











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink1. Introduction
Shape analysis plays an important role in many kinds of biological studies (Zelditch et al., 2004). One such kind of study is ichnology: using shape (besides wall and filling) as a basic characteristic to define insect-trace fossils (Genise, 2017). In recent years, geometric morphometric techniques have been more frequently used in morphological analyses of different fossil taxa, including ostracods (e.g.,Elewa, 2004), neurosauropods (e.g.,Bonnan, 2007), brachiopods (e.g.,Bose, 2013), rodents (e.g.,Wilson, 2013; Calede and Glusman, 2017; Smith and Wilson, 2017), euprimates (e.g.,Bennet and Goswami, 2012) and hominins (e.g.,Bouhallier et al., 2004; Gómez-Robles et al., 2008), however, these techniques are uncommon in ichnological studies.
Geometric morphometric methods have been used in vertebrate trace fossils to infer the potential producer. Regarding dinosaur trackways studies, Rodrigues and Faria dos Santos (2004) characterized variation in shape across various, previously reported Sauropodomorpha ichnotaxa and other dinosaur footprints. Lallensack et al. (2016) used these methods in order to distinguish ornithopod and theropod footprints from the Upper Cretaceous in Münchehagen, Germany. Regarding mammals trackways, Ledoux and Boudadi-Maligne (2015) analyzed modern prints from two wolf subspecies and seven breeds of dogs. These results were compared with presumably dhole tracks documented at Chauvet Cave, France.
On the other hand, Fictovichnus gobiensis is a common ichnotaxon reported in the late Eocene outcrops from Santiago Yolomécatl, Oaxaca, southern Mexico (Guerrero-Arenas et al., 2017) (Figure 1a). F. gobiensis was originally described by Johnston et al. (1996) as being from the Upper Cretaceous Bayan Mandahu Formation of China and the Upper Cretaceous Djadokhta Formation of Mongolia. Their diagnosis was later amended (Alonso-Zarza et al., 2014). This ichnospecies is commonly preserved as hollow structures in paleosols, or as ellipsoid, with smooth casts and lacking a discrete wall (Genise, 2017). They show rounded extremes and a smooth surface in casts detached from the matrix. F. gobiensis is very frequent in carbonate rich paleosols. This ichnospecies is attributable to weevil or chafer pupation chambers, as well as other groups of Coleoptera (Johnston et al., 1996; Genise, 2017).

Figure 1 a) Fictovichnus gobiensis in situ, from locality Oax-13 La Cruda, Santiago Yolomécatl, Oaxaca, Mexico. Diameter of coin is 21 mm. b) Specimen (UMLP-004) showing marks. Landmarks are represented by black dots; red dots are semi-landmarks; blue dots are semi-landmarks considered as helper points. Scale are equal to one centimeter.
Traditionally, in studies of invertebrate trace fossils, the producers are not identified, given that different tracemakers may produce identical structures when behaving similarly (Bromley, 1996). However, in the case of insect pupation chambers, if the identity of potential producers could be determined, this information might be linked to more specific environmental conditions and even additional paleoecological information such as trophic relations in the paleocommunities could be obtained.
The objective of this study is to evaluate the variation in shape across specimens of F. gobiensis collected from a single bed in Santiago Yolomécatl outcrops. Potential producers of the ichnospecies include several taxa (Genise, 2017); thus, we expect that the shape of chambers reflects such different potential producers.
2. Material and methods
Forty-two fossil pupation chambers of Fictovichnus gobiensis were examined. The specimens were selected for their completeness and preservation quality. They were collected in outcrops of the Yolomécatl Formation, in southern Mexico (for detailed description of the area and the location of localities, see Jiménez-Hidalgo et al., 2015, 2017). All the specimens come from a single stratigraphic level.
Color images of the specimens were captured using a Kodak EasyShare C195, with the same conditions of fixed focus, camera angle and magnification ratio. The scar marking the exit hole determined the orientation of the specimens; when it was not present, the traces of the scar were used. Two-dimensional (2D) geometric morphometrics (GM) were used to collect shape data. Two landmarks (Type 3, sensu Bookstein, 1991) were positioned in the most exterior points of the rounded extremes of the specimens (point numbers 1 and 19), and 34 semilandmarks (point numbers 2 - 17 and 18 - 36) were positioned in the ellipse using a drawing fan generated by MakeFan8 (Sheets, 2010a) (Figure 1b). Two-dimensional coordinates of all the marks were digitized using tpsDig2 (Rohlf, 2013). Coordinates were imported into CoordGen 8 (Sheets, 2014a) to proceed to Procrustes superimposition followed by the semilandmark alignment with SemiLand 8 (Sheets, 2010b). The superimposition and alignment eliminated variation that was not due to form (translation, rotation and scaling). In the alignment, sixteen semilandmarks were used as helper points (4, 6 - 8, 12 - 14, 16, 22, 24 - 26, 30 - 32, and 34). Helper points were excluded from subsequent analysis, resulting in the reduction of the final variables number to two landmarks and 18 semi-landmarks.
A Principal Component Analysis (PCA) was performed with PCAGen (Sheets, 2014b) with the aim to explore the shape variation across individuals. Procrustes distances were calculated as a measure of total similarity among shapes; the matrix with pairwise distances was imported into the software PAST (Hammer et al., 2001) for cluster analyses and the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) and hierarchical grouping algorithm was used. Resulting clusters were analyzed with Canonical Variates Analysis (CVA) using CVAGen 8 (Sheets, 2010c) to estimate a MANOVA (Multivariate analysis of variance) for the differences in shape among groups.
3. Results
PCA analysis explains 78% of the variance in the first two axes, 50% in the first principal component and 28% in the second principal component (Figure 2a). The deformation grid over the two principal components shows a deformation along the ellipse’s contour, making some of them elongated and thinner, while others were shorter and wider (Figure 2b).
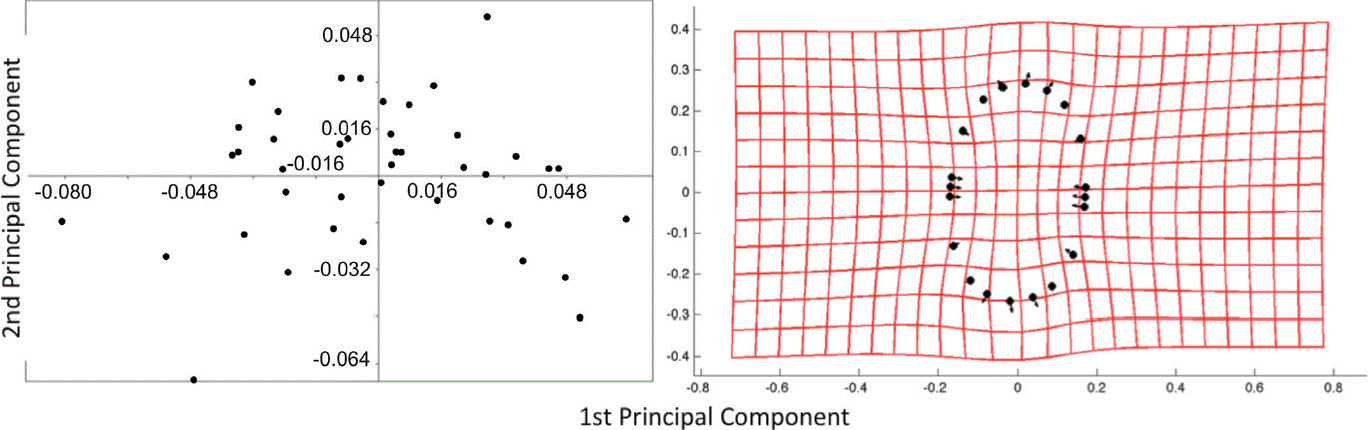
Figure 2 a) Principal Component Analysis of studied specimens. b) Deformation grid; the specimens varied from elongated and thinner to shorter and wider.
Cluster analyses with UPGMA formed four groups, three of them were segregated in the first principal component and the fourth one could be seen over the second component represented by only one individual (Figure 3). Although UPGMA analysis suggested four groups, CVA analysis was done with three of them (1, 2 and 4 in Figure 3) because of the lack of variance in the group number 3, composed of only one specimen (Figure 4). In Figure 4, the CVA analyses showed two significant distinct CVA axes on which the three morphs are clearly differentiated (Table 1). MANOVA analysis suggests that 78.4% (p < 0.001) of the variance is explained by the morph groups’ differences (Table 2).
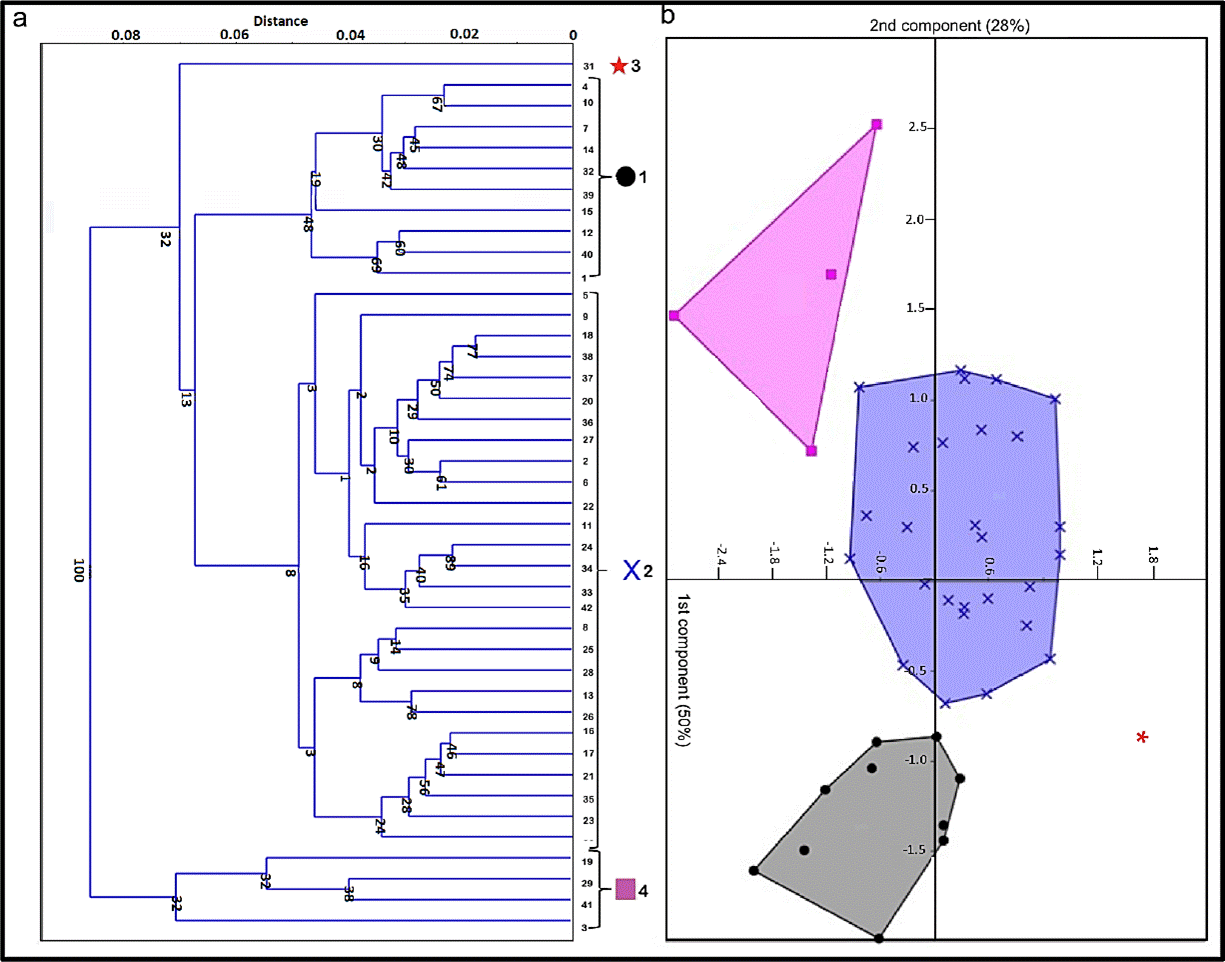
Figure 3 a) Clustering with UPGMA. Each symbol represents a different group of F. gobiensis specimens. b) Principal component analysis of F. gobiensis specimens.
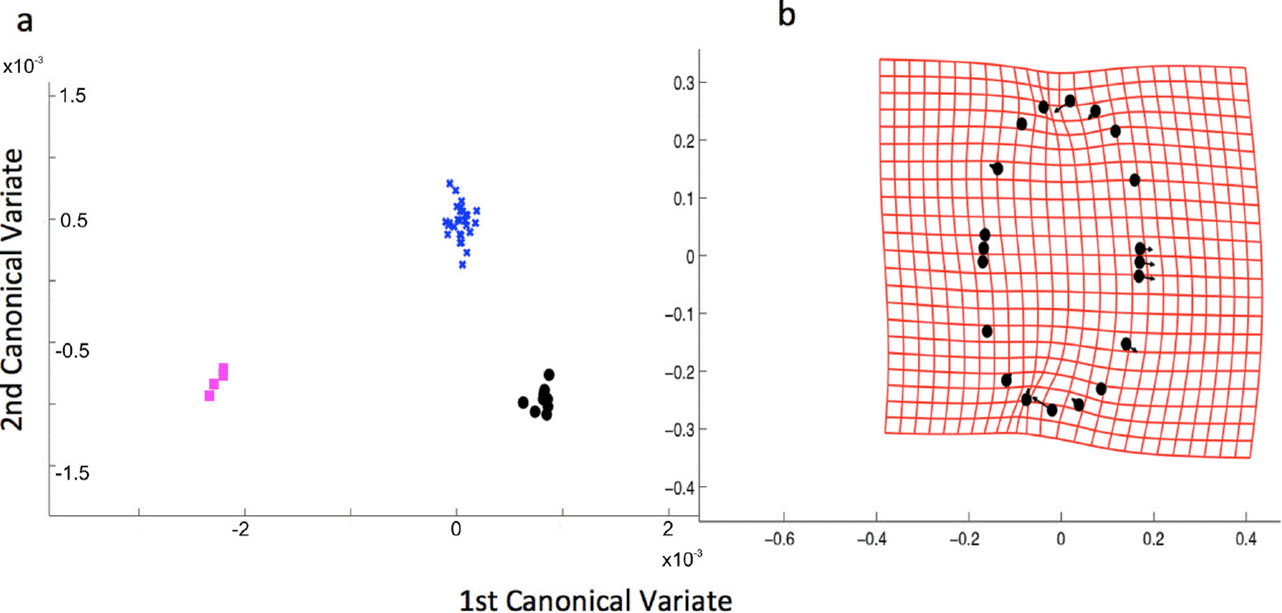
Figure 4 Canonic Variate Analysis of the shape of the recognized morphs. a) CVA of the three detected morphs. b) Deformation grid; the contour near the extremes showed the major variation.
Table 1 Canonical Variate Analysis.
| AXE | EIGENVALUE | LAMBDA | CHI SQUARE | DF | p |
|---|---|---|---|---|---|
| 1 | 132.111 | 0.003 | 169.538 | 72 | 7.672 e-10 |
| 2 | 28.339 | 0.0341 | 69.2684 | 35 | 0.0004906 |
DF: Degrees of freedom.
4. Discussion
The four morphs of Fictovichnus gobiensis identified in Yolomécatl suggest the presence of at least four potential producers. At present, there is scarce information about traditional morphometrics to infer the nature of insect trace fossil producers. However, several studies suggest the potential to use the shape as criteria for identification of producers. Sánchez et al. (2010) analyzed the relative sizes of Coprinisphaera specimens of Sarmiento Formation, Argentina, to infer the potential diversity of producers. In this kind of trace fossils, it was demonstrated that size of dung beetle brood balls could be directly related to the size of their producers (e.g.,Halffter and Matthews, 1966; Sato and Imamori, 1987).
Producers of F. gobiensis could be larvae of soil-dwelling coleopteran groups related to Curculionidae, Tenebrionidae and Scarabaeidae (Johnston et al., 1996; Genise, 2017). Available descriptions of shape or size of similar structures deposited in recent soil produced by these groups do not fit with Fictovichnus gobiensis chamber features. There are a few reports of brood masses of dung beetles (Scarabaeidae); Romero-Samper and Martín-Piera (1995) described simple branched brood masses of Onthophagus stylocerus. Huerta and García-Hernández (2013) measured volume and weight of brood masses of Onthophagus incensus. However, these structures are not comparable with F. gobiensis.
Differences in the morphology of pupal fossils could be related with the nature of a specific producer. Further analysis of chamber shape from modern taxa of Curculionidae, Tenebrionidae and Scarabaeidae will clarify the nature of F. gobiensis producers, and help us understand if shape variation is linked with different species, sex of pupal specimens, or even some environmental factors.
In our study, we concluded that the four recognized morphs suggest four potential producers. If every morph represents different taxa, we should have a community with at least four kinds of beetles. Vegetation type inferred from ichnofossils, isotopic values of pedogenic carbonates and the mammalian fauna suggest scrubland/woodland paleoenvironments in the Yolomécatl area (Guerrero-Arenas et al., 2017). In modern analogue communities, the diversity of beetles is comparable with our results. In a desert shrubland-grassland ecotone in Socorro County, New Mexico, two species of Curculionidae were collected into patches of creosotebush (Sánchez and Parmenter, 2002). Five species of Tenebrionidae were trapped in a shrub steppe habitat at Cerro Otto, Bariloche, Argentina (Mazía et al., 2006).
The four morphs recognized in this study by means of geometric morphometrics represent indirect evidence of beetle diversity that were not directly preserved in the fossil record.
5. Conclusions
Our geometric morphometric analysis of the Fictovichnus gobiensis chamber resulted in the recognition of four morphs. This shape variation in filling chambers can be related with a particular producer. Although producers of different taxa could be related directly with shape variation, additional studies are required to determinate if other variables (climate, intraspecific variation, phenotypic plasticity, etc.) influence the shape of F. gobiensis chambers.
Geometric morphometrics is a potential tool in paleoichnological studies, to obtain additional information about producers of trace fossils. However, at present, it is limited by the lack of information of morphometric data in recent analogues. Knowledge of modern structures is crucial to take advantage of this useful analysis.

