











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink1. Introducción
En México, el registro paleoicnológico de vertebrados terrestres es relativamente abundante y diverso; la mayor parte de la información generada al respecto corresponde a huellas del Mesozoico que fueron producidas por dinosaurios, reptiles voladores, cocodrilomorfos, tortugas y aves (Rodríguez-de la Rosa et al., 2004). Por otro lado, los registros del Cenozoico de icnitas son más bien escasos: una localidad en Sinaloa, una en Durango, dos en Jalisco, una en Coahuila y cuatro en Puebla. En estos registros se han reportado huellas de aves y diversos mamíferos (Rodríguez-de la Rosa et al., 2004; Lockley y Delgado, 2007; Lockley y Bishop, 2014).
Sin duda, la localidad cenozoica con el mayor número de icnitas referidas para mamíferos que se conoce hasta el momento en México es Pie de Vaca, en el municipio de Tepexi de Rodríguez, estado de Puebla, la cual es conocida por los moradores del área desde hace más de 100 años. En esta localidad se pueden reconocer nueve rastros de camélidos, tres rastros de félidos, un rastro parcial de un artiodáctilo pequeño, huellas aisladas de aves y el tafoglifo (impresión del cadáver de un animal sensu Sarjeant, 1975: 284) de un flamenco (Lockley, 1991; Cabral-Perdomo, 1995, 2000, 2013). Las impresiones de los camélidos y félidos son las más representativas y abundantes en el sitio.
Si bien es cierto que el registro icnológico de Pie de Vaca se ha mencionado en otros trabajos (Cabral-Perdomo, 1995, 2000, 2013; Rodríguez-de la Rosa et al., 2004; Beraldi-Campesi et al., 2006), no se ha realizado su análisis icnológico. El presente trabajo tiene como propósito establecer la caracterización icnotaxonómica formal de las huellas de camélidos y félidos de la localidad Pie de Vaca, así como establecer algunas consideraciones paleobiológicas relacionadas con el tamaño, modo de progresión, velocidad y comportamiento de los individuos que las generaron.
2. Área de estudio
La localidad Pie de Vaca se localiza 3 km al noreste del centro de Tepexi de Rodríguez, Puebla, entre las coordenadas 18o35’24’’N y 97o55’08’’W (Figura 1). Esta unidad se denomina acríticamente formación Pie de Vaca, la cual se asocia a depósitos de origen lacustre (Beraldi-Campesi et al., 2006). En el Léxico Estratigráfico de México del Servicio Geológico Mexicano, la denominación de Pie de Vaca como unidad estratigráfica es informal (Juárez-Arriaga, 2008), dado que no cumple con los artículos necesarios propuestos por el Código de Nomenclatura Estratigráfico de Norteamérica para recibir el nombre de una unidad litoestratigráfica formal (Artículos 3 - 16 en North American Commission on Stratigraphic Nomenclature, 2005). Con base en esto, en el presente estudio, el nombre Pie de Vaca se usa de manera informal para referirse a la secuencia portadora de las impresiones.
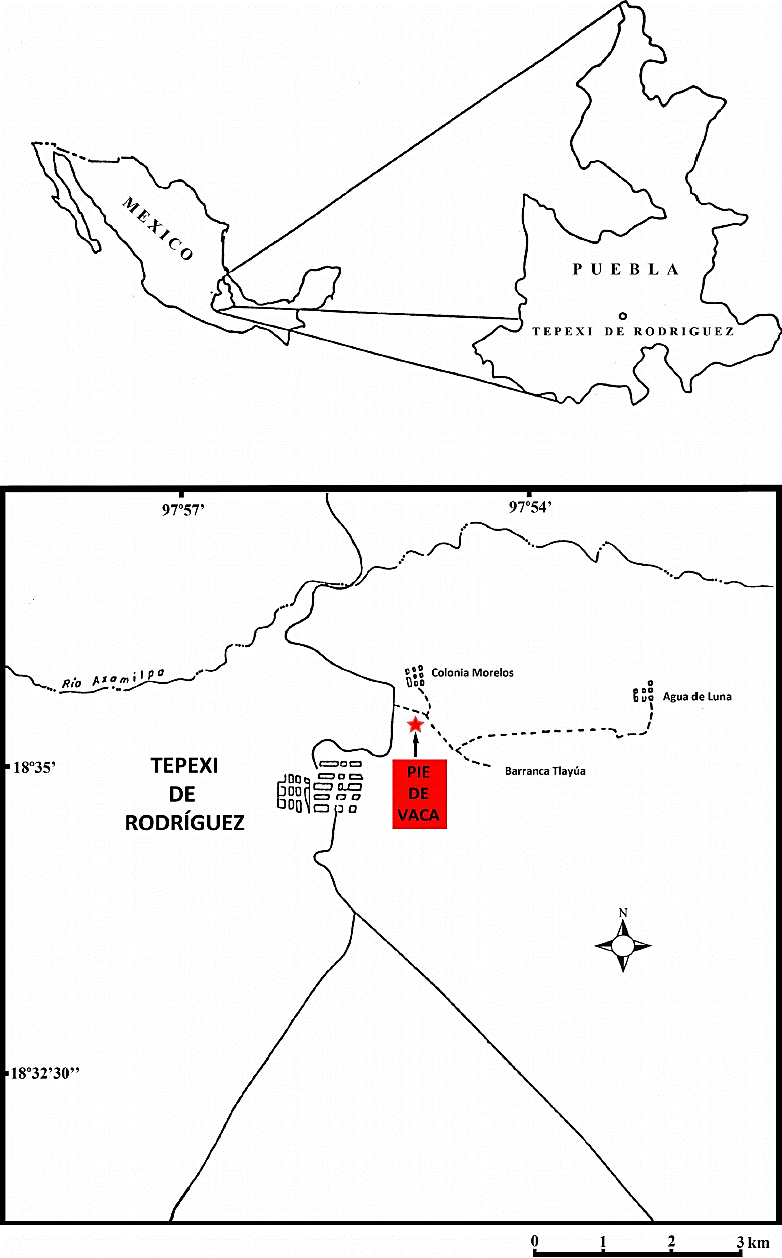
Figure 1 Mapa índice que muestra la localización de la localidad Pie de Vaca en la porción centro-meridional del estado de Puebla, centro de México.
En términos generales, la formación Pie de Vaca está compuesta por una sucesión de caliza micrítica de color que varía de anaranjado amarillento pálido (10YR 8/6) a anaranjado grisáceo (10YR 7/4) (Rock-Color Chart Committee, 2009). Tiene una estratificación delgada a mediana (1 - 30 cm), bandas de sílice e intraclastos de caliza, pedernal y roca volcánica, las cuales descansan en discordancia erosional sobre la Formación Tlayúa. La secuencia continúa con una serie alternante de grava, arena, limo y arcilla, seguidos de un conglomerado polimíctico bien compactado con fragmentos de caliza, pedernal, cuarzo y rocas volcánicas de composición variable (Pantoja-Alor et al., 1989) (Figura 2). La edad de esta unidad es controversial debido a que el registro de huellas de mamíferos sugiere una edad del Neógeno - Pleistoceno (Applegate et al., 1984; Cabral-Perdomo, 1995), mientras que evidencia palinológica recuperada de localidades cercanas a Pie de Vaca sugiere una edad más antigua del Paleógeno (Eoceno - Oligoceno) (Martínez-Hernández y Ramírez-Arriaga, 1999).
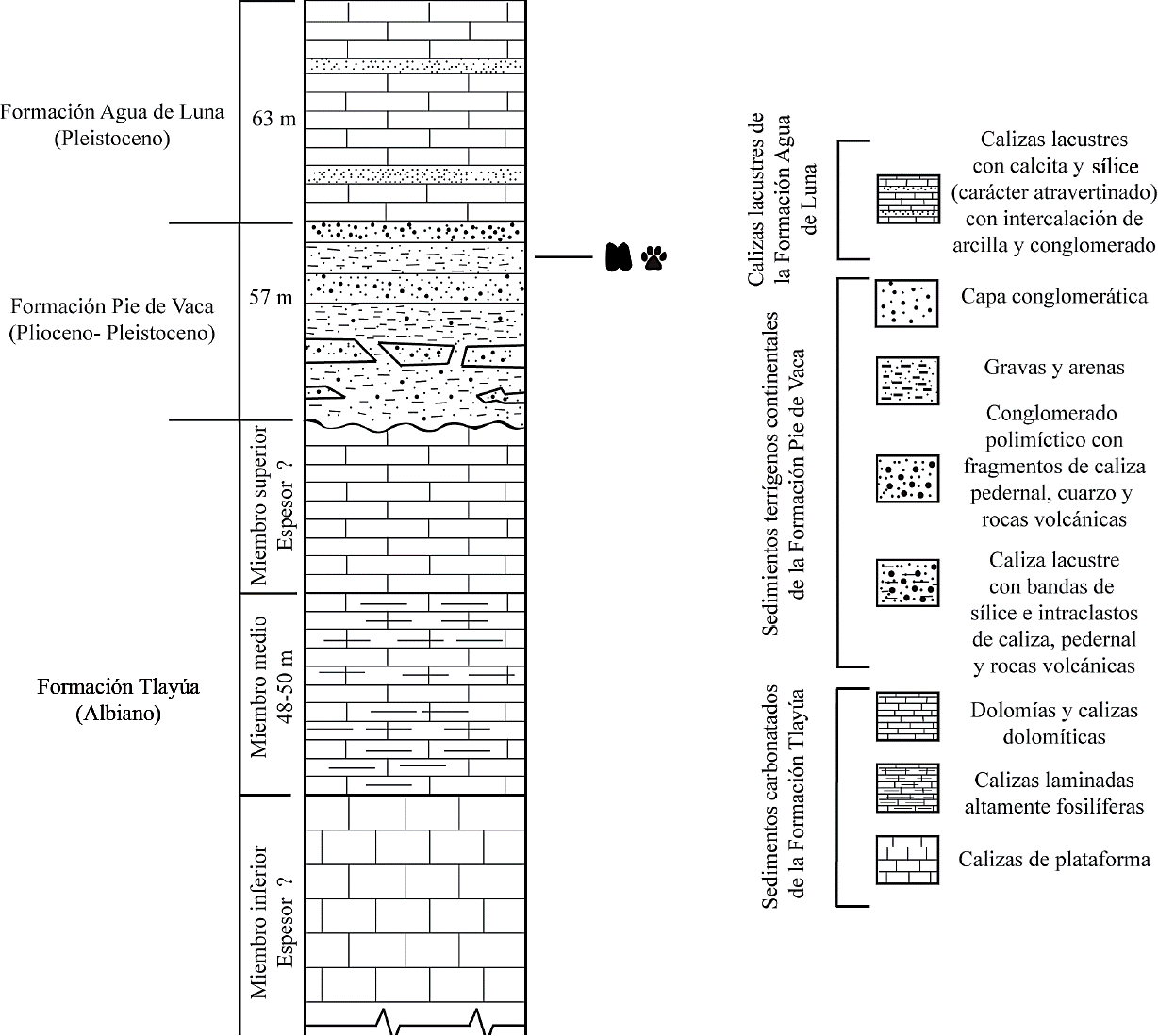
Figura 2 Columna estratigráfica del área de Tepexi de Rodríguez, Puebla (modificado de Cabral-Perdomo, 1995). Se indica el nivel portador de las huellas de camélidos y félidos de la localidad Pie de Vaca.
Precisar la edad de la unidad portadora está fuera del alcance del presente estudio; sin embargo, considerarla como parte del Paleógeno es poco probable si se toman en cuenta los siguientes hechos: (1) el tamaño de los camélidos que produjeron los rastros de Pie de Vaca no es consistente con el de animales del Paleógeno, dado que durante ese periodo tuvieron un tamaño semejante al de una gacela; (2) el félido norteamericano más antiguo corresponde a Pseudaelurus, un inmigrante euroasiático del Mioceno (Prothero, 2006); (3) huellas pertenecientes a proboscídeos también se han reportado en estratos referibles a la formación Pie de Vaca, en la localidad Pochote - Elefante (Cabral-Perdomo, 1995) y se sabe que los primeros representantes de ese grupo llegaron a Norteamérica durante el Mioceno (Prothero, 2006). Aunado a esto, la configuración de las huellas y el tamaño relativamente grande de los organismos que produjeron el registro icnológico de Pie de Vaca es consistente con lo que se observa en los camélidos y félidos del Plioceno - Pleistoceno de Norteamérica (Jiménez-Hidalgo et al., 2015). En este trabajo se considera que la edad más probable de la unidad portadora corresponde con este segmento geocronológico, como previamente algunos autores han indicado (Cabral-Perdomo, 1995, 2013; Jiménez-Hidalgo et al., 2015). El afloramiento icnofosilífero en el que se encuentran las huellas y rastros de camélidos (en lo sucesivo RC) y de félidos (en lo sucesivo RF) abarca una extensión de alrededor de 190 m2 (Figura 3A). En el mismo estrato portador de los fósiles se encontraba el tafoglifo de un flamenco, el cual fue recolectado debido a que fue objeto de vandalismo y trasladado al Museo de Geología de la Universidad Nacional Autónoma de México.
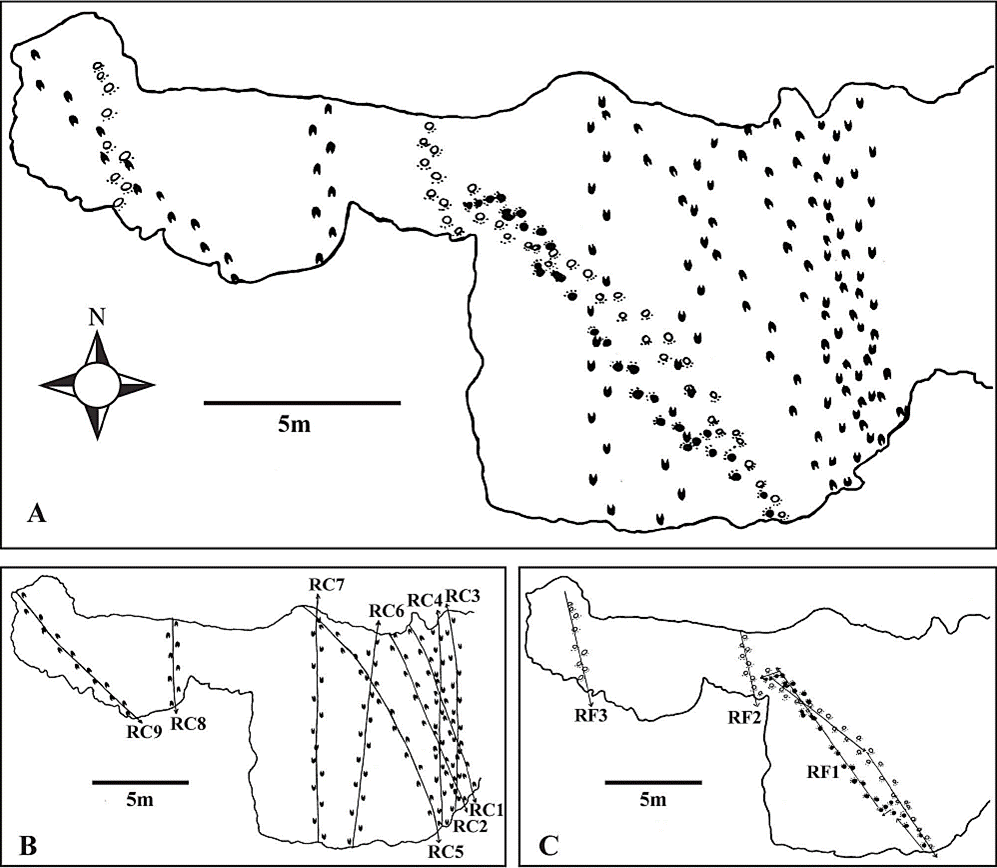
Figura 3 Distribución espacial de (A) las huellas de camélidos y félidos de Pie de Vaca, Cenozoico tardío de Puebla, centro de México. Se incluye la dirección de progresión de los individuos que generaron (B) los rastros de los camélidos (RC) y (C) los rastros de los félidos (RF). Barra de escala = 5 metros.
3. Materiales y Métodos
La muestra considerada en este estudio incluye un total de 233 huellas: 154 de camélidos organizadas en nueve rastros y 79 de félidos organizadas en tres rastros.
Las huellas y los rastros se midieron con una cinta métrica flexible directamente en el afloramiento durante los meses de mayo y junio de 1994, como parte del proyecto de tesis de licenciatura del primer autor (Cabral-Perdomo, 1995). La estadística univariada (promedio, desviación estándar y rango observado) de las medidas se estimó usando el programa de estadística PAST para MacOS (Hammer et al., 2001). Las medidas están expresadas en cm.
3.1. Dimensiones estimadas
En cada una de las huellas atribuidas a camélidos, se estimaron las siguientes medidas (Figura 4A): longitud y ancho internos de la huella; longitud y ancho de cada dígito; y ángulo interdigital. Por otro lado, en cada una de las huellas atribuidas a félidos se estimó el ancho y la longitud máximos (Figura 4B).
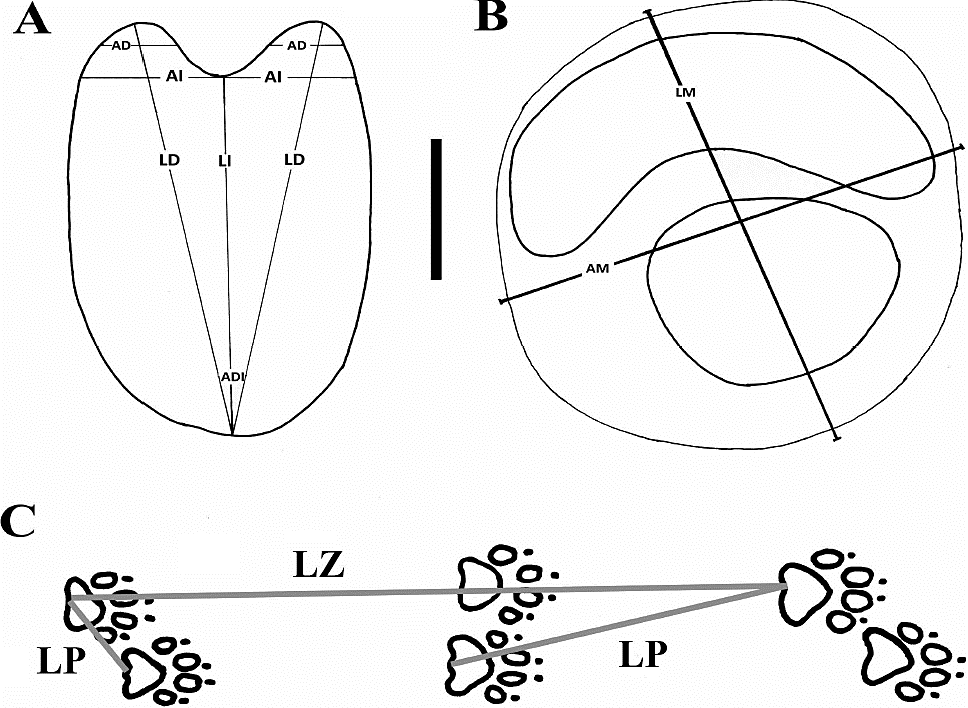
Figura 4 Esquemas de huellas de (A) camélido y (B) félido de Pie de Vaca, en los que se indican las dimensiones estimadas, así como un diagrama de un rastro de Canis familiaris (C) en el que se indican la longitud del paso (LP) y la longitud de la zancada (LZ). Abreviaturas: AI, Ancho interno de la huella; ADI, Ángulo Interdigital; AD, Ancho de los dígitos III y IV; AM, Ancho máximo de la huella; LI, Longitud interna de la huella; LD, Longitud de los dígitos III y IV; LM, Longitud máxima de la huella. Barra de escala = 5 cm.
En todos los rastros se estimó la longitud del paso y la zancada (Figura 4C), de acuerdo con lo propuesto por Sarjeant (1975, 1989).
3.2. Identidad de los productores de las icnitas
Con la finalidad de establecer la identidad de los productores potenciales de las huellas, se llevó a cabo una comparación del tamaño y morfología de las icnitas estudiadas con huellas producidas tanto por taxones extintos como por actuales, previamente reportados en la literatura (Aramayo y de Bianco, 1987; Remeika, 2001; Lucas y Hunt, 2007; Rodríguez-de la Rosa et al., 2007; Pérez-Lorente et al., 2009; Diedrich, 2011; Aramayo et al., 2015).
3.3. Estimación del tamaño y la velocidad
La estimación del tamaño de los productores de las huellas se fundamenta en la correlación positiva entre la longitud de una huella con la altura de las extremidades de un mamífero bípedo o cuadrúpedo en reposo (Alexander, 1976; Thulborn, 1990; Henderson, 2003). Para tal efecto, se utilizó el método morfométrico de Alexander (1976) modificado por Thulborn (1990). En el presente estudio se describe con base en la siguiente ecuación:
AE corresponde a la altura promedio de las extremidades (anteriores y posteriores), L corresponde a la longitud promedio de las huellas y X se refiere a la proporción AE/L.
En el caso de los camélidos, la proporción AE/L se estableció tomando como referencia el promedio de la altura de las extremidades anteriores y posteriores (AE = 97.9 cm) (Bonacic y Franklin, 2006; Tabla 1 en Van der Sluijs et al., 2010: 292), así como la longitud promedio de huellas (L = 10.8 cm) (Tabla 3 en Remeika, 2001: 70) pertenecientes a la llama Lama glama. Para los félidos se llevó a cabo el mismo procedimiento, usando como referencia el promedio de la altura de las extremidades anteriores y posteriores (AE = 120 cm) (Packer, 2006), así como la longitud promedio de huellas (L = 15.81 cm) (fig. 75.3c en Liebenberg, 1990: 179) pertenecientes al león africano Panthera leo. Con base en los datos disponibles, la estimación de la altura promedio de las extremidades a partir de la longitud de la huella se determinó mediante las fórmulas AE = 9L y AE = 7.6L para camélidos y félidos respectivamente.
La velocidad de los productores de las huellas se estimó con base en la ecuación de Alexander (1976, 2006) como sigue:
Donde V corresponde a la velocidad, g es la aceleración de la gravedad (9.8 m/s2) y LZ corresponde a la longitud promedio de la zancada. La velocidad estimada se expresa en m/s.
El modo de progresión de los camélidos y félidos de Pie de Vaca, se estableció mediante la proporción entre la longitud promedio de la zancada y la altura promedio de las extremidades (LZ/AE), dado que en términos generales, se sabe que un mamífero terrestre cambia de caminar a trotar/ correr cuando esta proporción alcanza un valor de aproximadamente 2.0 (Alexander, 1976). Con base en Thulborn (1982) y Thulborn y Wade (1984) se consideraron los siguientes modos de progresión: caminar (LZ/AE < 2.0), trotar (2.0 < LZ/AE < 2.9) y correr (LZ/AE > 2.9).
La dirección de progresión de los individuos que generaron los rastros se determinó con base en la posición del norte magnético del Plioceno-Pleistoceno, la cual es prácticamente indistinguible de la actual (García-Ruiz et al., 2016).
4. Icnología Sistemática
División Vertebratichnia Vialov, 1966
Clase Mammalipedia Vialov, 1966
Orden Artiodactipedia Vialov, 1966
Morfofamilia Pecoripedia Remeika et al., 1995
Icnogénero Lamaichnum Aramayo y de Bianco, 1987
Lamaichnum guanicoe Aramayo y de Bianco, 1987
Material referido. 154 huellas organizadas en 9 rastros, mismas que se encuentran en la localidad Pie de Vaca.
Descripción. Huellas conservadas en epirelieve cóncavo, subdigitígradas, de forma subovalada, longitud promedio de 13.2 cm y ancho promedio de 10.5 cm, con un ángulo interdigital promedio de 29.6o (Tabla 1). Las huellas muestran una morfología homogénea, lo que dificulta diferenciar las huellas producidas por las extremidades anteriores de las producidas por las posteriores (Homopodia). En términos generales, se distinguen por presentar impresiones dactilares subiguales en tamaño; dos marcas paralelas de uñas en punta separadas en la parte anterior; la región posterior de estas dos marcas confluye medialmente en una forma acorazonada y no se aprecia un surco medial (= septo interdigital); los bordes laterales son más o menos rectos y paralelos, mientras que el borde posterior es ligeramente convexo (Figura 5A).
Tabla 1 Promedio, desviación estándar y rango observado (entre paréntesis) de los parámetros evaluados en las huellas de camélidos de Pie de Vaca, Cenozoico tardío de Puebla, centro de México. Abreviaturas: LI, longitud interna de la huella; AI, ancho interno de la huella; LDIII, longitud del dígito III; LDIV, longitud del dígito IV; ADIII, ancho del dígito III; ADIV, ancho del dígito IV; ADI, ángulo interdigital; LP, longitud de paso; LZ, longitud de la zancada; N, número de huellas. Las medidas están en cm a excepción del ángulo interdigital.
| Rastro | N | LI | AI | LDIII | LDIV | ADIII | ADIV | ADI | LP | LZ |
|---|---|---|---|---|---|---|---|---|---|---|
| RCl | l7 | 13.14 ± 1.15 (10.5 - 15.5) | 10.58 ± 0.75 (9.5 - 12) | 14.93 ± 0.77 (13.5 - 16) | 15.15 ± 0.96 (13 - 17) | 4.63 ± 0.78 (2.5 - 6) | 5.09 ± 0.61 (4 - 6) | 29.6° ± 3.4 (20° - 35°) | 68 ± 2.98 (60 - 71) | 253.15 ± 6.86 (240 - 261) |
| RC2 | l6 | 13.21 ± 1.21 (11 - 15.5) | 10.31 ± 0.35 (10 - 11) | 14.50 ± 1.06 (12 - 16) | 14.26 ± 0.77 (12.5 - 15.5) | 4.63 ± 0.39 (4 - 5) | 4.70 ± 0.56 (3.5 - 5.5) | 29° ± 3.87 (25° - 35°) | 69.3 ± 3.08 (63 - 74) | 259.91 ±3.14 (254 - 265) |
| RC3 | 18 | 12.47 ± 0.67 (11 - 13.5) | 10.11 ± 0.63 (9.5 - 11.5) | 15.17 ± 0.86 (13.5 - 16.5) | 14.46 ± 0.82 (13 - 15.5) | 4.64 ± 0.38 (4 - 5) | 4.65 ± 0.35 (4 - 5) | 29.1° ± 3.3 (25° - 35°) | 68.26 ± 1.71 (65 - 71) | 253.5 ± 10.94 (230 - 268) |
| RC4 | 19 | 13.1 ± 0.47 (12.5 - 14) | 10.43 ± 0.49 (9.5 - 11.5) | 14.83 ± 0.72 (13.5 - 16) | 15.03 ± 0.35 (14.5 - 15.5) | 4.76 ± 0.41 (4 - 5.5) | 4.76 ± 0.41 (4 - 5.5) | 28.6° ± 3.5 (25° - 35°) | 68.36 ± 2.30 (63 - 71) | 249.46 ± 8.7 (231 - 264) |
| RC5 | 23 | 13.5 ± 0.58 (12.5 - 14.5) | 10.13 ± 0.45 (9.5 - 11) | 14.58 ± 0.59 (13.5 - 16) | 14.60 ± 0.45 (14 - 15.5) | 4.0 ± 0.45 (3 - 4.5) | 4.0 ± 0.39 (3.5 - 4.5) | 29.6° ± 3.3 (25° - 35°) | 68.7 ± 2.07 (64 - 72) | 251.84 ± 5.02 (243 - 263) |
| RC6 | l7 | 13.67 ± 0.76 (12.5 - 15.5) | 10.08 ± 0.64 (9.5 - 11.5) | 14.26 ± 0.50 (13.5 - 15) | 14.20 ± 0.39 (13.5- 15) | 4.38 ± 0.37 (4 - 5) | 4.41 ± 0.36 (4 - 5) | 29.7° ± 3.3 (25° - 35°) | 69.62 ± 1.57 (65 - 72) | 259.61 ± 4.57 (251 - 266) |
| RC7 | 18 | 13.42 ± 0.38 (13 - 14) | 10.39 ± 0.34 (10 - 11) | 14.53 ± 0.53 (14 - 15.5) | 14.60 ± 0.28 (14 - 15) | 4.32 ± 0.50 (3.5 - 5) | 4.32 ± 0.24 (4.0 - 4.5) | 30° ± 2.7 (25° - 35°) | 70.20 ± 1.57 (67 - 72) | 266.6 ± 3.98 (260 - 271) |
| RC8 | 9 | 13.16 ± 0.35 (12.5 - 13.5) | 10.4 ± 0.39 (10 - 11) | 14.38 ± 0.33 (14 - 15) | 14.4 ± 0.39 (14 - 15) | 4.2 ± 0.44 (3.5 - 5) | 4.16 ± 0.35 (3.5 - 4.5) | 28.3° ± 2.5 (25° - 30°) | 68.8 ± 2.70 (64 - 72) | 248 ± 3.67 (243 - 252) |
| RC9 | 17 | 13.29 ± 0.50 (12.5 - 14) | 10.29 ± 0.39 (9.5 - 11) | 14.38 ± 0.37 (13.5 - 15) | 14.26 ± 0.31 (13.5 - 14.5) | 4.0 ± 0.43 (3.5 - 5) | 3.94 ± 0.34 (3.5 - 4.5) | 28.8° ± 3.3 (25° - 35°) | 68.59 ± 2.16 (64 - 71) | 248.30 ± 8.46 (229- 258) |
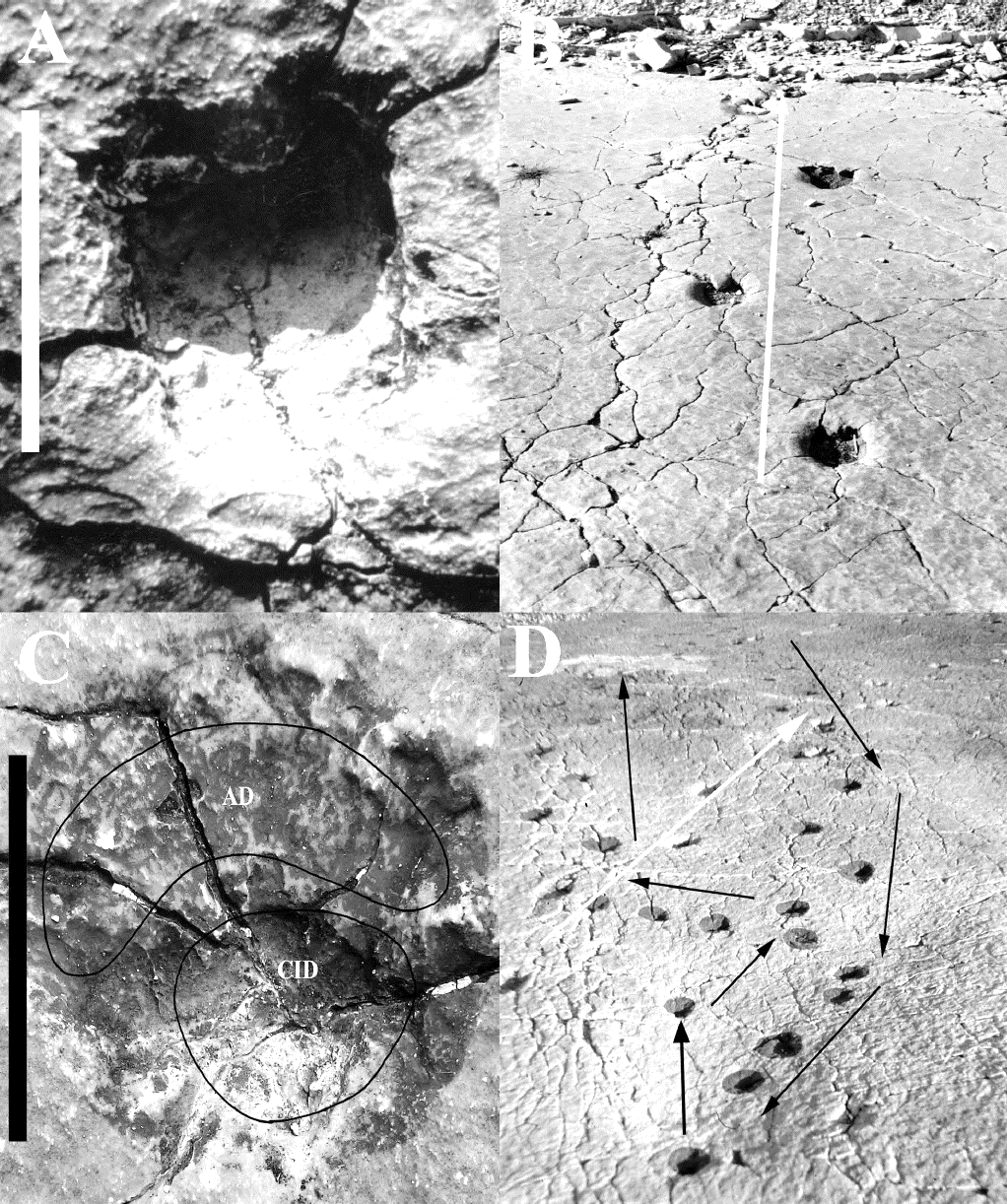
Figura 5 Huellas y rastros de Pie de Vaca: (A) Huella de camélido; (B) Secuencia de cuatro huellas del rastro 3 de camélido con la dirección de progresión indicada con la flecha blanca; (C) Huella de felino; (D) Vista parcial del afloramiento hacia el norte en donde se aprecia la progresión del felino del rastro 1 (flechas negras) y la progresión del camélido del rastro 6 (flecha blanca). Abreviaturas: AD, Arcada Digital; CID, Cojinete Interdigital. Barra de escala = 10 cm.
Para cada uno de los nueve rastros, la longitud de paso promedio es de 69 cm y la longitud de zancada promedio es de 253 cm (Figuras 3B y 5B, Tabla 1).
Discusión. Las ligeras diferencias en los bordes anterior y posterior de las huellas, así como la ausencia del surco interdigital, representan variación extramorfológica asociada a la interacción de pies-manos en un sustrato relativamente suave y con cierta humedad, así como alteración postdiagenética por intemperismo y erosión. La variación extramorfológica observada en huellas de camélidos es amplia y común (Lucas y Hunt, 2007).
De acuerdo a Lucas y Hunt (2007), a partir del Mioceno y hasta el Pleistoceno, los camélidos adquirieron una estructura podial estereotípica, lo que dificulta distinguir fehacientemente las huellas dejadas por diferentes géneros o especies de ese grupo de artiodáctilos. Estos autores proponen que las huellas de camélidos que abarcan dicho lapso sean asignadas únicamente al icnogénero Lamaichnum. Para estos autores, este icnotaxón está integrado por las icnoespecies L. guanicoe y L. macropodum, las cuales se diferencian prácticamente por el tamaño. Las huellas con una longitud de 4 a 15 cm se asignan a L. guanicoe y aquellas con una longitud de 16 a 20 cm se atribuyen a L. macropodum (Lucas y Hunt, 2007). Recientemente, (Aramayo et al., 2015) se ha propuesto una nueva icnoespecie dentro del icnogénero: L. tulipensis, misma que se caracteriza por tener heteropodia, por tener las impresiones de los dedos de las manos dirigidos hacia afuera y por tener dimensiones mayores a las de L. guanicoe. Sin embargo, Oliva y Arregui (2018) consideran que la morfología de estas huellas se debe a patologías en los individuos productores, por lo que sinonimizan a L. tulipensis con L. guanicoe. Las huellas de Pie de Vaca tienen una longitud promedio de 13 cm, la cual está en el límite superior de las L. guanicoe y, por lo tanto, se asignan a esta icnoespecie.
La mayoría de las icnitas referidas para el país son de dimensiones que se encuentran en el rango de Lamaichnum guanicoe. De esta forma, en la localidad Las Pintas en el estado de Durango, Lockley y Delgado (2007) describen un rastro en cuyas huellas estimaron longitudes de 5 a 8 cm y ancho de 5 a 6 cm. Los autores confirieron estos icnofósiles al icnogénero Lamaichnum sin pretender identificarlas a nivel específico.
En otro reporte de índole meramente geológico (Dávalos-Álvarez et al., 2007), se menciona la presencia de icnofósiles de camélidos en la unidad Sedimentos Lacustres Villa Alegría de la Formación Tehuacán en el estado de Puebla, pero no se describen formalmente. En la ilustración que presentan (fig. B 1 en Dávalos-Álvarez et al., 2007: 212) se puede apreciar que la morfología y dimensiones de las huellas de este sitio son comparables a las de Tepexi de Rodríguez, por lo que los autores consideran que la edad de la unidad portadora de las huellas de Tehuacán sería semejante a la de la localidad Pie de Vaca.
Un último reporte de icnofósiles de camélidos para el país es el publicado por Rodríguez-de la Rosa y Guzmán-Gutiérrez (2012), quienes reportaron huellas de Lamaichnum macropodum en la localidad de San Juan de Los Lagos en Jalisco. Las icnitas tienen en promedio 16 cm de longitud y 11.7 cm de ancho, dimensiones que caen dentro del rango propuesto por Lucas y Hunt (2007) para dicha icnoespecie.
Orden Carnivoripedida Vialov, 1966
Morfofamilia Felipedidae Remeika, 2001
Material reportado. 79 icnitas repartidas en tres rastros que continúan en la localidad Pie de Vaca.
Descripción. Huellas conservadas en epirelieve cóncavo, digitígradas, subredondeadas, longitud promedio de 13.5 cm y ancho promedio de 12.8 cm (Tabla 2). La forma general de las huellas es muy semejante (Homopodia) y, por lo tanto, es difícil distinguir entre impresiones manuales y podiales. Las huellas tienen una profundidad que varía de 1 a 3 cm. En la parte posterior presentan un cojinete interdigital de forma subtriangular a semiredonda que en ocasiones es muy profundo, mientras que en la parte anterior se observa una depresión reniforme continua menos marcada que el cojinete interdigital y que corresponde al área en la que se dispone la arcada de los cojinetes dactilares. El borde anterior de dicha depresión es convexo y no se aprecian impresiones de garras o uñas (Figura 5C). En este conjunto se incluyen 79 huellas organizadas en tres rastros. La longitud promedio del paso es de 44 cm y la longitud promedio de la zancada es de 171 cm (Figuras 3C y 5C-D, Tabla 2).
Tabla 2 Promedio, desviación estándar y rango observado (entre paréntesis) de los parámetros evaluados en las huellas de félidos de Pie de Vaca, Cenozoico tardío de Puebla, centro de México. Abreviaturas: LM, longitud máxima de la huella; AM, ancho máximo de la huella; LP, longitud de paso; LZ, longitud de la zancada; N, número de huellas. Las medidas están en cm.
| Rastro | N | LM | AM | LP | LZ |
|---|---|---|---|---|---|
| RF1 | 17 | 12.71 ± 1.79 (8 - 20) |
12.54 ± 1.63 (8 - 16) |
50.31 ± 9.02 (35 - 60) |
191.36 ± 3.06 (187 - 197) |
| RF2 | 16 | 13.44 ± 1.26 (11 - 15) |
12.33 ± 1.47 (10 - 15) |
39.75 ± 1.70 (38 - 42) |
154.4 ± 7.33 (145 - 163) |
| RF3 | 18 | 14.44 ± 1.33 (13 - 17) |
13.5 ± 1.95 (10 - 16) |
42 ± 2.64 (40 - 45) |
167 ± 7.07 (162 - 172) |
Discusión. La configuración de las huellas de félidos de Pie de Vaca es semejante a las impresiones incluidas en la morfofamilia Felipedidae, por su condición digitígrada, la configuración de la arcada de los dígitos, así como la forma y tamaño del cojinete interdigital (Remeika, 2001). Las huellas de félidos de Pie de Vaca tienen el doble de tamaño que las consideradas en Felipeda (longitud ≤ 5 cm) y en Pumaeichnum (longitud ≈ 6 cm), son un 40% más grandes que las de Pycnodactylopus (longitud ≈ 8 cm) y un 25% más grandes que las de Mitsupes (longitud ≈ 10 cm). Proponer un nuevo icnotaxón con base en el tamaño grande de las huellas de Pie de Vaca es poco pertinente. Aunado a esto, su pobre detalle en otros rasgos diagnósticos, tales como el tamaño, configuración y disposición de cada uno de los dígitos, limitan una evaluación y comparación completas de su patrón morfotípico. Por lo tanto, se prefirió identificarlas icnotaxonómicamente a nivel de la morfofamilia. En lo que respecta a la diversidad icnotaxonómica de félidos en Norteamérica, Sarjeant et al. (2002) reconocieron a los icnogéneros Pycnodactylopus y Felipeda en la morfofamilia Felipedidae, mientras que Remeika (2001, 2006) considera únicamente al icnogénero Pumaeichnum con tres icnoespecies (P. biancoi, P. milleri y P. stouti) del Plioceno temprano de California. Recientemente, Rodríguez-de la Rosa y Guzmán-Gutiérrez (2012), erigieron a la icnoespecie Mitsupes dugesi para identificar a un conjunto de huellas atribuidas a macairodontinos del Neógeno tardío de Jalisco, centro-norte de México. En el Mioceno de España y Rumania se describieron huellas de félidos referidas al icnogénero Felipeda con dos icnoespecies: F. lynxi y F. parvula (Anton et al., 2004).
5. Consideraciones paleobiológicas del registro
5.1. Afinidad del productor de las huellas de camélidos
Las huellas de camélidos de Pie de Vaca son semejantes a las impresiones que dejan estos artiodáctilos en los siguientes rasgos: (1) digitígradas, (2) huellas de configuración ovoidal, (3) impresiones manuales y podiales con una morfología semejante, (4) ápices de los dígitos dirigidos anteriormente, así como (5) borde lateral de los dígitos convexo y borde medial recto (Aramayo y de Bianco, 1987; Sarjeant y Langston, 1994; Remeika, 2001; Lucas y Hunt, 2007).
El tamaño promedio de las huellas de Pie de Vaca es un 15% mayor al de huellas atribuidas a Hemiauchenia sp. del Plioceno de California y aproximadamente un 20% más grande al de las llamas actuales de la especie Lama glama (Remeika, 2001, 2006). Entre los camélidos más comunes del Cenozoico tardío de Norteamérica con un tamaño algo más grande que las llamas y guanacos actuales está Hemiauchenia (Hulbert y Webb, 2001; Lange, 2002). Por lo anterior, es probable que los productores de las huellas de camélidos de Pie de Vaca hayan sido integrantes de ese género.
5.1.1. Tamaño, velocidad y modo de progresión
La altura promedio de las extremidades de los camélidos de Pie de Vaca es relativamente homogénea, aproximadamente de 1.2 m. El individuo que produjo el rastro 3 correspondería al más pequeño (1.12 m) y el más grande al del rastro 6 (1.23 m) (Tabla 3). Los rastros 1, 2, 5, 8 y 9 son paralelos, se dirigen hacia el sureste y están espaciados por una distancia que varía de 2.5 m a 12 m. Por su parte, los rastros 3, 4, 6 y 7 son paralelos, se dirigen al norte-noreste y están separados por una distancia promedio de 3 m (Figura 3B, Tabla 3). La velocidad promedio de los productores de los rastros es de 3.04 m/s. La proporción LZ/AE es en promedio de 2.13 y se asociaría a un cambio en el modo de progresión de caminar a trotar (Alexander, 1976; Thulborn, 1982) (Tabla 3).
Tabla 3 Estimación de la altura promedio de las extremidades (AE), dirección de progresión (Dp), velocidad (V) y modo de progresión de los camélidos y félidos de Pie de Vaca, Cenozoico tardío de Puebla, centro de México. N, número de huellas. La proporción de la longitud de la zancada y altura de la extremidad (LZ/AE) se asoció a los siguientes modos de progresión: LZ/AE < 2.0 = caminar, 2.0 < LZ/AE < 2.9 = trotar, LZ/AE > 2.9 = correr.
| Rastro | N | AE | Dp | V (m/s) | LZ/AE | Progresión* |
| Camélidos | ||||||
| RC1 | 17 | 118.32 | 22° SE | 3.03 | 2.13 | trote |
| RC2 | 16 | 118.96 | 24° SE | 3.15 | 2.18 | trote |
| RC3 | 18 | 112.25 | N | 3.22 | 2.25 | trote |
| RC4 | 19 | 117.9 | N | 2.98 | 2.12 | trote |
| RC5 | 23 | 121.5 | 31° SE | 2.91 | 2.07 | caminar-trote |
| RC6 | 17 | 123.08 | 07° NE | 3.01 | 2.11 | trote |
| RC7 | 18 | 120.85 | N | 3.23 | 2.2 | trote |
| RC8 | 9 | 118.5 | 02° SE | 2.93 | 2.09 | caminar-trote |
| RC9 | 17 | 119.64 | 43° SE | 2.91 | 2.07 | caminar-trote |
| Félidos | ||||||
| RF1 | 61 | 96.6 | 36° NO - 43° SE | 2.41 | 1.98 | caminar-trote |
| RF2 | 9 | 102.14 | 10° SE | 1.57 | 1.51 | caminar |
| RF3 | 9 | 109.74 | 12° SE | 1.66 | 1.52 | caminar |
*La caracterización del modo de progresión es conforme a Alexander (1976) y Thulborn (1982).
La altura promedio de las extremidades de los camélidos de Pie de Vaca es mayor (en un 30%) en comparación a la que alcanzan vicuñas y alpacas (ca. 0.85 m), mientras que es ligeramente más grande (alrededor de un 10%) a la de guanacos y llamas (ca. 1.10 m) (Bonacic y Franklin, 2006; Van der Sluijs et al., 2010). Las llamas del género Hemiauchenia del Plioceno y Pleistoceno de Norteamérica son algo más grandes que una llama actual (Lange, 2002). Por lo anterior, es probable que los camélidos que generaron los rastros hayan sido representantes de ese género de lamines, el cual fue bastante común a lo largo del territorio nacional durante el Cenozoico tardío (Jiménez-Hidalgo y Carranza-Castañeda, 2010; Bravo-Cuevas et al., 2012, 2016a).
La dirección de los rastros, los valores de velocidad y modalidad de progresión de los organismos que los produjeron, así como la profundidad de las huellas, son evidencia utilizada para establecer un comportamiento gregario a través de huellas (Castanera et al., 2012). La distribución y dirección de los rastros de camélidos de Pie de Vaca se asociaría a un grupo de cinco individuos que se dirigen hacia el sureste (RC 1, 2, 5, 8 y 9) y otro grupo de cuatro individuos que se desplazan hacia el norte-noreste (RC 3, 4, 6 y 7). Aunado a esto, el traslape que se observa entre los rastros 5 y 6, así como entre los rastros 1 y 2 con los rastros 3 y 4 (Figura 3A y 3B), indica que los grupos propuestos deambularon por el sitio en momentos diferentes. Con base en la dirección de los rastros, el hecho de los organismos que los produjeron exhiben una velocidad y modo de progresión semejante, así como a la homogénea profundidad de las huellas (≈ 4 cm), se propone un comportamiento gregario que a su vez sugiere cierta interacción social.
Entre las especies actuales de llamas, solamente Lama guanicoe (guanaco) y Vicugna vicugna (vicuña) son salvajes y se distribuyen exclusivamente en Sudamérica (Lange, 2002); ambas especies muestran una marcada organización social y comportamiento territorial (Tomka, 1992). Los guanacos constituyen tres grupos sociales principales: (1) grupo familiar, (2) tropas de machos y (3) machos solitarios. Los grupos familiares están integrados por un macho adulto, el harem de cinco o seis hembras adultas, una o dos crías y un juvenil. Las tropas de machos están formadas por tres y hasta 60 jóvenes-adultos que no son territoriales. Los machos solitarios suelen ser individuos adultos maduros con o sin territorio (Raedeke, 1979; Pérez-Lorente et al., 2009; Franklin, 2011).
La poca variación en la altura promedio de las extremidades de los individuos que generaron los rastros de Pie de Vaca, sugiere un grupo de camélidos de tamaño semejante. Con base en el comportamiento gregario que testifica el registro icnofosilífero y asumiendo una organización social similar a la que se observa en los guanacos actuales, es probable que los rastros de camélidos de Pie de Vaca evidencien la presencia de tropas.
Como ya se ha mencionado, las huellas de camélidos de Pie de Vaca son muy homogéneas en tamaño y morfología, lo cual en la mayoría de los casos imposibilita distinguir las impresiones manuales de las podiales. Sin embargo, la sucesión de huellas que forman los rastros indica una alternancia de las extremidades (anterior/posterior) asociada a pasos bien distanciados que resulta en un patrón de distribución de las impresiones a manera de zigzag. Este patrón es muy semejante al que se produce cuando un animal cuadrúpedo está en canter (Alemán: Galopp) (fig. 2 D en Collins y Stewart, 1993: 335). El canter es un movimiento semejante al galope, aunque relativamente más lento, en el que las extremidades anterior derecha/ posterior izquierda se mueven juntas en una fase (Webb, 1972; Collins y Stewart, 1993; Kienapfel et al., 2014). La velocidad promedio de los camélidos de Pie de Vaca es mayor en comparación a la velocidad promedio de una llama o una alpaca al caminar, la cual es de 1.13 m/s y 0.97 m/s respectivamente (Pfau et al., 2011; Kienapfel et al., 2014). Con base en esto, se propone que los camélidos que generaron los rastros de Pie de Vaca estaban realizando un caminar rápido similar al canter.
5.2. Afinidad del productor de las huellas de félidos
La configuración de estas huellas es semejante a las impresiones de félidos en los siguientes rasgos: (1) digitígradas, (2) arcada de los dígitos semicircular y ubicada enfrente del cojinete interdigital, (3) cojinete interdigital que ocupa una buena parte de la huella, así como (4) impresiones de las garras usualmente ausentes (Remeika, 2001; Sarjeant et al., 2002).
Las huellas de félidos de Pie de Vaca son similares a las impresiones referidas a macairodontinos del Plioceno de Jalisco (Rodríguez-de la Rosa et al., 2007) y a las impresiones referidas al león de las cavernas Panthera spelea del Pleistoceno tardío del norte de Alemania (fig. 2 en Diedrich, 2011: 1885) en la presencia de un cojinete interdigital grande de forma subtriangular a semiredondeada, así como arcada de los dígitos con borde anterior convexo. En lo que respecta al tamaño, son más cercanas a las atribuidas al león de las cavernas de Alemania (longitud de aproximadamente 14 cm) y relativamente más grandes que las de los macairodontinos de Jalisco (longitud de 10 cm).
En el Plioceno de California se conoce un conjunto de huellas significativamente más pequeñas (longitud promedio de 6 cm) en comparación con las de Pie de Vaca, las cuales se asociaron a las especies extintas Felis lacustris (félido semejante a un lince) y Acinonyx (Miracinonyx) studeri (chita semejante en tamaño a un puma) (figuras 8 y 9 en Remeika, 2001: 62 y 64 respectivamente) del Blancano - Irvingtoniano del occidente de Norteamérica (Werderlin, 1985) y del Irvingtoniano temprano de California (Jefferson y Tejada-Flores, 1995), respectivamente.
El tamaño y morfología de las huellas de félidos de Pie de Vaca son indicativos de una forma de talla mediana a grande, tal vez un panterino o un macairodontino; sin embargo, la pobre definición de las huellas imposibilita precisar la identidad de sus productores.
5.2.1. Tamaño, velocidad y modo de progresión
La altura promedio de las extremidades de los félidos es aproximadamente de 1 m. El individuo que generó el RF 1 correspondería al más pequeño y el más grande al que generó el RF 3 (Tabla 3). El RF 1 tiene una dirección al noroeste y luego al sureste, así como se traslapa con los rastros 6 y 7 de camélidos. Los RF 2 y RF 3 son paralelos, dirigidos hacia el sureste y espaciados por unos 8 m; en particular, el RF 2 se traslapa con el rastro 9 de camélidos (Tabla 3, Figura 3A y 3C). La velocidad promedio de los individuos que generaron los rastros es de 1.88 m/s. La proporción LZ/AE es en promedio de 1.67 y se asociaría a organismos en caminata; sin embargo, la determinada para el individuo que se asocia al rastro 1 (LZ/AE ≈ 2) sugiere un cambio en el modo de progresión de caminar a trotar (Alexander, 1976; Thulborn, 1982) (Tabla 3).
La altura promedio de las extremidades de los félidos de Pie de Vaca (ca. 1.0 m) es indicativa de organismos de talla mediana a grande (Kleiman y Eisenberg, 1973), es mayor en comparación con la de un puma (Puma concolor; AE = 0.75 m) (Currier, 1983), con la de un leopardo de las nieves (Uncia uncia; AE = 0.60 m) (Hemmer, 1972), con la de un leopardo africano (Panthera pardus; AE = 0.64 m) (Stein y Hayssen, 2013) y con la de un chita (Acinonyx jubatus; AE = 0.80 m) (Krausman y Morales, 2005); asimismo, es algo más pequeña (en aproximadamente un 15%) que la de un tigre (P. tigris) y un león africano (P. leo) (Mazák, 1981; Haas et al., 2005). En promedio, el león americano (Panthera atrox) del Pleistoceno tardío de Norteamérica es un 33% más grande que un león africano actual (Van Valkenburgh et al., 2016; Bravo-Cuevas et al., 2016b). Por otra parte, los félidos del género Homotherium del Plioceno - Pleistoceno de Norteamérica son de un tamaño similar al de un león africano. Por lo tanto, los félidos que generaron los rastros de Pie de Vaca fueron semejantes en tamaño a un tigre dientes de cimitarra y más pequeños que un león americano. Esto indicaría la presencia de macairodontinos en lo que ahora es la región centro-meridional del estado de Puebla durante el Cenozoico tardío. Cabe señalar que se ha propuesto la presencia de este grupo de félidos a partir de huellas en otras localidades del Neógeno tardío del occidente de México (Rodríguez-de la Rosa et al., 2007; Rodríguez-de la Rosa y Guzmán-Gutiérrez, 2012). La orientación diferente entre el RF 1 y los RF 2 - RF 3, así como el traslape de estos rastros con otros producidos por camélidos, sugiere que los félidos que los produjeron pasaron en momentos diferentes. La trayectoria del RF 1 indicaría que el individuo que lo generó eventualmente cambió de dirección. Por su parte, la orientación paralela de los RF 2 y RF 3, así como una velocidad relacionada con un modo de progresión en caminata para los organismos asociados a estos rastros, son evidencias que conducen a considerar una pareja de félidos.
La organización social en grupos entre los grandes félidos actuales es poco frecuente. Con relación a esto, el león africano es el único que desarrolla una marcada organización social (Rudnai, 1973; Nowell y Jackson, 1996; Anton y Turner, 1997; Packer, 2006). Otros grandes félidos como los tigres muestran cierto grado de sociabilidad, los chitas suelen formar coaliciones temporales de machos jóvenes, mientras que los jaguares y leopardos suelen ser solitarios (Anton y Turner, 1997; Packer, 2006). Entre los félidos extintos se ha considerado que el tigre dientes de sable (Smilodon fatalis) del Pleistoceno tardío de Norteamérica tuvo un estilo de vida sociable, formando grupos familiares constituidos por parejas monógamas (Salesa et al., 2006); asimismo, un alto porcentaje de individuos juveniles y adultos de esta especie encontrados en la localidad Rancho La Brea, sugiere que era social como los lobos y leones actuales (McCall et al., 2003; Salesa et al., 2006; Carbone et al., 2009). El león americano Panthera atrox se sugiere que pudo ser solitario o tal vez organizarse en parejas (Jefferson, 2001; Salesa et al., 2006; Carbone et al., 2009; Bravo-Cuevas et al., 2016b). La información acerca de los macairodontinos del género Homotherium sugiere que eran solitarios, debido al escaso número de individuos encontrados en Rancho La Brea (Carbone et al., 2009).
Con base en lo anterior y en función de que el registro de huellas de félidos de Pie de Vaca testificaría de manera indirecta la presencia de un individuo (RF 1) y una pareja (RF 2 y RF 3) que transitaron en momentos diferentes la zona, se propone un comportamiento semejante al que se ha considerado para el león americano y el tigre dientes de cimitarra.
Se sabe que los leones africanos y los tigres en caminata alcanzan velocidades promedio de 1.4 m/s y 1.2 m/s respectivamente. Las modalidades de trote y galope son raras y poco frecuentes, aunque leones en trote muestran velocidades de alrededor 2 m/s (Rudnai, 1973). Anton et al. (2004) consideraron que la velocidad de los félidos que produjeron algunos de los rastros referidos a Felipeda lynxi y F. parvula del Mioceno de España, se asociaría a organismos en trote con una velocidad promedio de 1.3 m/s.
Con base en esto, la velocidad promedio de los félidos de Pie de Vaca (1.8 m/s) se asociaría a una caminata rápida o trote.
5. Conclusiones
Se describe el conjunto de huellas de camélidos y félidos de la localidad Pie de Vaca en el estado de Puebla, centro de México; las de camélidos fueron designadas a la icnoespecie Lamaichnum guanicoe en atención a su tamaño y se sugiere que fueron producidas por algún integrante del género Hemiauchenia que fue la llama más común del Cenozoico tardío de México. Por su parte, las huellas de félidos fueron asignadas a la morfofamilia Felipedidae; posiblemente fueron producidas por alguna forma de talla mediana a grande como un macairodontino o un panterino. El modo de progresión de los camélidos y félidos se relaciona con organismos en caminata relativamente rápida y desplazándose a una velocidad inferior a los 4 m/s. La distribución y las direcciones de los rastros de camélidos, así como la velocidad y modalidad de progresión propuestas para los individuos que los generaron, indican un comportamiento gregario asociado a cierta organización social, tal vez tropas de llamas. En el caso de los félidos se propone un comportamiento solitario y/o en pareja, como el que se observa en la mayoría de los grandes félidos actuales.

