











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink1. Introducción
El ámbar mexicano, localizado en el municipio de Simojovel de Allende, Chiapas; fue producido por una especie de árbol de la familia Fabaceae, Hymenea mexicana (Poinar y Brown, 2002, citado en Bickel y Solórzano-Kraemer, 2016). La resina fosilizada en esta región ha sido datado con una edad geológica de 23 Ma, correspondiente al Mioceno Inferior (Aquitaniano) (Vega et al., 2009; Perrillat et al., 2010; Serrano-Sánchez et al., 2015). Simojovel constituye uno de los depósitos ambarinos más importantes del mundo y en él se han encontrado alrededor de 200 familias de artrópodos (Solórzano-Kraemer, 2010). Muchas de las investigaciones realizadas con inclusiones biológicas en ámbar de esta región, han sido centradas en los diferentes Órdenes de la Clase Insecta (e.g.Macadam y Ross, 2016; Popov, 2016; Ross et al., 2016a, 2016b; Heiss, 2016); otros más con crustáceos (e.g.Serrano-Sánchez et al., 2016), plantas (e.g.Hernández-Damián et al., 2016) y arácnidos (e.g.Rivas et al., 2016; Judson, 2016), entre estos últimos las arañas (Petrunkevitch, 1963, 1971; García-Villafuerte y Penney, 2003; García-Villafuerte 2004, 2006a, 2006b, 2007, 2008). Actualmente, el órden Araneae se compone de 112 familias, la más numerosa de ellas es Salticidae (World Spider Catalog, 2018).
Los miembros de la familia Salticidae están presentes en todos los continentes con excepción de la Antártida, dentro de una amplia gama de micro hábitats, desde la hojarasca hasta la copa de los árboles (Richman et al., 2005). Dado su amplio rango de distribución es la familia con mayor riqueza de especies dentro del órden Araneae, representando el 10.68 % del total de especies de arañas válidas registradas para el mundo (World Spider Catalog, 2018). A nivel mundial, se han registrado un total de 6077 especies actuales, distribuidas en 634 géneros (World Spider Catalog, 2018). El conocimiento de la riqueza de la fauna de arañas saltadoras para México se conoce a través de los trabajos realizados por Hoffman (1976), Jiménez (1996), Castelo-Calvillo (2000) y Richman et al. (2011); a la fecha se han reportado 264 especies distribuidas en 65 géneros para el país (Richman et al., 2011). En lo que respecta al estado de Chiapas se han registrado 25 especies válidas, incluidas en 14 géneros (Richman et al., 2011; Garcilazo-Cruz y Álvarez-Padilla, 2015).
En lo que concierne al registro fósil de Salticidae, se enlistan las especies en Dunlop et al. (2018), las cuales han sido reportadas para los diferentes depósitos ambarinos; destacando principalmente el Báltico y la República Dominicana. Para el caso del ámbar mexicano los registros son escasos, incertae sedis (Petrunkevitch, 1971), Lyssomanes sp. (García-Villafuerte y Penney, 2003), Maevia? eurekaRiquelme y Menéndez-Acuña, 2017, gen. sp., indet. (García-Villafuerte, 2004) y cinco gens. sp., indeterminados, que se encuentran resguardados en museos extranjeros (Ross et al., 2016b).
En el presente estudio se reporta una nueva especie fósil del actual género GalianoraMaddison, 2006, de la tribu Lapsiini perteneciente a la subfamilia Spartaeinae, encontrada en ámbar de Chiapas, México. Hill y Richman (2009) mencionan que aunque muchos grupos de salticidos basales se encuentran en los trópicos del Viejo Mundo y en las Indias Orientales en particular; otros, incluyendo muchos lyssomaninos y lapsinos actuales, están ampliamente distribuidos en los trópicos del Nuevo Mundo.
La presencia de un lapsino en el ámbar de Chiapas, permite reflexionar puntos importantes, como detalles taxonómicos de la familia, aspectos biogeográficos del género y su posible permanencia en la fauna actual, también se discute el tema del origen de la familia Salticidae y su divergencia; finalmente, con base en el registro fósil y actual de arañas, se plantea la idea de que los géneros araneidos, aparentemente extintos, pueden permanecer en la región, y su probable sobrevivencia a la evolución geológica de América Central.
2. Área de Estudio y Método
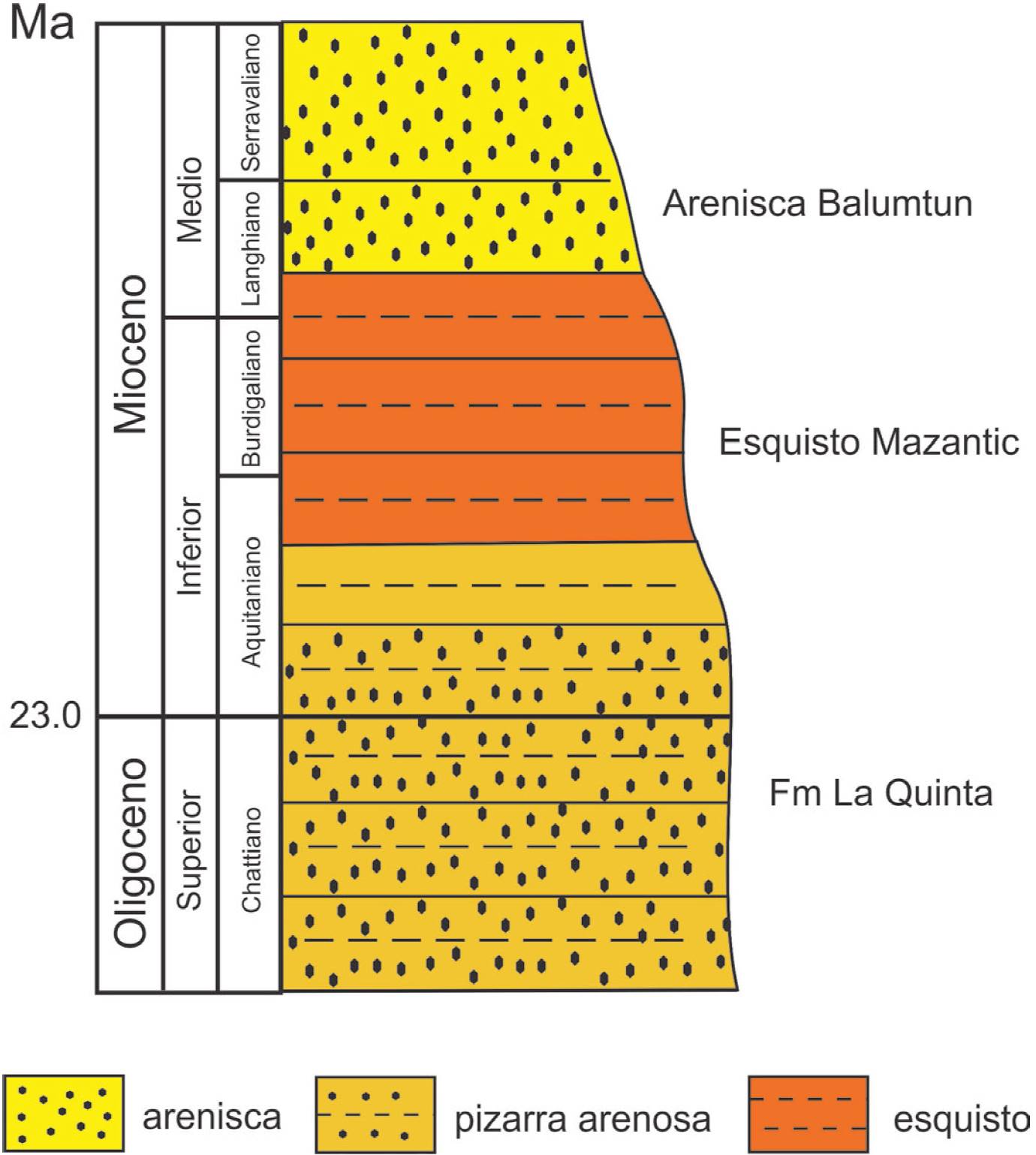
La pieza de ámbar estudiada proviene del municipio de Simojovel de Allende en el estado de Chiapas, que está ubicado a 50 km de la ciudad de Tuxtla Gutiérrez, al norte del estado y al sur de México (Figura 1). En el área, la resina fósil es extraída a través de túneles de depósitos marinos. Las minas principales son los Pocitos, Campo La Granja y El Pistón, donde interactúan tres unidades litoestratigráficas: Formación La Quinta, Mazantic y Balumtum (Figura 2); éstas unidades están expuestas en la Sierra Madre del Sur de Chiapas, desde el borde de la Depresión Central (Totolapa) hasta Palenque, cerca de las costas del Golfo de Tabasco (Alisson, 1967; Frost y Langenhein, 1974; Perrillat et al., 2010; Riquelme et al., 2014; Serrano-Sánchez et al., 2015).
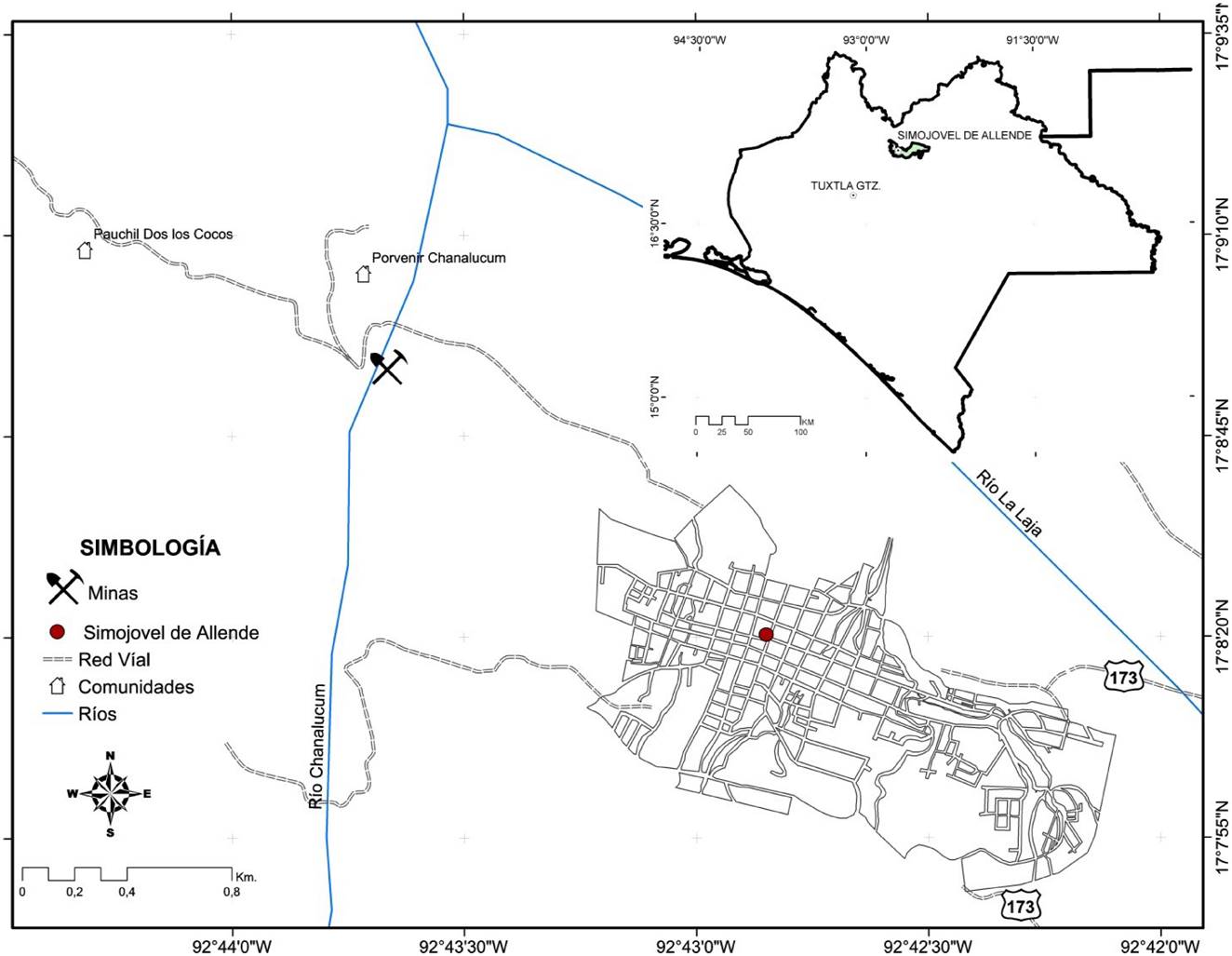
El espécimen fósil en ámbar proviene de la mina los Pocitos, fue adquirida directamente con un artesano de la localidad por el Biólogo Marco Antonio Coutiño José, quien la donó al Museo de Paleontología “Eliseo Palacios Aguilera”, Dirección de Paleontología de la Secretaria de Medio Ambiente e Historia Natural (SEMAHN), donde actualmente se resguarda bajo el número de catálogo: IHNFG-5494 (Instituto de Historia Natural, Fósil Geográfico).
Para obtener una mejor visualización del espécimen, se utilizaron lijas de diferentes medidas, con el fin de aminorar el tamaño de la pieza y finalmente, se utilizó un abrasivo de la marca Brasso®, para pulir la pieza, que entre sus múltiples usos está el eliminar y reparar arañazos en los plásticos, CD’s, DVDS, pantallas y piscinas; el uso de este material en el ámbar ha demostrado que elimina las ralladuras que deja la lija más fina. Las fotografías fueron realizadas con una cámara Leica MC170HD conectada a un microscopio estereoscópico Leica M205C. Utilizando la técnica de apilamiento se realizaron una serie de tomas de las estructuras bajando el enfoque gradualmente; posteriormente, con el software Helicon focus 6, las fotografías se sobrepusieron para lograr una composición con amplia profundidad de campo y enfoque; las imágenes se importaron a Ligth room 5.5 para el proceso de revelado. La descripción del espécimen se da con base en lo propuesto por Galiano (1963) y Maddison (2006). Las medidas son dadas en milímetros y fueron tomadas con una gratícula ocular adaptada a un Estereoscopio Zeiss Stemi 2000. Abreviaturas: AO (área ocular), OMA (ojos medios anteriores), OLA (ojos laterales anteriores), OMP (ojos medios posteriores), OLP (ojos laterales posteriores), Ci (cimbio), E (émbolo), Am (apófisis media), Te (tegulum), Bc (pestaña del cimbio), fe (fémur), pat (patela), tib (tibia), mt (metatarso), es (espina), r (retrolateral), p (prolateral), pa (prolateral apical).
3. Paleontología sistemática
Órden Araneae Clerck, 1757
Familia Salticidae Blackwall, 1841
Genero GalianoraMaddison, 2006
Especie tipo: Galianora sachaMaddison, 2006
Galianora marcoi sp. nov. (Figuras 3a-f).

Figura 3. Galianora marcoi sp. nov., a: vista dorsal; b: vista ventral; c: vista lateral, escala: 1 mm; d: Palpo, vista prolateral; e: Palpo, vista dorsal; f: dibujo de Palpo, vista prolateral, escala: 0.3 mm.
Material examinado. Holotipo macho IHNFG-5494 en ámbar de Simojovel de Allende, Chiapas México. Mioceno (Aquitaniano) - 23 Ma.
Diagnosis. Galianora marcoi sp. nov., se diferencia de las dos especies actualmente descritas, Galianora bryicolaMaddison, 2006 y G. sachaMaddison, 2006, por la forma de la apófisis medía que es alargada, recta, en forma de muleta; la punta del émbolo es más grueso, semibifurcado, se erige distalmente en el lado retrolateral y por la presencia de una pestaña retrolateral en la base del cimbio en forma de cuernos.
Etimología. El epíteto específico es un nombre en aposición y es dedicado a Marco Antonio Coutiño José (Museo de Paleontología “Eliseo Palacios Aguilera”), por la importante labor en la adquisición de piezas significativas de ámbar con inclusiones biológicas, las cuales ha donado al Museo de Paleontología antes mencionado, y por su interés en el conocimiento de la araneofauna fósil chiapaneca.
Descripción del macho holotipo (Figuras 3a-f). Longitud total 3.72. Longitud prósoma 1.76, ancho 0.98, alto: 0.58; AO: largo 0.84, línea anterior de ojos: 1.03 ancho, línea posterior ojos 0.78; estría torácica alargada. Distancia entre OLA y OMP 0.14; distancia entre OMP y OLP 0.21; altura del clípeo: es difícil tomar la altura por la posición de la araña, pero se observa un clípeo alto. Diámetro de OMA 0.29: Patas IV, I, III, II aproximadamente. Quetotaxia: I fe1-1 d; pat 0; tib 1-2-2 v, 1pt; mt 2-2 v. II fe 1-1d; pat 0; tib 1-1-1 r; mt 2-2v, con un verticilio apical de 4 es. III fe 1-1-1d, 1pa, pat 1p; mt 1v, 1-1r, 1p, con un verticilio apical de 4 es. IV fe 1-1d, pat 1r; tib 1-1r, 1-1p; mt 1-1-1p. Palpo: fe 0.58, pat?, tib 0.31, ci 0.54; el cimbio tiene una forma cónica, la punta se observa esclerozada con un patrón de setas medianas agrupadas de tal manera que tienen la apariencia de un estilo punk en una vista lateral (Figura 3b). En vista dorsal se observa una pestaña en la base retrolateral del cimbio, que tiene la forma de dos cuernos uno más grande que otro (Figura 3e). Prolateralmente se logra observar un tegulum redondeado, el émbolo periférico distintivo del género y la apófisis media delgada y alargada, recta, en forma de muleta, 0.089 de longitud (Figuras 3d y f), dorsalmente se observa el extremo distal del émbolo de manera ensanchada y semibifurcada (Figura 3e). La ART tiene forma de un cáliz y está fuertemente esclerosado. Opistosoma: 1.69 de longitud.
Aspecto y color en ámbar. el prósoma no es muy elevado y presenta un suave declive torácico, con una fóvea larga. En el centro hay una banda ancha, con setas amarillentas y blanquecinas, que nace muy cerca de la base de los OMP y desemboca cerca del pedicelo terminando en punta; ambos costados de la base del prosoma de color negro y de la parte media hacia la línea de ojos anteriores de color pardusco. En la base de los OLP se observan sedas de color anaranjado oscuro, el mismo color se presenta en la base de los OMP hacia los OMA; los quelíceros son de color café claro; el opistosoma es de color pardo amarillento y todas las patas de color amarillo con ciertos tonos moteados pardos.
Hembra: desconocida.
4. Discusión
4.1. Detalles taxonómicos
El espécimen estudiado ha sido ubicado dentro de la familia Salticidae, ya que las arañas saltadoras se caracterizan por la presencia de grandes ojos medios anteriores (OMA) (Scheuring, 1914; Ramírez, 2014) y por el característico arreglo de los ojos (Maddison, 2015). Penney (2010) menciona que esta característica es muy distintiva en los salticidos por lo que pueden identificarse incluso a partir de especímenes juveniles, aun cuando se encuentran en la resina fosilizada. Coddintong y Levi (1991) señalan que la sinapomorfia más sobresaliente de la familia es el par de ojos medios anteriores altamente especializados.
Hill y Richman (2009) mencionan que el desarrollo único de la visión en Salticidae ha respaldado la evolución de una gama extraordinariamente diversa de estilos de vida, todos asociados con un comportamiento visualmente mediado. Los salticidos son miembros del clado ART (por la apófisis retrolateral en los machos), este clado incluye por lo menos la mitad de las arañas conocidas (Blackledge et al., 2009) y con pocas excepciones, la familia está entre los miembros del clado que aparentemente perdieron la capacidad de construir telarañas (Hill y Richman, 2009).
Determinar si una araña pertenece o no a la familia Salticidae es relativamente sencillo, pero la ubicación taxonómica de un espécimen dentro de una subfamilia, género o especie en particular, requiere una detallada observación, no solo de los caracteres somáticos, sino también de las estructuras genitales, sean machos o hembras. Harms y Dunlop (2008) mencionan que las arañas fósiles en ámbar a menudo se asemejan incluso a géneros y especies actuales y por lo general se pueden ubicar dentro de familias actuales. Pero también son difíciles de comparar con otros fósiles o especies cercanas, debido a que una observación clara de los genitales (palpo, en los machos y/o epiginio, en las hembras) a menudo es difícil de distinguir.
En efecto, los paleoaracnólogos se enfrentan a una serie de dificultades al estudiar arañas fósiles incluidas en ámbar; por ejemplo, la posición en la cual quedan fosilizados los especímenes, las fracturas y burbujas que obstruyen la visibilidad de estructuras clave en la determinación taxonómica, etc. En este caso, los palpos del espécimen estudiado se encuentran doblados hacia dentro, de tal manera que el dorso del cimbio se observa ventralmente (Figura 3b). Al principio de la investigación esto dificultó su determinación taxonómica; consecuentemente, se tuvo que lijar y pulir la pieza por la parte posterior, del lado del opistosoma, teniendo el cuidado de no dañarla a fin de observar las estructuras genitales. Al hacer esto, se logró visualizar uno de los palpos en una vista prolateral, observando así el tegulum, el émbolo y la apófisis media (Figuras 3d y 3f). Harms y Dunlop (2008) mencionan que los caracteres del pedipalpo masculino en las arañas, representan la característica taxonómica más importante para su determinación. De la misma manera, Prószyński (2017) menciona que los palpos son estructuras conservadoras, estables, que caracterizan amplios ‘grupos de géneros’, con pequeñas modificaciones en los detalles en relación con géneros y especies. Estas son las únicas estructuras que permiten, apenas dándoles un vistazo, la identificación de subfamilia o géneros.
A continuación se argumenta porqué el espécimen fósil estudiado se ha ubicado dentro del actual género Galianora. Después de comparar el ejemplar con claves taxonómicas de arañas actuales para el norte de América (Richman et al., 2005), consultar diferentes bibliotecas ilustradas sobre salticidos (e.g.Prószyński, 2015; Metzner, 2015) y visitar el catalogo mundial de arañas (World Spider Catalog, 2018); se consideró el trabajo de Maddison (2015), quien realiza una clasificación filogenética de las arañas saltadoras con base en análisis morfológicos y moleculares, un trabajo valioso para el estudio de la evolución de salticidos (Prószyński, 2017). Maddison (2015) establece siete subfamilias: Onomastinae, Asemoneinae, Lyssomaninae, Spartaeinae, Eupoinae, Hisponinae y la más numerosa de ellas, Salticinae.
Con base en la descripción de cada una de las subfamilias, Galianora marcoi sp. nov., fue excluida de Onomastinae, Asemoneinae y Lysomaninae ya que carece de la translucencia verde o amarilla que identifica a las antes mencionadas, así como el característico patrón de ojos que diferencia a sus miembros. También fue descartada de la Subfamilia Eupoinae porque no tiene un cuerpo pequeño brillante y una escuta en el dorso del abdomen. Se separó de la subfamilia Hisponinae debido a que no presenta el surco o constricción transversal en el caparazón, justo detrás de los ojos medios posteriores (ver figura 23 en Maddison, 2015). También fue diferenciada de la subfamilia Salticinae porque el palpo de Galianora marcoi sp. nov., presenta una apófisis media; por lo tanto, se ubicó como miembro de la Subfamilia Spartaeinae.
Maddison (2015) menciona que las especies que componen a Spartaeinae, no tienen sinapomorfias morfológicas conocidas y aparentemente están unidos por estados de carácter posiblemente ancestrales; sin embargo, señala que los subgrupos o tribus que la componen, se encuentran claramente definidos por un rango geográfico, más que por su morfología. La subfamilia Spartaeinae se compone de tres tribus, una de ellas es Sparteini, principalmente de distribución africana, asiática, con pocos representantes en Europa y Australasia; la Tribu Cocalodini, cuyas especies (a excepción de Depreissia) están restringidas a Australasia, al este de la línea de Wallace (Wanless, 1982; Maddison, 2006) y son comunes en la araneofauna de Nueva Guinea, con variadas formas corporales (Maddison, 2006 citado en Maddison, 2015). Galianora marcoi sp. nov., fue descartado de estas dos tribus debido a que las investigaciones hasta hoy realizadas (García-Villafuerte y Penney, 2003; García-Villafuerte, 2006a, 2006b, 2008) indican que la araneofauna fósil en el ámbar de Chiapas es similar a la reciente fauna Neotropical. Además, se revisaron los artículos relacionados a los géneros de cada tribu, que presentan una apófisis media en el palpo, para comparar la estructura genital con el espécimen estudiado, y existen diferencias significativas (e.g.Wanles, 1982, 1984; Deeleman-Reinhold y Floren, 2003; Logunov y Azarkina, 2008; Prószyński y Deeleman-Reinhold, 2012; Maddison et al., 2014; Prószyński, 2017 y otros en: World Spider Catalog, 2018).
La tercer tribu, llamada Lapsiini, tiene una distribución Neotropical en el Nuevo Mundo y aunque no hay sinapomorfías que distingan a este grupo (Maddison, 2015), las especies que la integran, al igual que los lyssomaninos, comparten un rasgo ancestral adicional, la presencia de una apófisis media en el palpo del macho y la uña en los palpos de las hembras, lo cual no se observa en otras especies de salticidos neotropicales (Maddison 2006). Por otro lado, se revisó la descripción de las especies fósiles de arañas saltadoras con apófisis media en el palpo, según descritas en Wunderlich (2004a), que han sido registradas tanto para el Báltico como para la República Dominicana y Galianora marcoi sp. nov., muestra marcadas diferencias en cuanto a la posición de la apófisis media y la forma del émbolo y el tegulum. Por lo tanto, se llegó a la conclusión que el espécimen estudiado forma parte de la subfamilia Spartaeinae, como miembro de la tribu Lapsiini, siguiendo lo establecido por Maddison (2015).
La tribu Lapsiini, contiene 20 especies válidas distribuidas en cinco géneros actuales, GalianoraMaddison, 2006; LapsamitaRuiz, 2013; LapsiasSimon, 1900; SoesiladeepakiusMakhan, 2007 y ThrandinaMaddison, 2006 (Maddison, 2015; Prószyński, 2017; World Spider Catalog, 2018).
De los géneros que componen a los lapsinos, Galianora marcoi sp. nov., se corrobora con la descripción del género Galianora, ya que se diagnostica entre los salticidos neotropicales por poseer un tegulum redondeado con un émbolo periférico (Figura 3d y 3f). Las hembras son reconocidas por la presencia de una uña en los palpos. Se llegó a la misma conclusión, según lo establecido por Prószyński (2017), sin embargo, él menciona que las hembras se caracterizan por tener un epiginio con espermatecas y ductos muy elaborados o complicados.
Para este género se han descrito dos especies: Galianora bryicolaMaddison, 2006 y G. sachaMaddison, 2006. Galianora marcoi sp. nov., tiene un gran parecido, en la forma del cuerpo, con G. bryicola, pero se diferencia de esta por la forma del émbolo, la apófisis media y la ART. Por otro lado, el émbolo es muy parecido a G. sacha; sin embargo, Galianora marcoi sp. nov., se diferencia de ella por la forma de la apófisis media, la cual es más larga y delgada, en forma de muleta y no tiene el cuerpo alongado, sino más bien compacto. Y principalmente, por la presencia de una pestaña retrolateral en la base del cimbio en forma de cuerno (Figura 3e), se establece Galianora marcoi sp. nov. Al parecer, Galianora marcoi sp. nov., es una especie intermedia en la forma del cuerpo y el palpo.
4.2. Aspectos Biogeográficos
Después de la descripción del género tipo, LapsiasSimon (1900), ninguna otra especie fue correctamente agregada a la tribu Lapsiini por más de un siglo (Maddison, 2006). De acuerdo con Maddison (2015), los lapsinos neotropicales están confinados al Nuevo Mundo; miembros del género Lapsamita han sido reportados para Brasil, el género Lapsias para Ecuador y Venezuela, Soesiladeepakius para Suriname y Brasil, Thrandina tiene presencia en Ecuador (Maddison, 2006).
Hasta hoy día, no se había reportado al género Galianora en el registro fósil, y tampoco se tiene contemplada para el catálogo de especies actuales en el Estado de Chiapas (García-Villafuerte, 2009; Ibarra-Núñez, 2013) o en el listado a nivel Nacional en México (World Spider Catalog, 2018). El único estudio a la fecha sobre las especies que lo componen, las ubica geográficamente en Ecuador (Maddison, 2006; World Spider Catalog, 2018) y algunas especies aún no descritas para Venezuela (Maddison, 2006). La presencia de Galianora marcoi sp. nov., en el ámbar de Chiapas, proporciona información paleobiogeográfica importante sobre las actuales especies sudamericanas, ampliando el rango de distribución del género hacia el norte de la región Neotropical. Un caso similar ocurre con Maevia? eureka, una especie de araña de la subtribu Marpissina, encontrada en el ámbar de Totolapa, cerca de Simojovel; la cual extiende el rango geográfico del género, actualmente restringido a Norteamérica, hacia una región meridional. Tampoco este género ha sido reportado en el listado de especies actuales del órden Araneae para Chiapas (García-Villafuerte, 2009; Richman et al., 2011; Ibarra-Núñez, 2013).
4.3. ¿Géneros extintos?
Riquelme y Menéndez-Acuña (2017) sugieren, por la presencia de Maevia? eureka en el ámbar de Totolapa (Chiapas), y que coincide con otro registro actual al norte de México (Tamaulipas), M. poultoniPeckham y Peckham, 1901 (Richman et al., 2011), que muy probablemente el género Maevia? se dispersó a lo largo de los neotrópicos en el Mioceno y que finalmente se extinguió en el sur, manteniéndose casi exclusivamente en la región Neártica.
Esta sugerencia es sin duda precipitada; en primer lugar, Penney et al. (2003) mencionan el caso de la familia Archaeidae, la cual fue descripta por primera vez a partir de registros fósiles en el ámbar del Báltico por Koch y Berendt (1854); posteriormente en 1881, O.P. Cambridge descubre un archaeido vivo en Madagascar, Eriauchenius workmani O. Pickard-Cambridge, 1881 (Wood, 2008). Más de un siglo después se han registrado nuevas especies actuales para esta familia, que se consideraba extinta, en Madagascar, Sudáfrica y Australia (World Spider Catalog, 2018). En segundo lugar, debido a la complejidad y diversidad de ambientes, el estudio de la araneofauna actual en Chiapas es aún insuficiente y hasta la fecha no se tienen registros válidamente reportados de arañas para los municipios chiapanecos de Simojovel y Totolapa; y aún queda mucho territorio por muestrear en el Estado (ver mapa 1 en García-Villafuerte, 2009 y el último listado araneofaunístico en Ibarra-Núñez, 2013). En tercer lugar, sobreviene el caso del género Hemirrhagus Simon, 1903, una tarántula de la familia Theraphosidae, a quien en un principio se registró como representante de la araneofauna fósil chiapaneca (García-Villafuerte, 2008) y posteriormente, como parte del proyecto “Fortalecimiento de la Colección de Arácnidos de la Universidad de Ciencias y Artes de Chiapas”, se reportó la presencia del género dentro del listado de arañas actuales para Chiapas (García-Villafuerte, 2009), y que resultó ser una nueva especie para la ciencia, Hemirrhagus perezmilesiGarcía-Villafuerte y Locht, 2010 ¿Podría ocurrir el mismo caso con Maevia?, Galianora y los géneros aparentemente extintos reportados por Petrunkevitch (1963, 1971)? ¿Es posible que hoy día haya representantes actuales de estos géneros en Chiapas?
Por lo tanto, podríamos esperar que representantes de los géneros Maevia? y Galianora, reportados como registros fósiles en el Estado de Chiapas, se encuentren actualmente en esta región del mundo, como sucede con los géneros Hemirrhagus, Episinus, Selenops y Lyssomanes (García-Villafuerte y Penney, 2003; García-Villafuerte, 2006a, 2006b, 2008, 2009; García-Villafuerte y Locht, 2010), los cuales tienen representantes en la actual araneofauna chiapaneca, aun después de 23 Ma. Pero para constatar esto, es importante realizar investigaciones sobre la riqueza de arañas actuales aledañas a las minas de Simojovel y otros municipios; probablemente estos géneros aun ocupan su hábitat, manteniéndose en el tiempo desde el Mioceno (Aquitaniano) a la fecha.
4.4. Origen y divergencia de Salticidae
El origen de la familia Salticidae fue tema de debate; se pensaba que la familia se había originado en el Mesozoico (durante el Cretácico), con base en los registros publicados por Grimaldi et al. (2002), quienes registraron una araña saltadora para el ámbar de Nueva Jersey; al igual que Néraudeau et al. (2002), al publicar la existencia de Salticidae en el ámbar de Francia y por un espécimen reportado como salticido por Kaddumi (2005) para el ámbar de Jordania (Penney et al., 2003); sin embargo, Penney (2007) menciona que tales determinaciones son erróneas y dichos organismos no pertenecen a Salticidae.
Sin lugar a dudas, los registros más antiguos de Salticidae que se tienen actualmente, son por la presencia de los géneros AlmolinusPetrunkevitch, 1958; GorgopsinaPetrunkevitch, 1955 y ProlinusPetrunkevitch, 1958; considerados extintos; de la subfamilia Hisponinae, para el ámbar del Báltico (Maddison, 2015; Dunlop et al., 2018), con una edad estimada de 44 - 49 Ma (Weitschat y Wichard, 2002 citado en Bodner y Maddison, 2012). Por lo tanto, con base en el registro fósil, se ha sugerido que los salticidos aparecen en la Era Cenozoica (Wunderlich, 2004b; Penney, 2010; Penney y Selden, 2011; Riquelme y Hill, 2013). Por otro lado, los análisis filogenéticos realizados por Maddison y Hedin (2003), Maddison et al. (2014) y un estudio elaborado por Bodner y Maddison (2012), quienes utilizan el método de reloj molecular Bayesiano, datan la edad de Salticidae en 47 - 57 Ma. No obstante, Penney (2010) menciona que no se debería descartar la posibilidad de encontrar a Salticidae en el Mesozoico, en los registros del Cretácico; ya que existe una gran brecha en nuestro conocimiento sobre la presencia de fauna salticida (o la ausencia de ella), en el ámbar de Francia del Eoceno y el ámbar del Cretáceo más joven (53 Ma - 76, 5 Ma) (Penney, 2008). Podría ser solo cuestión de tiempo y paciencia, a fin de que un paleoaracnólogo descubra la primera araña mesozoica saltadora. Si esto fuera así, como lo menciona Bodner y Maddison (2012), el descubrimiento de arañas saltadoras para esa época, podría alterar la edad raíz de Salticidae.
Ahora, sobre la divergencia de Salticidae, en la investigación realizada por Bodner y Maddison (2012) se menciona que los principales clados de la familia están restringidos a regiones muy particulares en los distintos continentes (como ya se mencionó, y ocurre lo mismo en el caso de los lapsinos [incluyendo a Galianora marcoi sp. nov.], quienes están restringidos a los trópicos del Nuevo Mundo [Maddison, 20015]); sugiriendo así que sus radiaciones probablemente se dan después de la separación continental en el Mesozoico. Consecuentemente, con base en las edades estipuladas para la familia (47 - 57 Ma), Bodner y Maddison (2012) mencionan que la diversificación temprana ocurrió en el Eoceno, durante la expansión de los bosques Megatermales y probablemente por el aumento de sus presas. Comenzando esto en el Paleoceno (Paleógeno) (alrededor de 66 Ma), cuando ocurre la expansión de los bosques tropicales debido al calentamiento de la Tierra; persistiendo esto a lo largo del Eoceno (Paleógeno) (Morley, 2000 citado en Bodner y Maddison, 2012).
Sobre este tópico, Hill y Richman (2009) también sugirieron que la divergencia y rápida especiación de las arañas saltadoras probablemente ocurrió entre el Paleoceno (Paleógeno) y el Mioceno (Neogéno). La diversidad de las subfamilias en Salticidae y la presencia de formas relativamente modernas de salticidos en el ámbar depositado en América Central, indica una mayor divergencia dentro de los trópicos desde el Mioceno (Riquelme y Hill, 2013). La riqueza de especies de Salticidae encontradas en el ámbar de República Dominicana y las especies reportadas para el ámbar de Chiapas (ambas del Neogeno), incluyendo a Galianora marcoi sp. nov., es consistente con una subsecuente radiación en el Neogeno, no solo con implicaciones Neárticas, como el caso del género Maevia? (Riquelme y Menéndez-Acuña, 2017), sino también con una distribución más al sur en la región Neotropical, como es el caso de Galianora.
4.5. Evolución geológica: dispersión y/o permanencia de Araneae.
Entonces, se tienen dos géneros en el registro fósil chiapaneco, cuyos representantes actuales están aparentemente restringidos a una región Neártica (Maevia?) y otros más hacia el Sur, en la región Neotropical (Galianora). Indudablemente la historia geológica en América Central es una pieza clave para entender su posible distribución. La evolución tectónica y sedimentaria de lo que conocemos como América Central incluye México, Centroamérica y las Antillas (para una mejor comprensión del tema ver James, 2007); definitivamente esta evolución afectó y modificó no solo el ambiente sino también el clima en estas regiones. La evolución geológica de Chiapas, desde el Oligoceno hasta el Plioceno, también está vinculada con la formación de América Central (Meneses-Rocha, 2001; Mandujano-Velázquez y Keppie, 2009 citado en Riquelme y Hill, 2013). Así que, los fenómenos tectónicos y los materiales sedimentarios que formaron las actuales montañas en Chiapas, durante el Oligoceno-Mioceno, alteraron la distribución de las cuencas y modificaron las barreras de dispersión (Meneses-Rocha, 2001 citado en Riquelme y Hill, 2013); con base en esto, Riquelme y Hill (2013) mencionan que esto provocó cambios en los ecosistemas locales y representó un potencial impulsor para la diversificación, forzando la dispersión de la paleobiota en Chiapas (incluyendo a Salticidae). Así que en la actualidad, géneros de salticidos como Maevia? y Galianora, ocupan regiones norteñas y sureñas en el Nuevo Mundo, respectivamente.
Actualmente, el área donde se encuentran las minas de ámbar en Simojovel presenta un clima cálido subhúmedo, con lluvias en verano y la vegetación es de selva mediana (Inafed, 2017), pero hace 23 Ma era una zona de Manglar, con un clima más tropical (Langenheim, 1995; Serrano-Sánchez et al., 2015 Figura 11). Es importante mencionar que el ámbar de República Dominicana comparte similitudes con el de Chiapas, son aproximadamente contemporáneos y ocurre en escenarios similares (Grimaldi, 1996); sin embargo, el ámbar dominicano fue depositado, probablemente, en un complejo de laguna costera (Iturralde-Vinent y MacPhee, 1996). En estos depósitos ambarinos la araneofauna fósil ha sido ampliamente estudiada (e.g.Ono, 1981; Schawaller, 1984; Reiskind, 1986; Wolff, 1990; Wunderlich, 2004a, 2004b, 2004c, 2004d, 2004e, 2004f, 2004g, 2004h, 2004i, 2004j, 2004k, 2004l; Huber y Wunderlich, 2006; Penney, 1999, 2000, 2001, 2005a, 2005b, 2009; Wunderlich, 2011; Poinar Jr, 2015). Es más, Wunderlich (2004m), Penney (2008) y Penney y Selden (2011) sugirieron que casi todas las familias de arañas fósiles y sus géneros, aún ocupan la misma zona geográfica; para Riquelme y Hill (2013) este tema es debatible.
Nudds y Selden (2008) compararon la biota fósil en el ámbar dominicano con la actual biota de esa región y sugieren que la paleobiota en el ámbar de República Dominicana fue forzada a dispersarse y extinguirse debido a las fluctuaciones climáticas durante la edad de hielo (Neogeno al Pleistoceno). Siguiendo el mismo principio, Riquelme y Hill (2013) mencionan que al parecer la dispersión y extinción de la paleobiota en ámbar de Chiapas se produjo en alopatria por cambios en la tierra y que eso fue seguido por una dispersión forzada a nuevas áreas durante largos periodos de tiempo que abarcaron del Mioceno al Plioceno.
En el contexto anterior, debemos considerar el estudio realizado por Penney y Pérez-Gelabert (2002) sobre la araneofauna fósil y reciente de la Hispaniola (que incluye a República Dominicana y Haití). En ese trabajo se provee un listado de las especies fósiles y actuales de arañas hispaniolicas; las 29 familias encontradas en ámbar, así como 28 de esos géneros están presentes actualmente en esa región; sin embargo, 25 géneros y todas las especies descritas hasta el momento, aparentemente están extintos. Penney y Pérez-Gelabert (2002) mencionaron que las familias presentes en el ámbar dominicano, pero no reportadas para la actual araneofauna hispaniolica, como Cyrtaucheniidae, Ochyroceratidae, Tetrablemmidae, Hersiliidae, Anapidae y Mysmenidae, pueden predecir que tienen representantes actuales en la Hispaniola.
Pérez-Gelabert (2008), elabora un listado, con base en la literatura publicada hasta 2007, de los artrópodos actuales y fósiles de la Hispaniola (República Dominicana y Haiti), incluyendo arañas, él menciona los siguientes registros para la araneofauna actual de esa región, Yabisi guabaRheims y Brecovit, 2004 (Hersillidae), Ochyrocera cachoteHormiga et al., 2007 (Ochyroceratidae) y Microdipoens sp. (Mysmenidae, en Hormiga et al., 2007); de esta manera se corrobora lo antes mencionado por Penney y Pérez-Gelabert (2002). Gabriel de los Santos (com. pers., 2017) menciona que los registros de arañas para la Hispaniola han aumentado considerablemente después de lo publicado por Pérez-Gelabert (2008), pasando ya las 420 especies; pero esta última cifra todavía resulta desactualizada, ya que hace falta incluir nuevas descripciones y realizar una revisión bibliográfica más minuciosa de lo que ya se ha registrado. Probablemente, los nuevos y futuros hallazgos traigan consigo innovadora información sobre el conocimiento de la araneofauna actual y fósil en esa región, que tendrá implicaciones a nivel mundial.
Algo similar ocurre con la araneofauna fósil en Chiapas; en donde 11 géneros, y por lo tanto las especies que los componen, son considerados extintos (ver Dunlop et al., 2018) y seis de ellos tienen representantes en la fauna actual, tal es el caso de Hemirrhagus (Teraphosidae, en García-Villfuerte, 2008), Lyssomanes (Salticidae, en García-Villafuerte y Penney, 2003), Episinus (Theridiidae, en García-Villafuerte, 2006a) y Selenops (Selenopidae, en García-Villafuerte, 2006b). Así que, siguiendo el pensamiento de Penney y Pérez-Gelabert (2002), no se puede predecir que las familias araneidas (y sus componentes), que se encuentran en la actual araneofauna chiapaneca, vayan a ser descubiertas en el ámbar mexicano proveniente de Simojovel (aunque existe una gran probabilidad, dada la evidencia que se tiene por los hallazgos ya realizados), pero con base en el registro de arañas fósiles y actuales de Chiapas (García-Villafuerte, 2009), existe la posibilidad de que los géneros de las diferentes familias de arañas descubiertos en el ámbar mexicano, aun formen parte de la actual araneofauna en Chiapas.
Por lo tanto, con base en las investigaciones realizadas por Penney y Pérez-Gelabert (2002), Hormiga et al. (2007) y Pérez-Gelabert (2008), no solo se debe considerar la evolución geológica y los factores ambientales ya mencionados, para sugerir que la biota fósil dominicana y chiapaneca, y muy en particular la araneofauna incluida en el ámbar de ambos sitios, se ha extinguido y se vio forzada a una dispersión hacia otras áreas, ya que las arañas (así como sus presas, los insectos), han mostrado un fuerte carácter adaptativo a las condiciones cambiantes (Decae, 1984; Dimitrov et al., 2011), y esto es confirmado por la permanencia de representantes no fósiles en la actual araneofauna de la Hispaniola (Pérez-Gelabert, 2008), los cuales fueron primeramente reportados como registros fósiles (e.g. Hersillidae y Mysmenidae).
5. Conclusiones
Probablemente muchas de las especies de arañas en ámbar (sino es que todas) que se han encontrado en Chiapas, se extinguieron; pero existe una gran probabilidad, con base en los descubrimientos hechos, que otras especies, representando los mismos géneros y/o familias, hayan ocupado su lugar; pero para fortalecer esta hipótesis, aceptarla o rechazarla, se propone que se incrementen los estudios araneofaunísticos y se adhiera la información a los trabajos previamente realizados (e.g.Ibarra-Núñez et al., 1997; Medina-Soriano, 2005; Santos, 2005; Ibarra-Núñez et al., 2011), que se han centrado en la riqueza y diversidad de las especies de arañas actuales en Chiapas, así como la consideración de estudios filogenéticos con la inclusión de taxa fósiles en ambas áreas. Entonces, si representantes de los géneros Mysmena, Maevia? y Galianora, por mencionar algunos, son encontradas en Chiapas, implicaría que la evolución geológica y los cambios ambientales, por más drásticos que hayan sido, no perturbaron la permanencia de estos géneros en esta región del mundo y que es probable, con base en el registro fósil, que Chiapas fue, en su momento, un centro de origen para muchas especies de arañas, que en la actualidad ocupan diferentes hábitats. Futuras investigaciones relacionadas con arañas fósiles y actuales, podrían confirmar o rechazar esto.

