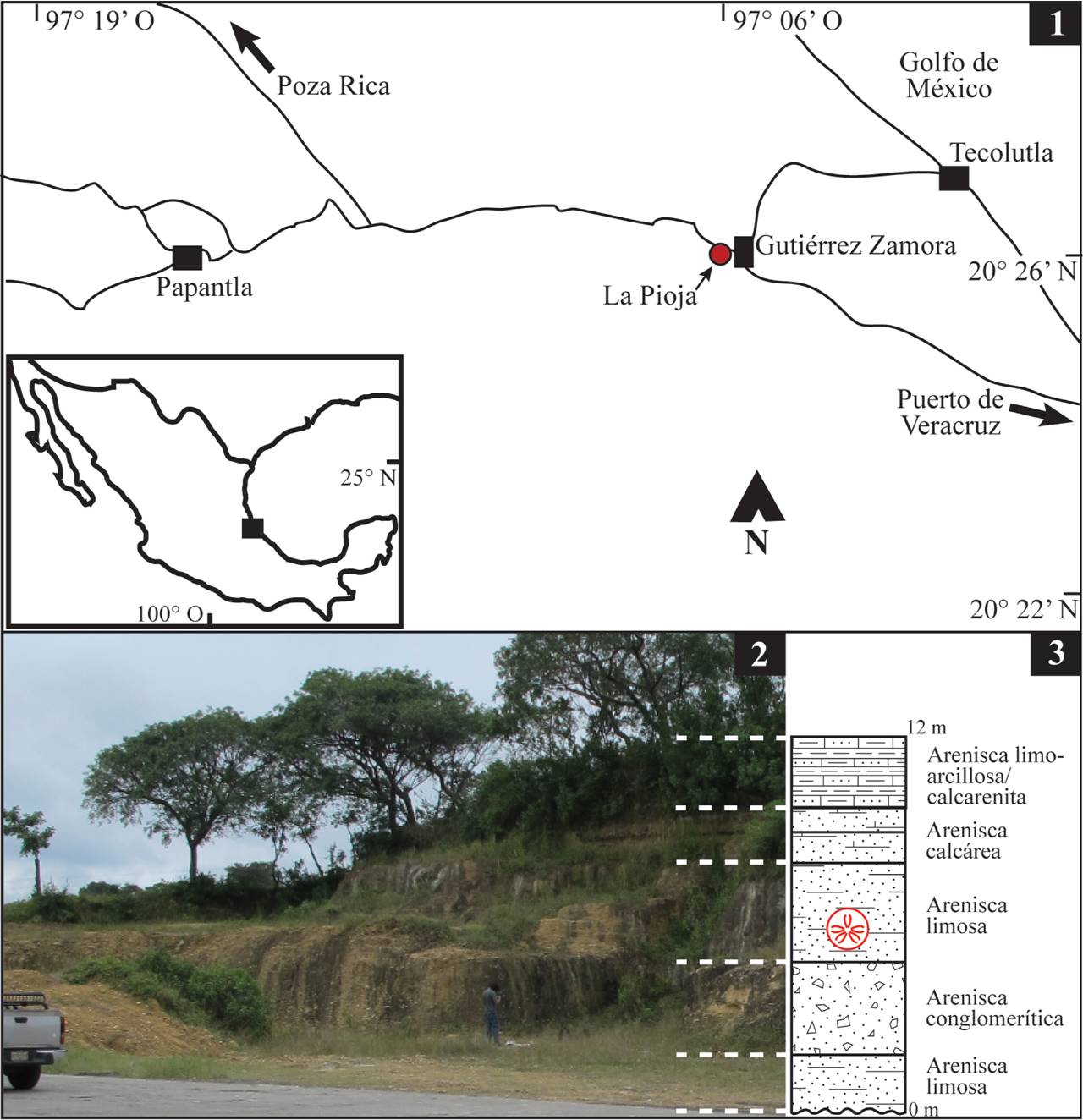
2.2. Marco geológico y estratigráfico
En la región que abarca el norte del Estado de Veracruz y el sur del Estado de Tamaulipas, el basamento cortical está pobremente expuesto en el flanco este de la Sierra Madre Oriental y consiste principalmente en rocas paleozoicas muy metamorfizadas que se correlacionan con el cinturón orogénico Apalache-Marathon-Ouachita. Sobre el basamento se encuentran rocas mesozoicas de origen marino, cubiertas por rocas de edades que abarcan desde el Paleoceno hasta el Plioceno. Los principales elementos tectónicos que controlaron el depósito y la deformación de la sucesión sedimentaria postpaleozoica son el Anticlinorio de Huayacocotla, el Macizo de Teziutlán, la Plataforma de Córdoba, el Macizo Volcánico de los Tuxtlas, la Cuenca Salina del Istmo y la Cuenca de Tampico-Misantla. Esta última incluye a la antefosa de Chicontepec, a la parte sur de la plataforma de Tamaulipas y el flanco este de la Sierra Madre Oriental, que en conjunto ocupan además de la región norte del Estado de Veracruz y sureste de Tamaulipas, las porciones más orientales de San Luis Potosí e Hidalgo, el norte de Puebla y la plataforma continental hasta su isobata de 200 m (PEMEX, 1988). La Cuenca de Tampico-Misantla tiene la forma de un arco semielíptico, abierto hacia el Golfo de México y está delimitada al norte por el Arco de Tamaulipas y el Homoclinal regional de San José de las Rusias, al oeste con la Sierra Madre Oriental y al sur por el Bloque tectónico de Teziutlán. El carácter de cuenca de esta región obedece al hundimiento del área ocupada por el archipiélago de Tamaulipas, provocado en gran parte por los esfuerzos que plegaron y fallaron la Sierra Madre oriental a principios del Paleógeno (Castro-Mora et al., 1994) y que favorecieron el depósito de los terrígenos que conforman a las unidades del Eoceno, Oligoceno y Mioceno que afloran a lo largo de la Planicie Costera del Golfo (Castro-Mora et al., 1994). En esta secuencia, la Formación Tuxpan representa la unidad más joven. Fue descrita originalmente por Dumble (1911) y redescrita por el mismo autor en 1918. Thalmann (1935) propuso como localidad tipo la sección de 150 metros que afloraba a unos 120 metros al oeste de la iglesia principal de la Ciudad de Tuxpan. Posteriormente, Barker y Blow (1976) analizaron la distribución de foraminíferos en las unidades litoestratigráficas de la Cuenca Sedimentaria Tampico-Misantla y señalaron una edad del Langhiano para la Formación Tuxpan. Actualmente esta unidad aflora en numerosas localidades formando una faja de aproximadamente 125 km de largo, con una anchura media de 15 km a lo largo de la costa del Golfo de México, desde el noreste del Cerro Azul, Veracruz hasta cerca de 20 km al sur del Río Tecolutla. Los afloramientos presentan espesores muy variables, alcanzando hasta 200 metros en los que predominan estratos de limolita arenosa que alternan con capas de arenisca, arenisca calcárea y caliza arenosa. Localmente se llegan a apreciar estratos de conglomerado que representan a la base de la formación. En los estratos arcillo-arenosos y arenosos de la Formación Tuxpan, como ya se mencionó, se ha encontrado una gran diversidad y cantidad de restos de invertebrados y vertebrados acuáticos. En conjunción con esta fauna, se ha referido la presencia de ostrácodos (Gío-Argáez, 1982) y foraminíferos (Carreño, 1986) que permiten asignar los estratos fosilíferos al Mioceno medio. Con la fauna fósil y considerando también las características litológicas, para esta formación es posible sugerir un depósito en ambientes litorales de latitudes tropicales, con facies marginales, supra, meso e infralitorales sin llegar a zonas marinas profundas (Vega et al., 1999). También se puede inferir que la unidad se depositó durante un evento transgresivo, seguido por la regresión que marcó la posición actual del margen costero del Golfo de México.
El ejemplar de Plagiobrissus que se describe, se encontró en posición de vida en estratos de la sección de la Formación Tuxpan que aflora en la localidad La Pioja (Figura 1). Dicha sección tiene un espesor de casi 12 metros, la parte basal se caracteriza por cerca de 2 metros de estratos de arenisca limosa con fauna típica de ambientes infralitorales seguidos por cerca de 2 metros de estratos de arenisca con clastos de grava y fragmentos de conchas, 3 metros de arenisca limosa fosilífera, 2.5 metros de arenisca calcárea y, en la parte superior, se encuentran estratos delgados, no fosilíferos de arenisca limo-arcillosa que alternan con estratos de calcarenita; todas las rocas son de color crema. Los rasgos tafonómicos que se observan en los restos fósiles indican que en gran medida fueron enterrados en posición de vida o después de un ligero transporte. En algunos estratos de arenisca se tiene el registro de abundantes restos esqueléticos, principalmente de moluscos mayormente desarticulados y fragmentados, cuyo origen se asocia al oleaje y a posibles tormentas que provocaron su acumulación en un ambiente de playa. El ejemplar de Plagiobrissus se encontró en la parte media de la sección, en un estrato de arenisca limosa (Figura 1).
3. Paleontología sistemática
Phylum Echinodermata Klein, 1754
Clase Echinoidea Leske, 1778
Orden Spatangoida Agassiz, 1840
Familia Brissidae Gray, 1855
Género Plagiobrissus Pomel, 1883
Especie tipo. Echinus grandis Gmelin, 1788, p. 320, por designación original. Reciente, Indias occidentales.
Diagnosis. Testa oval, con o sin surco anterior somero, deprimida en el ámbito. Sistema apical de tipo etmolítico, con cuatro gonoporos. Ambulacro anterior somero y ligeramente hundido; poros de igual tamaño, pareados simples y pequeños; placas adapicales con altura igual al ancho. Otros ambulacros estrechos y ligeramente hundidos, el par anterior forma un ángulo obtuso, el posterior se flexiona hacia el margen distal. Pétalos paralelos con un espacio perradial pequeño entre las columnas de poros pareados. Placas distales de los pétalos cerradas. Periprocto largo sobre una faceta corta y truncada. Peristoma ancho y largo, de forma arriñonada, con faceta oblicua hacia el frente. Placas labrales cortas y amplias y en amplio contacto con ambas placas esternales que son largas, estrechas y paralelas entre sí. Placas episternales largas y pareadas, con alrededor de dos tercios de la longitud de la placas esternales, formando la parte posterior del plastron. La sutura entre las placas esternales y episternales es posteriormente cóncava. Placas post-episternales pareadas. Zona posterior ambulacral, sobre la superficie oral, subparalela y estrecha. Placa ambulacral VI se expande, penetrando la fasciola subanal. Espinas y tubérculos primarios comúnmente dentro de la fasciola peripetaloide, formando hileras horizontales sobre el ambulacro I; tubérculos no deprimidos. Plastron totalmente tuberculado. Fasciolas subanal y peripetaloide bien desarrolladas. Fasciola subanal con forma de coraza, ramificaciones anales, con cuatro tubos pedicelares sobre cada lado.
Plagiobrissus grandis (Gmelin, 1788)
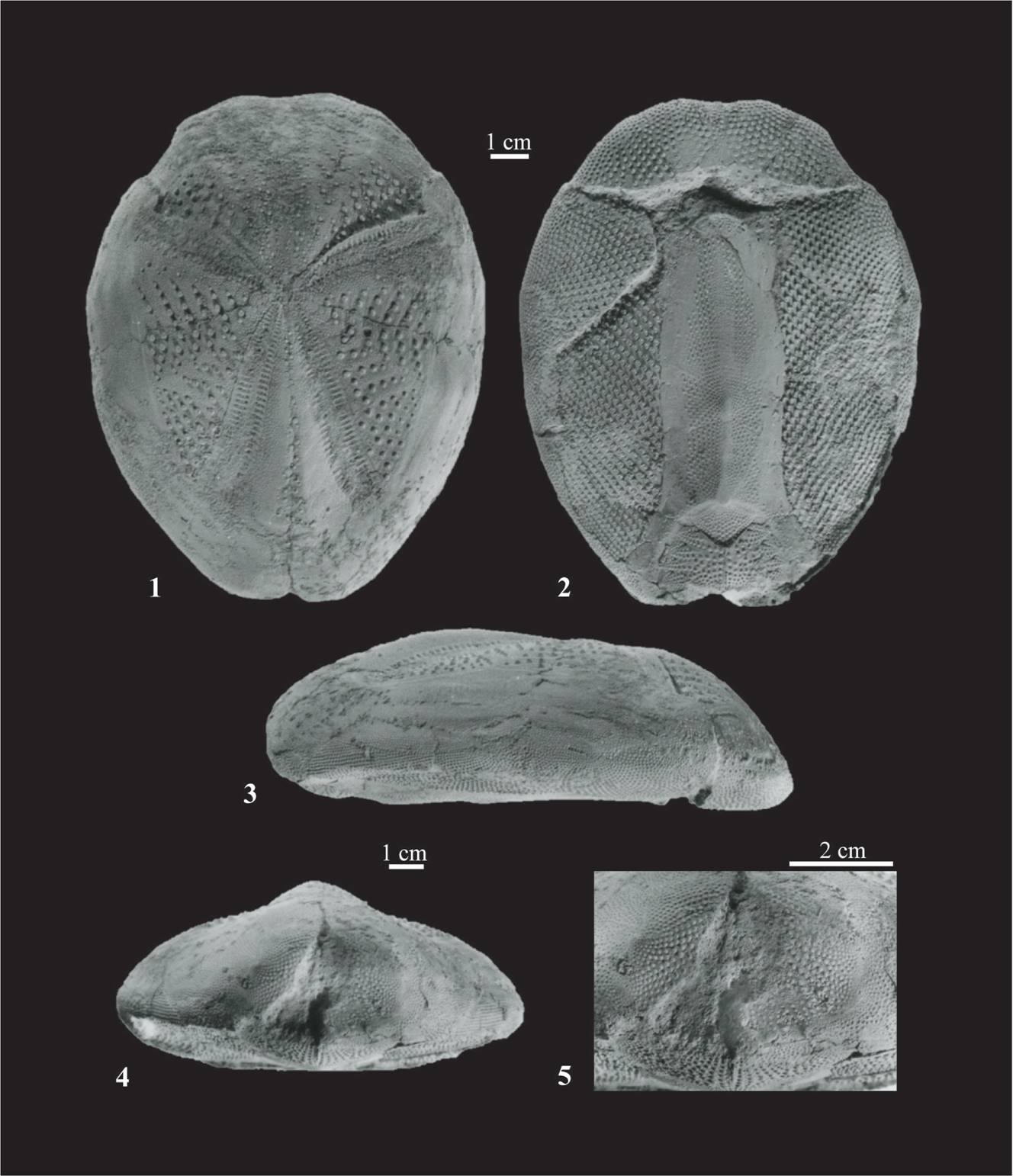
Figura 2. (1-5) Plagiobrissus grandis (Gmelin, 1788), ejemplar FCMP-1257: 1. Región aboral; 2 Región oral; 3. Vista lateral; 4. Vista posterior; 5. Aproximación de la zona anal.
Sinonimia.
Echinus grandis Gmelin, 1788, p. 3200.
Spatangus pectoralis Lamarck, 1816, p. 29.
Brissus (Plagionotus) pectoralis (Lamarck). Agassiz y Desor, 1847, p. 119, lám. 16, fig. 15.
Eupatagus pectoralis (Lamarck). D’Archiac, y Haime, 1853, p. 217.
Metalia pectoralis (Lamarck). Agassiz, 1872, p. 144, 600, lám. 21, figs. 4, 5.
Plagiobrissus pectoralis (Lamarck). Pomel, 1883, p. 29, 30.
Eupatagus (Plagiobrissus) grandis (Gmelin). Cooke, 1942, p. 54.
Plagiobrissus grandis (Gmelin). Clark, 1917, p. 207-208, lám. 146, figs. 21, 22; Mortensen, 1951, láms. 40, 41, 43 figs 13, 16; Cooke, 1959, p. 86, 87, lám. 39, figs, 1, 2; lám. 40, fig. 6.
Plagiobrissus (Plagiobrissus) grandis (Gmelin). Fischer, 1966, p. 602-603, fig. 484.4a,b
Descripción. Testa grande, subovalada en el ámbito, con surco anterior somero, elongada antero-posteriormente y plano-convexa en perfil lateral. Con una longitud antero-posterior de 136.31 mm, un diámetro transversal de 107.43 mm y altura apical de 39.19 y 46.16 mm sobre el peristoma y el esternón respectivamente. Superficie actinal plana desde los bordes, con presencia de un peristoma grande y semicircular; disco apical muy pequeño, mal conservado, no se observan las placas oculares a excepción de la G2 o madreporita, muy ancha posteriormente, forma que permite inferir la condición etmolítico del sistema apical que está en posición anterior y cubierto por múltiples poros. El ambulacro impar se encuentra en un surco somero, algo más profundo hacia la parte posterior del surco. Los ambulacros pareados son estrechos y ligeramente hundidos en un surco, las placas ambulacrales son simples, de forma rectangular, con el par de poros en posición posterior, pequeños, isopóricos y bien diferenciados. El par de ambulacros anteriores, II y IV, forman entre sí un ángulo divergente de 125 grados, el par posterior presenta una pequeña flexión hacia el margen distal. El ambulacro III es muy angosto, se extiende hasta el borde anterior de la región oral y está cubierto por tubérculos de diversos tamaños. Fasciola peripetaloidea no dentada en la región posterior a los pétalos, con tubérculos primarios pequeños, perforados y crenulados dispuestos en series horizontales sobre las placas interambulacrales. Adyacentes al ambulacro impar, tubérculos secundarios, prominentes y perforados; tubérculos subanales pequeños, perforados, crenulados y dispuestos en filas. Periprocto supramarginal rodeado de una fasciola. Peristoma y periprocto no se conservaron. Plastron metasterno, totalmente tuberculado, alargado y angosto con placa labral corta y ancha, poco diferenciada, par de placas esternales muy largas y par de episternales muy cortas con alrededor de un séptimo de la longitud de la placas esternales. La sutura entre las placas esternales y episternales es muy marcada y cóncava. Placas post-episternales pareadas, con filas simétricas y triangulares de tubérculos que forman una costelación aparente, tubérculos no deprimidos en la zona posterior ambulacral, sobre la superficie oral, sub-paralela y estrecha. Placa ambulacral VI se expande hacia la fasciola subanal que presenta forma de coraza con ramificaciones anales muy marcadas y cuatro tubos pedicelares sobre cada lado.
Material y ocurrencia. Ejemplar FCMP 1257, Localidad la Pioja, km 50.6 carretera 180, Papantla-Gutiérrez Zamora, Veracruz. Las siglas FCMP son las del Museo de Paleontología de la Facultad de Ciencias de la Universidad Nacional Autónoma de México.
Discusión. El ejemplar de Plagiobrissus de la Formación Tuxpan se distingue de las especies de Metalia por la presencia de tubérculos primarios grandes dentro de la fasciola peripetaloide. Es similar en su región aboral a Macropneustes Agassiz (Agassiz y Desor, 1847), pero en la región oral la disposición de las placas del plastron es muy diferente y Macropneustes carece de las placas pareadas episternales y post-episternales y la fasciola subanal no tiene forma de escudo. Rhabdobrissus (Cotteau, 1889), que es considerado un subgénero de Plagiobrissus, carece de surco frontal. De las especies que se han descrito para Plagiobrissus, P. grandis es muy similar a la especie actual P. africanus (Verrill, 1871) en dimensiones y rasgos de la región aboral, pero se puede distinguir de ella por presentar una testa ovada y más delgada, una fasciola peripetaloidea posteriormente no dentada, menos extendida y con tubérculos menos prominentes, un ambulacro anterior estrecho y poco conspicuo con poros pequeños e indiferenciados. Además, en P. africanus las placas episternales están separadas de las esternales por un surco que bordea, en conjunto, a las placas episternales y la fasciola subanal. Una distinción conspicua es la posición del periprocto que en P. grandis es visible desde un ángulo perpendicular a la región oral mientras que en P. africanus el periprocto sólo es visible posteriormente. Plagiobrissus robustus (Arnold y Clark, 1927), del Oligoceno de Jamaica y región caribeña, también presenta un gran tamaño, pero su testa es angosta, proporcionalmente más alta, el sistema apical se encuentra en posición anterior y los ambulacros son rectos. La gran dimensión de P. grandis permite distinguirle de otras especies del género. Además, la posición central del sistema apical distingue a P. grandis de las especies P. costaricensis (Durham, 1961) y P. malavassii (Durham, 1961), del Mioceno de la región central de Costa Rica. Las dimensiones, la posición del sistema apical y la presencia de una escotadura en la región anterior de la testa permite distinguir a P. grandis de las especies del subgénero Rhabdobrissus que se han descrito para el presente (Ghiold, 1989): Plagiobrissus (Rhabdobrissus) costae (Gasco, 1876) del Mediterráneo y Atlántico Norte, P.(R) jullieni (Cotteau, 1889), del Golfo de Guinea y Oeste de África, y P.(R) pacificus (Clark, 1940) que se encuentra desde el Golfo de California hasta las costas de Ecuador.
4. Discusión, Consideraciones biogeográficas
La distribución actual del género Plagiobrissus se restringe a mares tropicales someros del Hemisferio Norte y está caracterizada por la ocupación de un nicho infra-faunal, casi superficial, en el que los equinoideos del género se mueven lentamente alimentándose de detritos. En particular P. grandis se encuentra en ambientes someros del Golfo de México y Caribe hasta las costas de Venezuela y Brasil. La distribución geográfica y temporal de otras especies del género se detalla en la Tabla 1, cuyo análisis permite inferir que el origen de Plagiobrissus ocurre en el Eoceno medio, en la región norte-caribeña en donde se ha encontrado a P. robustus en Jamaica y la región del Caribe (Arnold y Clark, 1927), P. loveni en Cuba (Sánchez-Roig, 1953), P. curvus en Florida (Oyen y Portell, 2001), y a P. dixie en Florida y Alabama (Toulmin, 1977; Oyen y Portell, 2001). Durante el Mioceno, Plagiobrissus se expande hacia el sur, hasta las costas de Costa Rica y Brasil, con las especies P. costaricensis y P. malavassii (Durham, 1961) y hacia el Golfo de México con P. grandis que ahora se refiere para México. Entre finales del Mioceno y principios del Plioceno, antes del cierre del Istmo de Panamá, el linaje de P. pacificus coloniza las costa americanas del Pacífico. El cierre del canal de Panamá provoca drásticos cambios en el patrón de corrientes del Atlántico Norte y favorece la dispersión, a lo largo de los últimos 5.3 millones de años, del género hacia el Mediterráneo, donde actualmente se encuentra a Plagiobrissus costae, y hacia la costa noroccidental de África, en que se tiene a P. africanus y P. jullieni. Considerando que el origen del linaje Plagiobrissus se marca a finales del Eoceno, su historia evolutiva y biogeográfica se enmarca en un periodo de alrededor de 33 millones de años. En esta historia resalta el hecho de que las diversas especies del género, entre ellas Plagiobrissus grandis, con una edad de por lo menos 14 millones de años, presentan tasas de duración muy altas. Evolutivamente también es notable que todos los eventos de especiación que se registran para el grupo fueron determinados por cambios geográficos y que el papel del posterior aislamiento solo se hace evidente en cambios morfológicos poco conspicuos. Un ejemplo de ello es la poca diferenciación que existe entre Plagiobrissus grandis y su descendiente P. africanus cuya divergencia debió iniciar por lo menos hace cinco millones de años.
Tabla 1. Distribución geográfica y estratigráfica de las especies del género Plagiobrissus, incluyendo las del subgénero Rhabdobrissus.
| Especie | Edad | Localidades | Referencia |
|---|---|---|---|
| P. (R. ) costae | Reciente | NE Atlántico, Mediterráneo-extremo noroeste de África hasta la costa del norte de Sierra Leona, y zonas profundas del Atlántico de África Central. | Ghiold, 1989 |
| P. africanus | Reciente | Costa de África Central (Costa de Marfil-Guinea Ecuatorial, incluyendo el Golfo de Guinea | Ghiold, 1989 |
| P. (R. ) pacificus | Reciente | Pacífico Sureste (Costa pacífica de México-Centroamérica) | Ghiold, 1989 |
| P. (R. ) jullieni | Reciente | Atlántico Sureste; Golfo de Guinea (Costa de Ghana-Togo, Benin, Lagos y Nigeria) | Ghiold, 1989 |
| P. sp. | Reciente | Oeste del Indo-Pacifico: Islas de Oceanía (Mar de Timor, norte de Australia) | Ghiold, 1989 |
| P. grandis | Reciente | Golfo de México-Caribe, Cuba, Costa de Venezuela-Brasil | Ghiold, 1989 |
| Mioceno | Brasil | Couto, 1967; Araújo-Távora et al. , 2010; * | |
| P. sp. | Plioceno | Florida | Oyen y Portell, 2001 |
| P. costaricensis | Mioceno | Región central de Costa Rica | Durham, 1961 |
| P. malavassii | Mioceno | Región central de Costa Rica | Durham, 1961 |
| P. curvus | Eoceno | Florida | Oyen y Portell, 2001 |
| P. dixie | Eoceno | Alabama y Florida | Toulmin, 1977; Oyen y Portell, 2001 |
| P. sp. | Eoceno | Estados Unidos (Carolina del Norte) | Kier, 1980 |
| P. loveni | Eoceno medio | Cuba | Sánchez-Roig, 1953 |
| P. robustus | Eoceno medio | Jamaica, Bahamas, Cuba | Arnold y Clark, 1927; Sánchez-Roig, 1953 |
*en este trabajo.
5. Conclusiones
La presencia de Plagiobrissus grandis en el Mioceno (Langhiano) de la Planicie Costera del Golfo, además de representar el primer registro fósil del género en México, permite corroborar su origen en el Mar Caribe, región desde la cual se dispersó durante el Plioceno-Reciente hacia las costas mediterráneas y del norte de África, hasta el Golfo de Guinea. La formación del Istmo de Panamá durante el Mioceno-Plioceno, y los cambios que provocó en las corrientes del Atlántico Norte, fueron un factor determinante en ese evento de dispersión. Plagiobrissus grandis, con una existencia de por lo menos 14 millones de años, es un ejemplo de las tasas de duración de gran extensión que presentan diversas especies de equinoideos.

