











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
The Zagros mountain belt of Iran, a part of the Alpine-Himalayan system, extends from the NW Iranian border to SW Iran, up to the Strait of Hormuz (Figure 1A) (Heydari et al ., 2003). It can be divided into a number of zones (Lurestan, Izeh, Dezful Embayment, Fars, High Zagros), which differ according to their structural style and sedimentary history (Figure 1B). The Asmari Formation is a stratigraphic unit, mostly composed of limestone with subordinate sandstone and shale of approximately Oligocene-Early Miocene age. This formation was developed regionally across the Zagros Mountain range, prograding from the north and the south into the Eocene-Early Miocene Pabdeh Formation (James and Wynd, 1965; Seyrafian et al ., 1996). The Asmari Formation is the main petroleum reservoir in southwest Iran (Beydoun et al ., 1992).
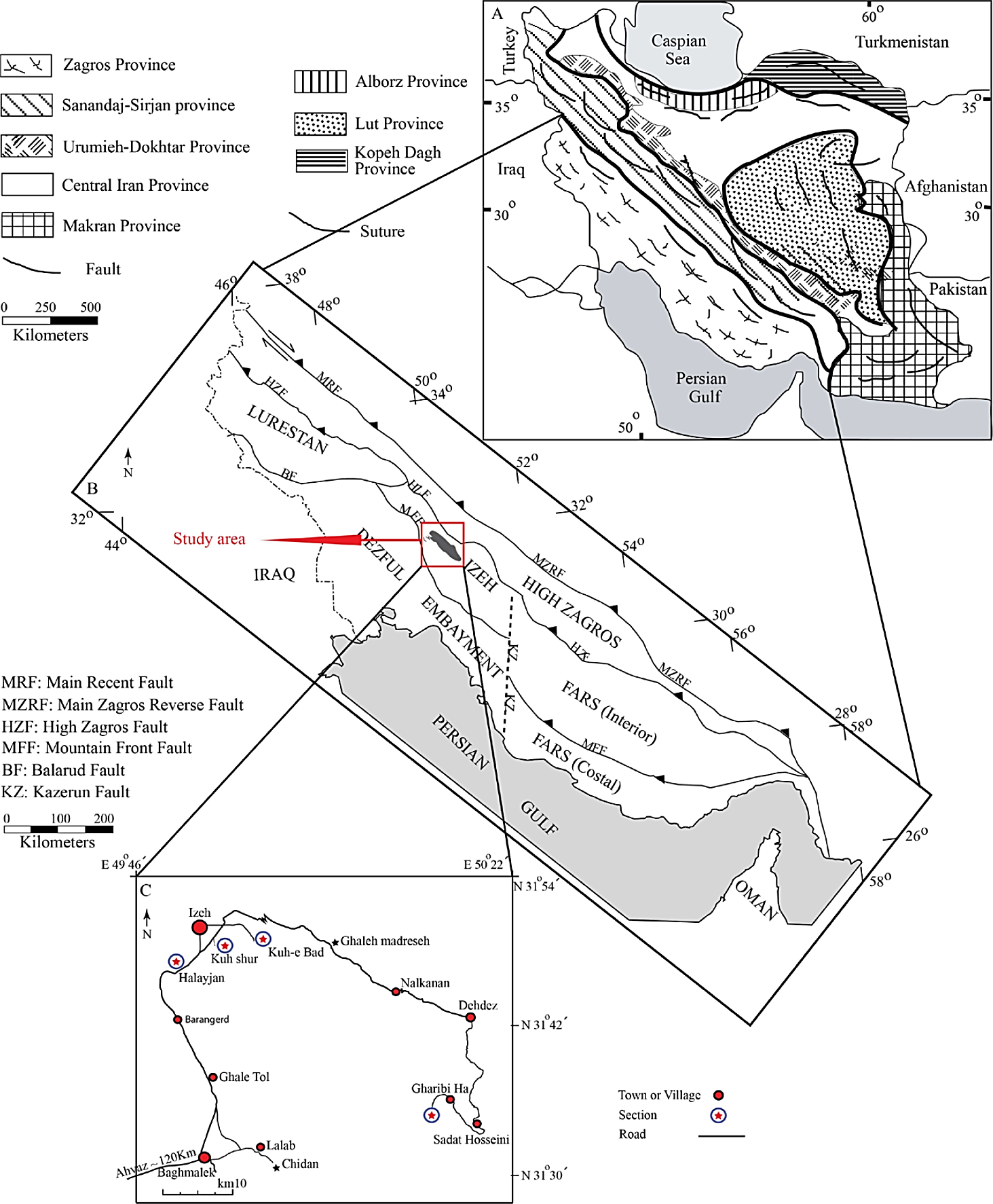
Figure 1 Location of the study area. A) General map of Iran showing eight geologic provinces (adapted from Heydari et al. , 2003). B) Subdivision of the Zagros province. C) Map of the study area in the Izeh zone, southwest Iran. Halayjan (Section 1), Kuh Shur (Section 2), Kuh-e Bad (Section 3) and Gharibi Ha (Section 4). The study area is located at the Izeh Zone adapted from Farzipour-Saein et al. (2009).
Lithologically, the Asmari Formation, at the type section in Tang-e Gel-e Tursh (Valley of Sour Earth) on the southwestern flank of the Kuh-e Asmari anticline, consists of 314 m of mainly limestone, dolomitic limestone and argillaceous limestone (Motiei, 1993). The shallow marine limestones of the Asmari Formation were conformably deposited over the Pabdeh Formation in the southwestern part of the Zagros Basin (Figure 2) and also unconformably covered the Jahrum and Shahbazan Formations to the northeastern part in the Fars and Lurestan regions (Figure 2). The Oligocene-Miocene in SW Iran is mostly characterized by exclusively shallow water carbonates.
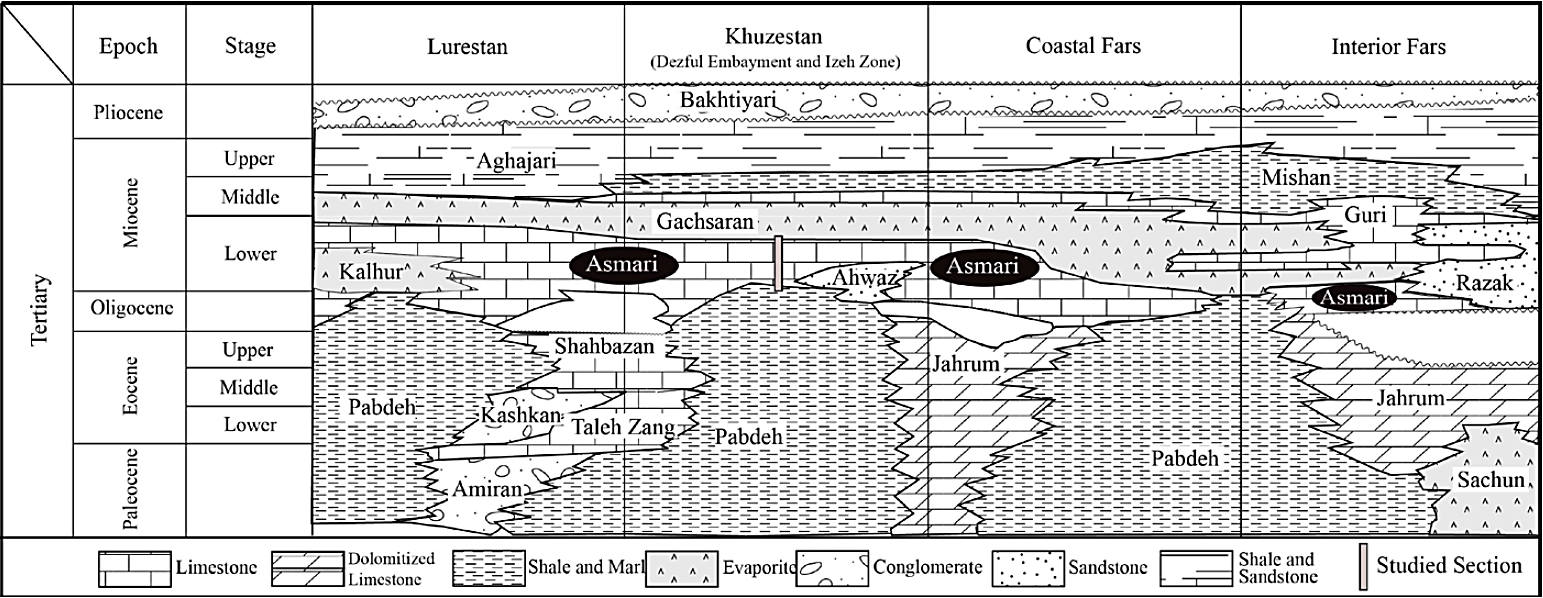
Figure 2 Cenozoic stratigraphic correlation of the Iranian Sector of the Zagros Basin adapted from James and Wynd (1965).
In all studied sections the contact of the Asmari Formation with the underlying Pabdeh Formation is transitional and conformable. The upper contact with Gachsaran Formation is conformable in all studied sections. The Asmari Formation in the study area is mainly composed of thick to medium bedded and massive limestone and subordinate marl and dolomitic limestone intercalations. The overall thickness of the formation vary from northwest to southeast including 330 m, 325 m, 415 m and 416 m in the Halayjan, Kuh Shur, Kuh-e Bad and Gharibi Ha, respectively.
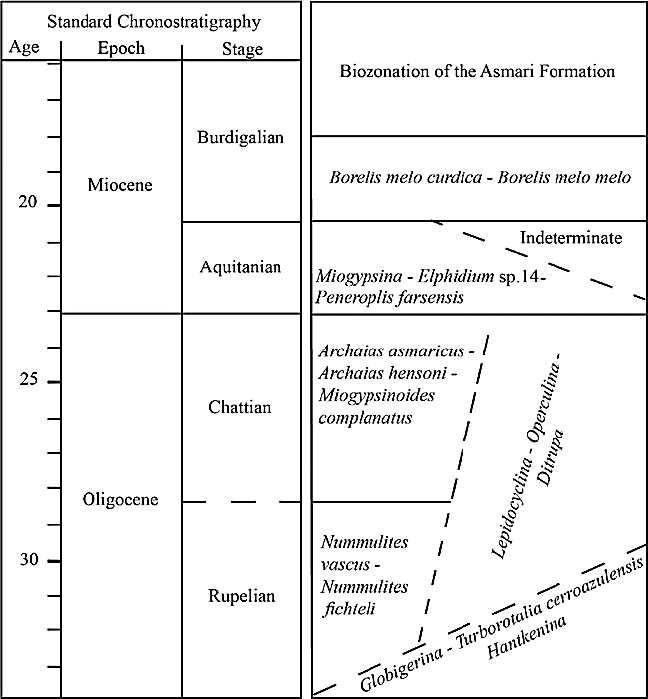
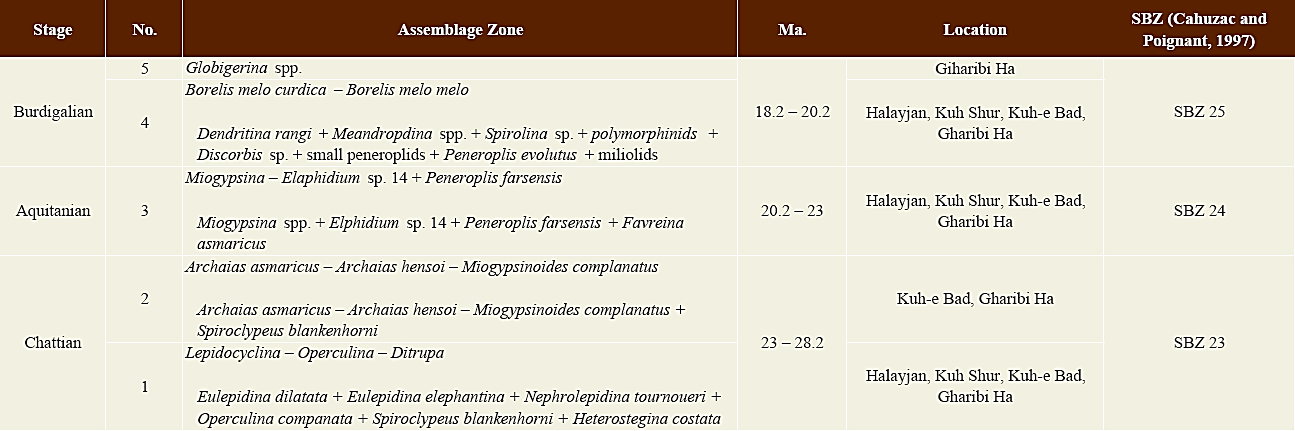
The biostratigraphy of the Asmari Formation was outlined by Wynd (1965) and reviewed by Adams and Bourgeois (1967). Adams and Bourgeois (1967) introduced four assemblage zones for the Asmari Formation (Table 1). They did not differentiate Rupelian from Chattian. These two reports were used by many authors (e.g., Seyrafian, 2000; Vaziri-Moghaddam et al ., 2006; Amirshahkarami et al ., 2007a, b).
Table 1 Biozonation of the Asmari Formation based on Wynd (1965), Adams and Bourgeois (1967) and Cahuzac and Poignant (1997).

Recently, Ehrenberg et al . (2007), Laursen et al . (2009) and Van Buchem et al . (2010) conducted strontium isotope dating to improve the biostratigraphic ages for the Asmari Formation across most parts of the Zagros basin (Izeh zone and Dezful Embayment) (Table 2). This biozonation is used by many recent authors (e.g ., Allahkarampour Dill et al ., 2010; Van Buchem et al ., 2010; Sooltanian et al ., 2011; Rahmani et al ., 2012). The biozonation scheme of Laursen et al . (2009) and Van Buchem et al . (2010) have some basic differences compared to the old scheme (Adams and Bourgeois, 1967). In the old scheme, the noticeable thickness of the Asmari limestone, containing Archaias, has been considered Aquitanian. Today this interval proved to be Oligocene. Also in the biozonation of Laursen et al . (2009) and Van Buchem et al . (2010), the Oligocene sediments are divided to Rupelian and Chattian.
The area of the new biostratigraphic studies which used the biozonation of Laursen et al . (2009) and Van Buchem et al . (2010) is very small in comparison to the total area of the Asmari basin (more than 150 km × 700 km). There are a lot of areas with unpublished sections for which the new biozonation should be considered. The studied area of this research is one of those areas with well preserved and untouched outcrop sections. The data obtained from biostratigraphic studies in this area will be used for the future regional study. The abundance of large benthic foraminifera in the limestones of the Asmari Formation offers an excellent opportunity to study paleoenvironmental controls on larger benthic foraminiferal assemblages in detail. Sedimentary successions, representing the Oligocene-Miocene interval, allow us to evaluate the influence of change in ecological parameters through time.
The objectives of this study are (1) to reconstruct a coherent biostratigraphic framework for the carbonate deposits of the Asmari Formation in the studied area based on available biozonation schemes (Cahuzac and Poignant, 1997; Laursen et al ., 2009; Van Buchem et al ., 2010) (Tables 1 and 2), and (2) to present the paleoenvironmental factors which affect the distribution of biogenic components of the Asmari Formation.
2. Material and method
To achieve the aim of this study, four unpublished sections (Halayjan, Kuh Shur, Kuh-e Bad and Gharibi Ha) (Figures 1C and 3-5 (4)) were measured, described and sampled for paleontological and petrographical studies. The measurement was carried out using Jacob’s staff. The description logs, containing lithology, texture and components (with hand lens), bedding and thickness, was prepared in the field. Around 1000 thin sections were prepared and studied to investigate texture (Dunham, 1962; Embry and Klovan, 1971) components and species identification. In this paper, LBF are used to date the carbonates using biostratigraphical zonation (Wynd, 1965; Adams and Bourgeois, 1967; Laursen et al ., 2009; Van Buchem et al ., 2010) and comparisons with the global Oligocene/Miocene zonation of Cahuzac and Poignant (1997) (Tables 1, 2 and 3).
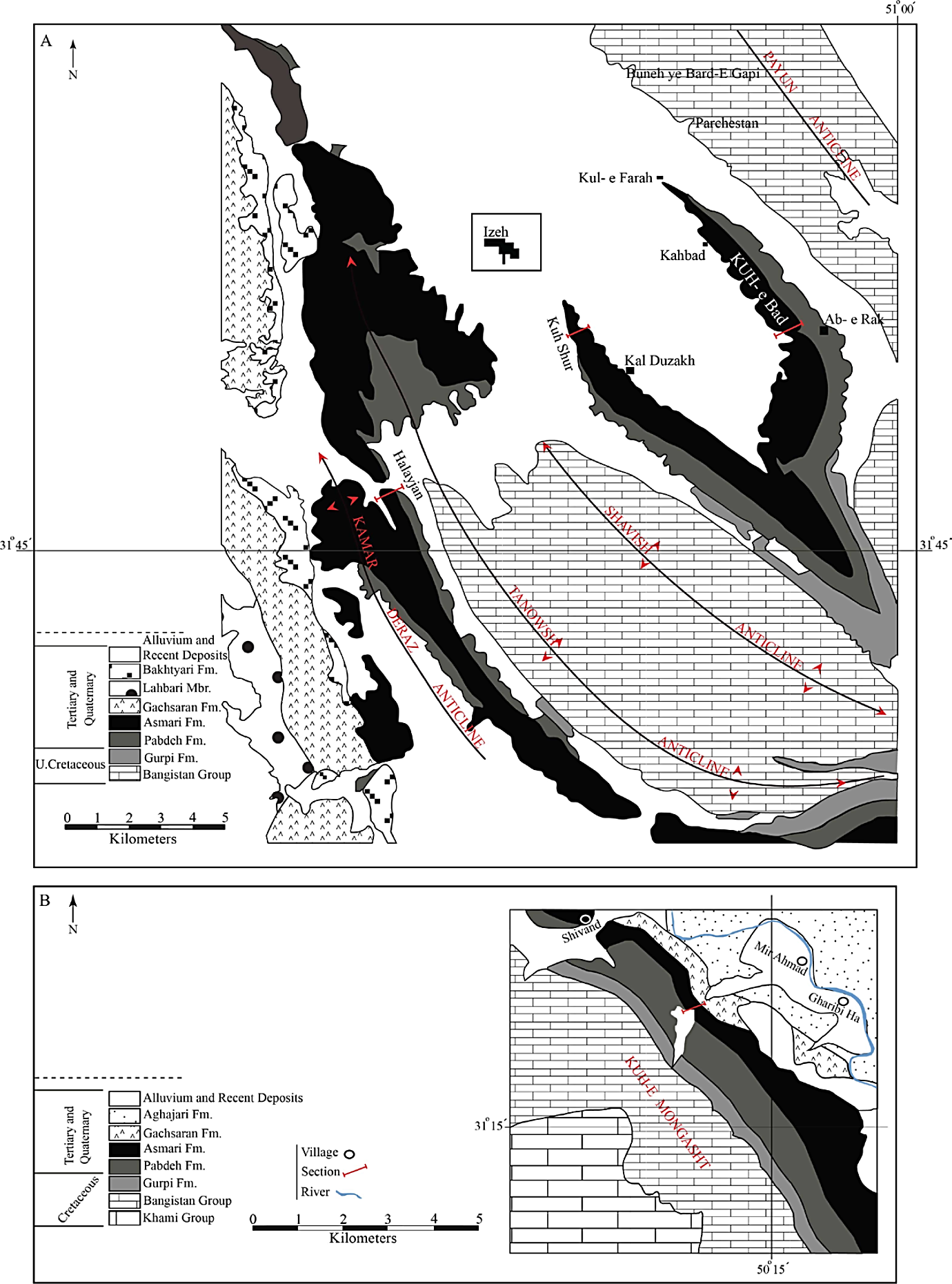
Figure 3 Location and geological map of the study area, southwest Iran. A. Halayjan, Kuh Shur and Kuh-e Bad sections. B. Gharibi Ha section, southwest Iran.


Figure 5 General view of studied sections (a: Kuh Shur; b. Kuh-e Bad and c. Gharibi Ha), view toward north and southeast.
To have the possibility of future reinterpretation and data control, a data set containing ~10000 digital photos were prepared by using the Nikon microscope suite in the National Iranian South Oil Company lab. Nearly all of components are documented in these photos. These photos were classified and used for biostratigraphy and paleoecological interpretation. Based on this method, for every component mentioned in the biostratigraphic log, one or more photo is available.
3. Biostratigraphy
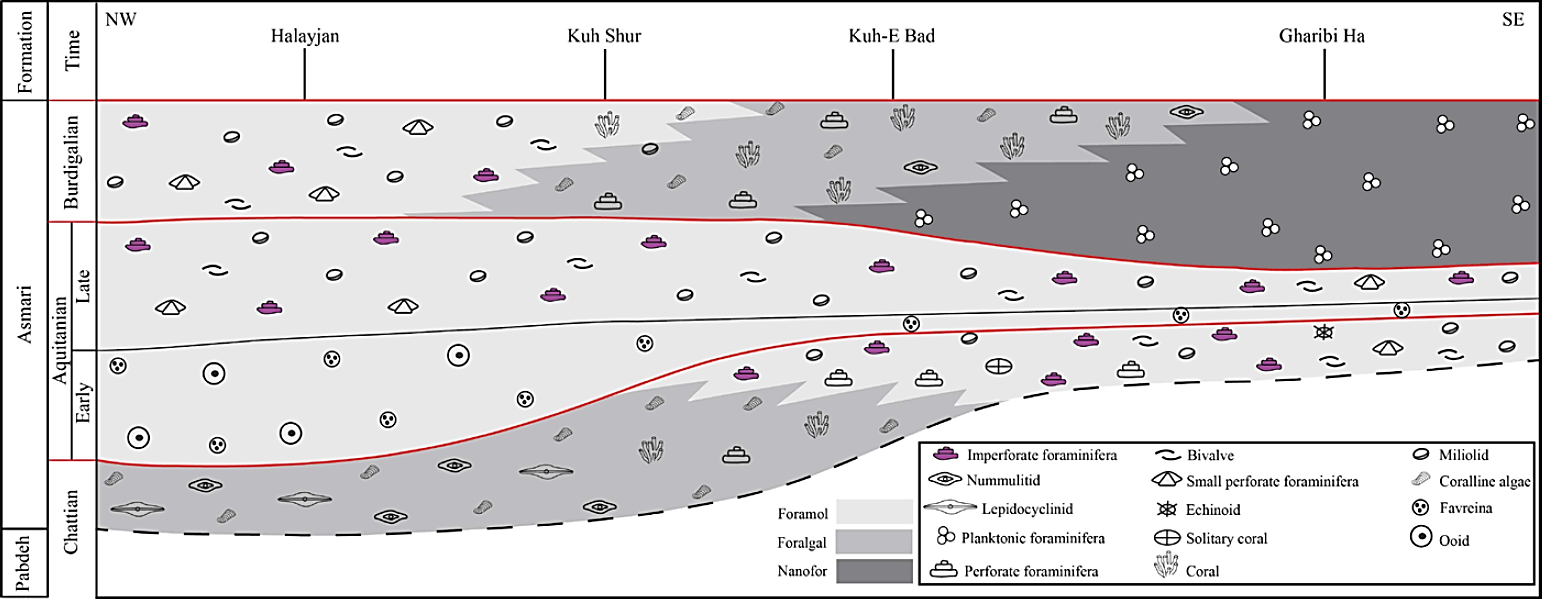
The Oligocene to Miocene benthic and planktonic foraminiferal assemblages, including a total of 17 genera and 23 species, were analyzed to establish the biostratigraphic framework of the Asmari Formation in the studied sections (Figures 6, 7, 8 and 9). From base to top, five Assemblage Zones were recognized (Figures 6, 7, 8, 9 and 10).
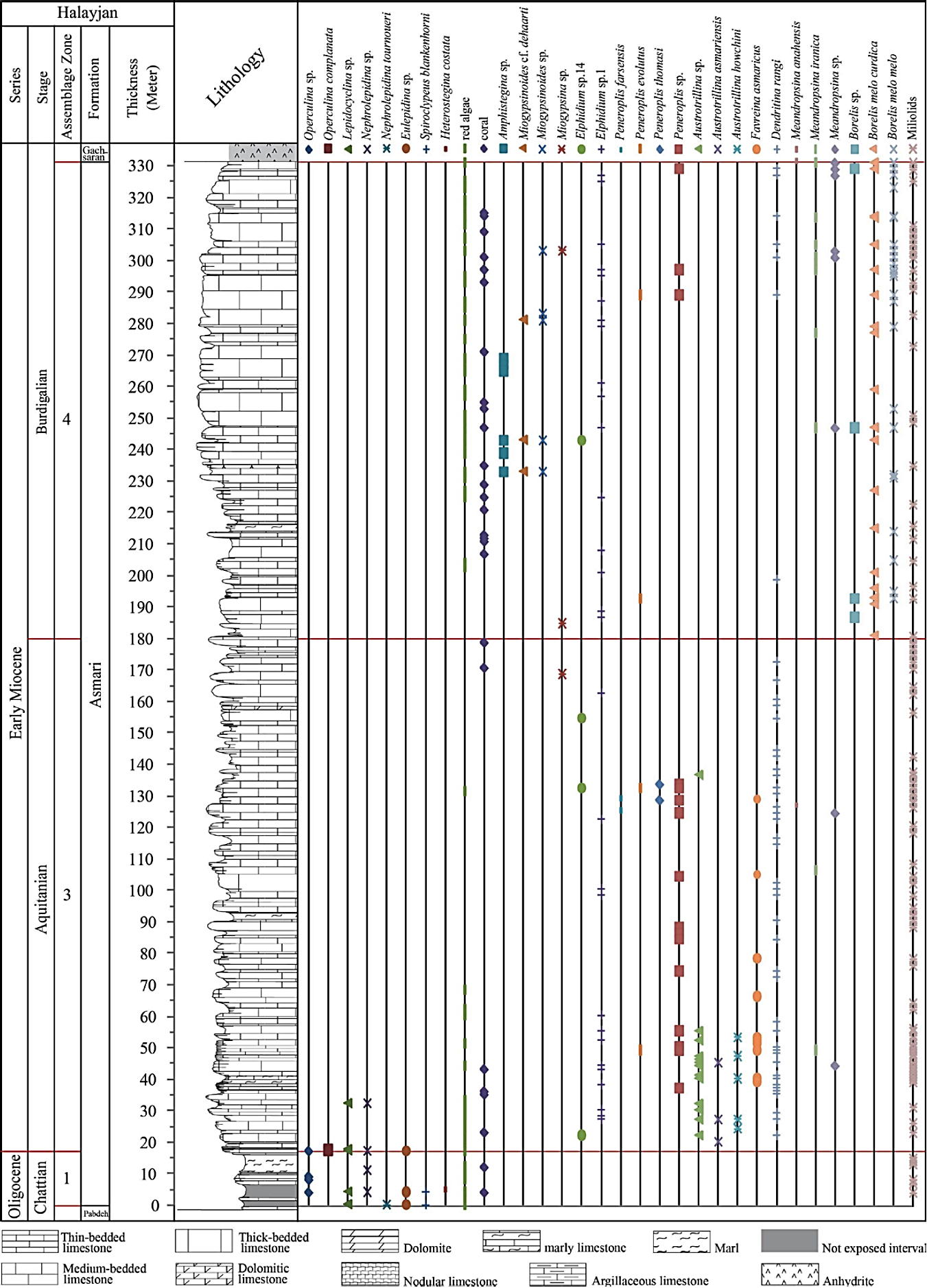
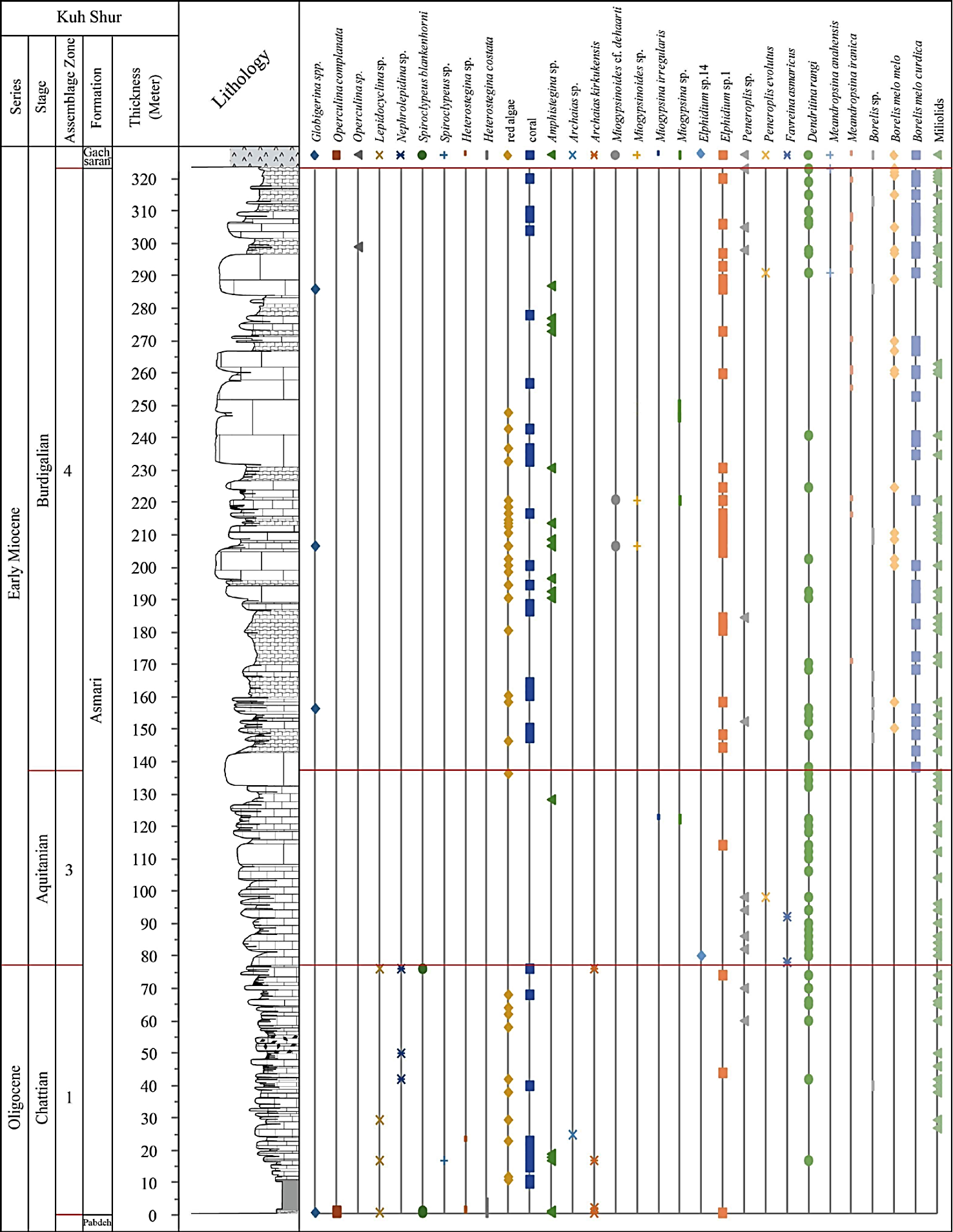
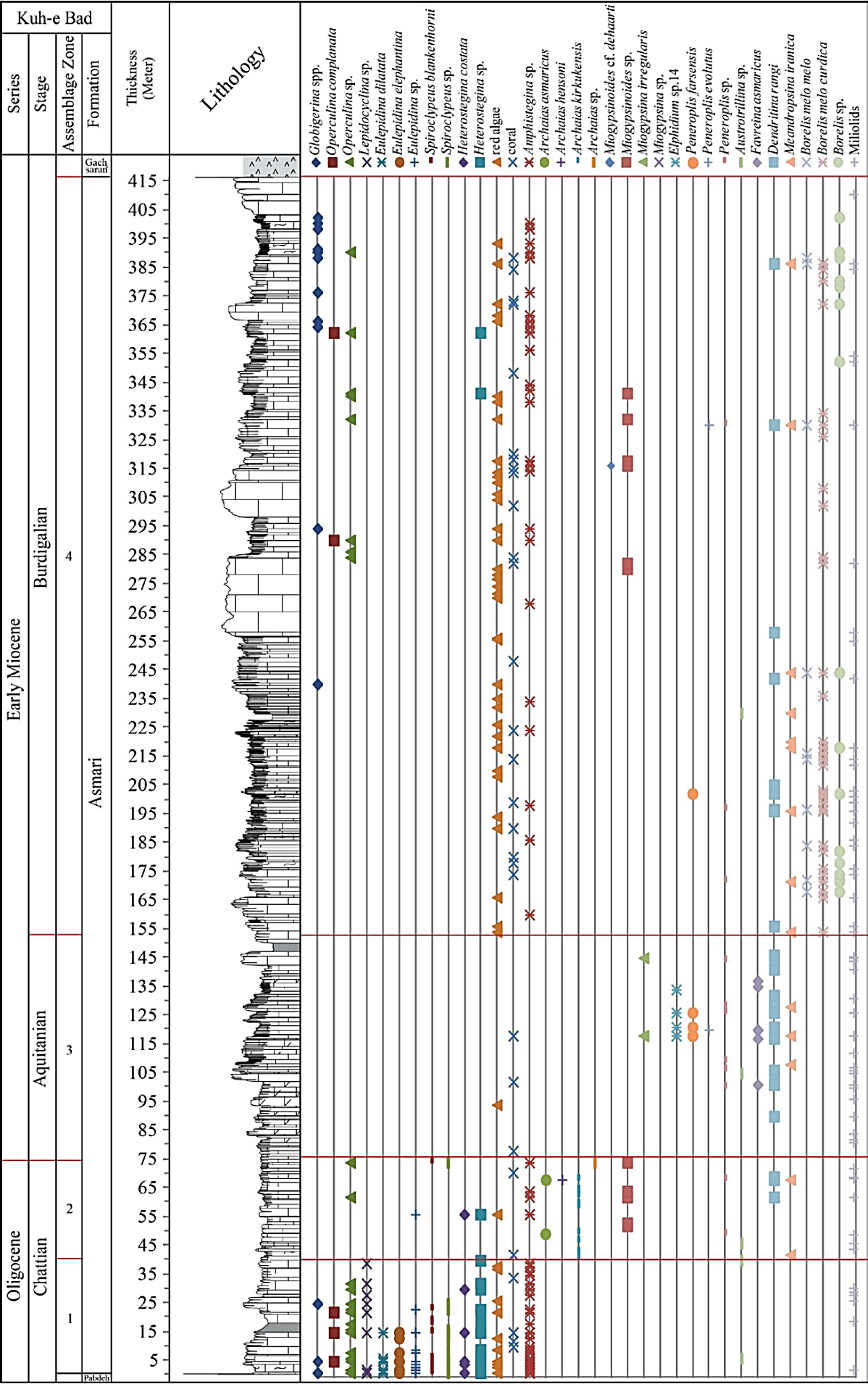
Figure 8 Biostratigraphy of the Asmari Formation at the Kuh-e Bad section (see Figure 6 for legend).
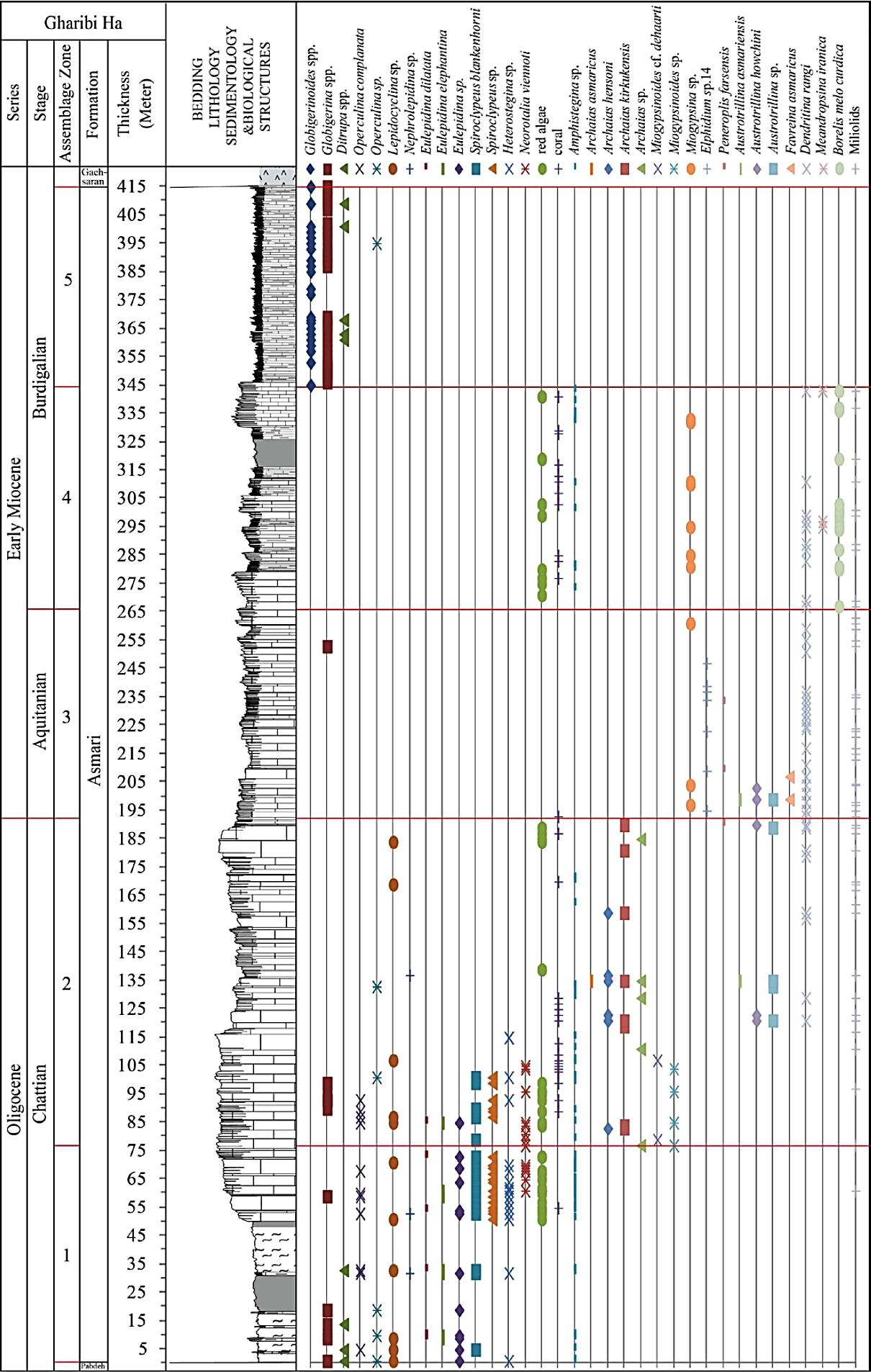
Figure 9 Biostratigraphy of the Asmari Formation at the Gharibi Ha section (see Figure 6 for legend).
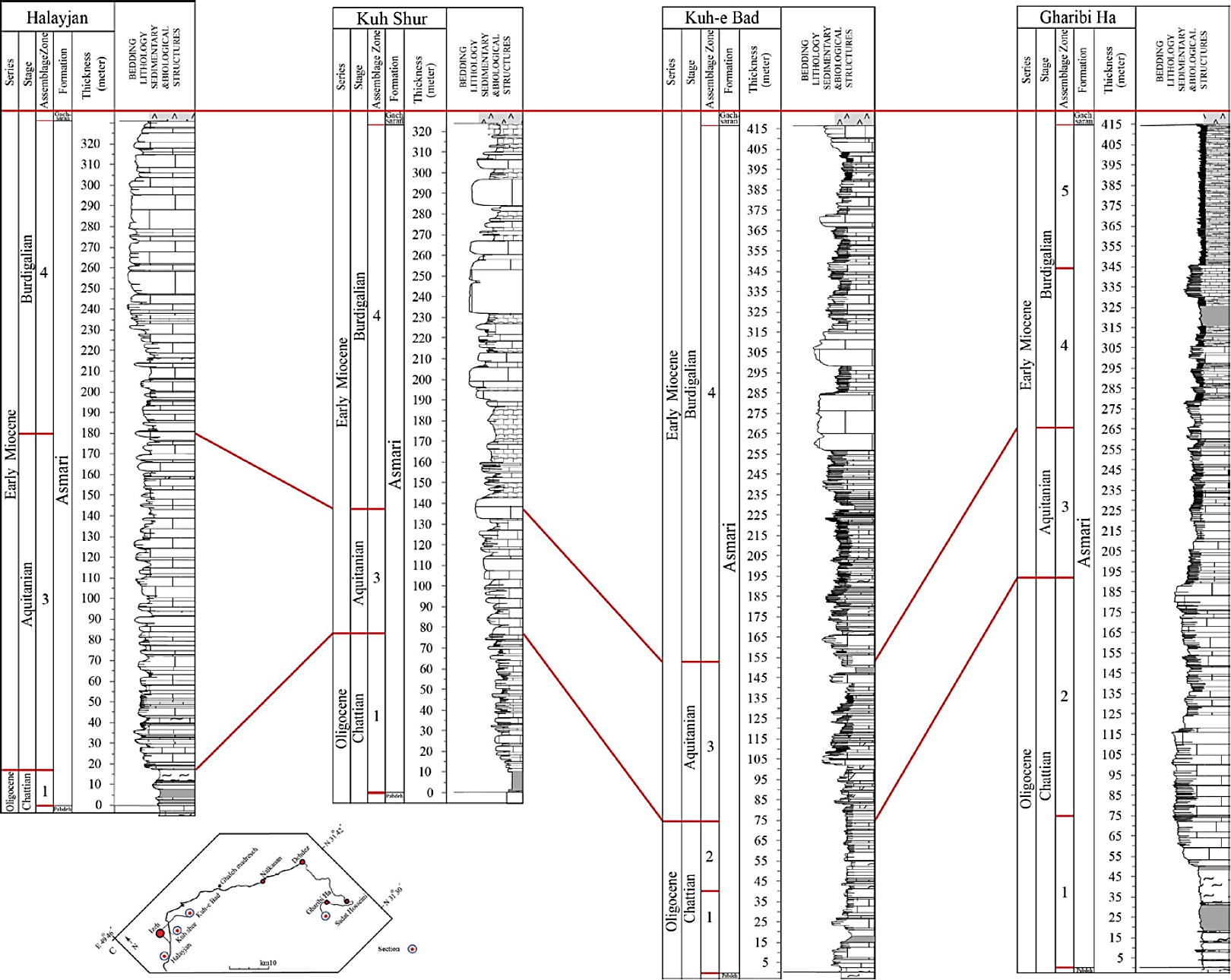
Figure 10 Time correlation of the Asmari Formation between the Halayjan, Kuh Shur, Kuh-e Bad and Gharibi Ha sections (see Figure 6 for legend).
3.1. Assemblage 1
This assemblage is present in all sections and consists of Eulepidina dilatata, Eulepidina elephantina, Eu lepidina sp., Nephrolepidina tournoueri, Nephrolepidina sp., Lepidocyclina sp., Heterostegina costata, Heteroste gina sp., Spiroclypeus blankenhorni, Spiroclypeus sp., Operculina complanata, Operculina sp., Neorotalia vien noti, Amphistegina sp., Ditrupa sp. and globigerinids (Figure 11). This assemblage is correlated with the Lepidocyclina -Operculina -Ditrupa Assemblage Zone of Laursen et al . (2009) and Van Buchem et al . (2010), and to the SBZ 22 of Cahuzac and Poignant (1997).

Figure 11 Photomicrograph of some selected fauna of the Asmari Formation in the studied sections. 1) Eulepidina elephantina (Lemoine and Douville, 1904), axial section, sample number K2; 2) Eulepidina sp., subaxial section, sample number K15; 3) Eulepidina dilatata (Michelotti, 1861), axial section, sample number G8; 4) Eulepidina dilatata, axial section, sample number K3; 5) Nephrolepidina tournoueri (Lemoine and Douville, 1904), axial section, sample number H0; 6) Operculina sp., subequatorial section, sample number H17.5; 7) Operculina complanata (Defrance, 1822), axial section, sample number K22; 8) Heterostegina sp., subaxial section, sample number G54; 9) Heterostegina sp., axial section, sample number G50; 10) Spiroclypeus blankenhorni (Henson, 1937), equatorial section, sample number G61; 11) Spiroclypeus blankenhorni (Henson, 1937), axial section, sample number G67.
Based on Laursen et al . (2009) this assemblage is attributed to the Rupelian-Chattian. However, based on Ehrenberg et al . (2007) who believes that the presence of Spiroclypeus blanckenhorni indicates a Chattian age, the age of this zone must be limited to the Chattian.
3.2. Assemblage 2
This assemblage is recorded in Kuh-e Bad and Gharibi Ha sections. The most diagnostic species include: Archaias kirkukensis, Archaias hensoni, Archaias asmaricus, Archaias sp., Miogypsinoides sp., Elphidium sp.1, Peneroplis sp., Austrotrillina sp., Dendritina rangi, Meandropsina iranica, Meandropsina anahensis, Mean dropsina sp., Amphistegina sp., Globigerina spp. (Figures 12 and 13). These microfauna correspond to the Archaias asmricus -Archaias hensoni -Miogypsinoi des complanatus Assemblage Zone of Laursen et al . (2009) and Van Buchem et al . (2010), and to the SBZ 23 of Cahuzac and Poignant (1997), and indicate Chattian.

Figure 12 Photomicrograph of some selected fauna and other elements of the Asmari Formation in the studied sections. 1) Archaias hensoni (Smout and Eames, 1958), subaxial section, sample number K68; 2) Archaias sp., subequatorial section, sample number K68; 3) Archaias kirkukensis (Henson, 1950), subaxial section, sample number K68; 4) Miogypsina cf. globulina (Michelotti, 1841), subequatorial section, sample number K118; 5) Miogypsina cf. irregularis (Michelotti, 1841), axial section, sample number K145; 6) Ditrupa sp., sample number G367; 7) Favreina asmaricus (Elliott, 1962), oblique section, sample Number K120; 8) Austrotrillina howchini (Schlumberger, 1893), equatorial section, sample number H47; 9) Peneroplis sp., equatorial section, sample number H192; 10) Neorotalia viennoti (Greig, 1935), axial section, sample number G84; 11) Neorotalia viennoti (Greig, 1935), subequatorial section, sample number G60; 12) Dendritina rangi (d’Orbigny emend. Fornasini, 1904), subequatorial section, sample number K141; 13) Dendritina rangi (d’Orbigny emend. Fornasini, 1904), subaxial section, sample number K68.

Figure 13 Photomicrograph of some selected fauna of the Asmari Formation in the studied sections. 1) Bigenerina sp., Longitudinal section, sample number G288; 2) Elphidium sp.14, axial section, sample number G238; 3) Elphidium sp.1, equatorial section, sample number G274; 4) Peneroplis cf. farsensis (Henson, 1950), equatorial section, sample number K69; 5) Meandropsina iranica (Henson, 1950), subaxial section, sample number H276; 6) Meandropsina anahensis (Henson, 1950), oblique equatorial section, sample number G128; 7) Borelis melo curdica (Reichel, 1937), axial section, sample number K171.5; 8) Discorbis sp., axial section, sample number H186; 9) Miogypsinoides sp., axial section, sample number K282; 10) Triloculina trigonula (Lamarck, 1804), equatorial section, sample number K121; 11) Triloculina tricarinata (d’Orbigny, 1826), equatorial section, sample number G312; 12) Valvulinid sp., Longitudinal section, sample number H72; 13) Spiroloculina cf. cylindracea (Henson, 1950), equatorial section, sample number KS41; 14) Globigerina sp., axial section, sample number G396; 15) Globigerinoides trilobus (Reuss, 1850), axial section, sample number G367; 16) Globigerinoides subqudratus (Broennimann, 1954), axial section, sample number G386; 17) Globigerinoides diminutus (Bolli, 1957), axial section, sample number G414.
3.3. Assemblage 3
This assemblage occurred in all sections and is defined by the concurrence of Austrotrillina how chini, Austrotrillina asmariensis, Amphistegina sp., Aus trotrillina sp., Peneroplis farsensis, Peneroplis evolutus, Peneroplis thomasi, Peneroplis sp., Miogypsina irregularis, Miogypsina sp., Miogypsinoides sp., Elphidium sp.14, Elphidium sp.1, Dendritina rangi, Meandropsina iranica, Meandropsina sp., Favreina asmaricus (Figures 12 and 13) and represents the Miogypsina -Elphidium sp.14-Peneroplis farsensis Assemblage Zone of Laursen et al . (2009) and Van Buchem et al . (2010), and to the SBZ 24 of Cahuzac and Poignant (1997). This zone is restricted to Aquitanian.
3.4. Assemblage 4
This assemblage is recorded in all sections. The most important fauna are: Borelis melo curdica, Bore lis melo melo, Borelis sp., Meandropsina iranica, Me andropsina anahensis, Meandropsina sp., Austrotrillina sp., Dendritina rangi, Peneroplis farsensis, Peneroplis sp., Peneroplis evolutus, Elphidium sp.14, Elphidium sp.1, Miogypsina sp., Miogypsinoides sp., Miogypsinoides cf. dehaarti, Amphistegina sp., Operculina complanata, Operculina sp., Globigerina spp., Globigerinoides spp. (Figures 12 and 13). This zone is restricted to the Burdigalian based on the first appearance of Borelis melo curdica, Borelis melo melo (Cahuzac and Poignant, 1997; Laursen et al ., 2009; Van Buchem et al ., 2010). Although the presence of Miogypsi noides has been reported by Adams and Bourgeois (1967) in Burdigalian time, it is not reported in this time in the recent regional biostratigraphic study of the Asmari Formation introduced by Laursen et al . (2009) and Van Buchem et al . (2010). In this study, we have reported abundant occurrences of this genus (Miogypsinoides) in the study area during the Burdigalian.
3.5. Assemblage 5
This assemblage occurs from 344 to 415 m of the Asmari Formation, only in Gharibi Ha section, and consists of Globorotalia obesa, Globigerina praebulloides, Globigerinoides immaturus, Globigerinoides trilobus, Globigerinoides diminutus, Catapsydrax sp., Glo bigerinoides spp. (Figure 13). This assemblage overlies 78 m shallower carbonates of Burdigalian age. The top of this assemblage is marked by anhydrite deposits of the base of the Gachsaran Formation. Globigerina spp.-Turborotalia cerroazulensis -Hantkenina Assemblage Zone was introduced by Laursen et al . (2009) (Table 2). The age of this Assemblage Zone is Late Eocene-Early Oligocene. However, Wynd (1965) introduced Globigerina spp. Assemblage Zone (Table 1) from different intervals of the Asmari Formation in different localities in the Zagros Basin. The age of this zone was considered Late Eocene-Oligocene-Early Miocene based on its relationship to the underlying and overlying sediments. We recorded abundance planktonic foraminifera in Gharibi Ha section from above the Borelis melo curdica -Borelis melo melo Assemblage Zone. The age of this Assemblage Zone is considered Burdigalian based on its planktonic foraminifera (Iaccarino et al ., 2005) and underling Burdigalian shallow carbonate sediments.
4. Paleoecological interpretation
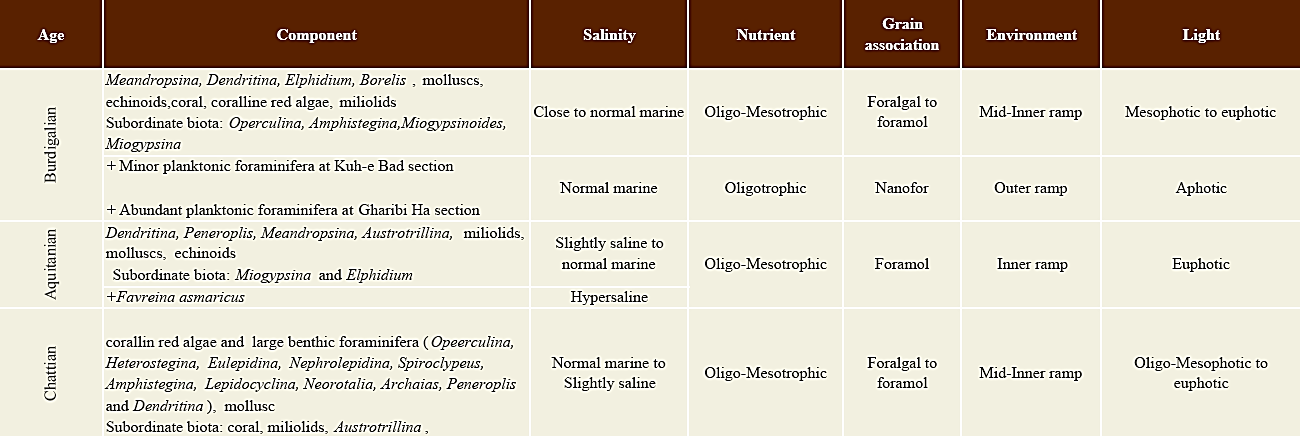
Paleoecology of Oligocene and Miocene carbonate platforms are studied by many authors with different points of view (e.g ., Hallock and Glenn, 1986; Hottinger, 1997; Pedley, 1998; Geel, 2000; Pomar, 2001; Romero et al ., 2002; Beavington-Penney and Racey, 2004; Cosovic et al ., 2004; Pomar et al ., 2004; Pomar and Hallock, 2008; Brandano et al ., 2009a, b; Bassi and Nebelsick, 2010; Flügel, 2010; Pomar et al ., 2012). In this study the most significant paleoecological elements such as salinity, light penetration, nutrients, temperature and water depth are discussed in detail.
4.1. Chattian
In the Chattian succession of the studied area the most significant and dominant biogenic components are coralline red algae; large, flat and perforate benthic foraminifera (Operculina, Heterostegina, Eulepidina, Nephrolepidina, Spiroclypeus, Amphistegina and Lepidocyclina); and imperforate foraminifera (Archaias, Peneroplis, Dendritina). Other skeletal grains consist of echinoid, gastropod, bryozoan and bivalve fragments (Table 4). Accordingly, the paleoecological conditions of the Asmari Formation during the Chattian could be interpreted as follows:
Table 4 Vertical distribution of faunal composition, paleoecological elements, grain association and depositional environment of the Asmari Formation in the study area.
Nummulitids, lepidocyclinids and red algae thrived in oligophotic to mesophotic mid-ramp environments (Kuh-e Bad and Gharibi Ha) preferably at water depths that ranged from 40 to 80 m (Bosence, 1983; Hottinger, 1997; Beavington-Penney and Racey, 2004).
Occurrence of large perforate benthic foraminifera (Eulepidina, Heterostegina, Neorotalia, Am phistegina), coralline red algae (Lithophyllum and Lithothamnion), imperforate foraminifera (Archaias, Peneroplis, Dendritina, Austrotrillina, miliolids) and molluscs (oysters, other bivalves, gastropods) in Kuh Shur, Kuh-e Bad and Gharibi Ha sections indicate normal to higher sea water salinity in inner ramp setting (Flügel, 2010; Allahkarampour Dill et al ., 2012) and could be consider as foralgal to foramol association (Wilson and Vecsei, 2005), (Figure 14) (Table 4).
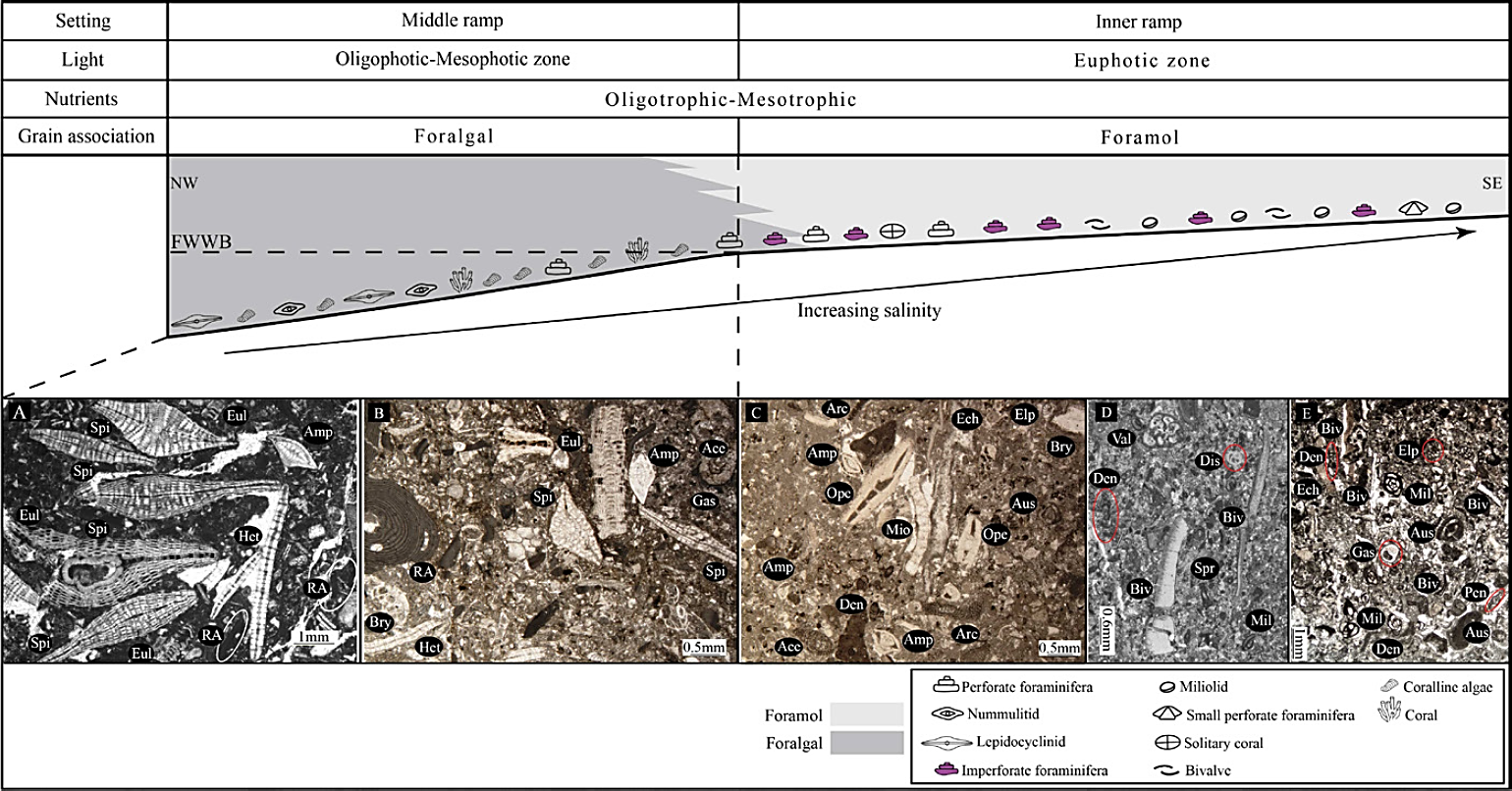
Figure 14 Sedimentary and paleoecological profile of the Asmari Formation during the Chattian time. A) floatstone to rudstone with abundant LBF (specially Spiroclypeus); B) packstone with abundant LBF and red-algal fragments; C) wackestone to packstone with perforate and imperforate benthic foraminifera; D) wackestone to packstone with high diversity of imperforate benthic foraminifera; E) packstone to grainstone with low diversity of imperforate foraminifera (mostly Dendritina), Amp: Amphistegina, Arc: Archaias, Aus: Austrotrillina, Biv: Bivalve, Bry: Bryozoan, Den: Dendritina, Dis: Discorbis, Ech: Echinoid fragment, Elp: Elphidium, Eul: Eulepidina, Gas: Gastropod, Het: Heterostegina, Mil: Miliolids, Ope: Operculina, RA. non-articulated red-algal fragments, Spi: Spiroclypeus, Spr: Spiroloculina, Val: Valvulinid.
The carbonates of the Asmari Formation in the study area were replaced by basinal marl and shale of the Pabdeh Formation towards the center of interashelf (toward the south). In the studied sections a major part of carbonate of the Asmari Formation (Chattian in age) in the Gharibi Ha and Kuh-e Bad sections is replaced by basinal marl and shale of the Pabdeh Formation in Halayjan section.
4.2. Aquitanian
The Aquitanian is characterized by abundant imperforate benthic foraminifera such as Dendritina, Peneroplis, Meandropsina, Austrotrillina and miliolids. Perforate foraminifera such as Miogypsina, Miogyp sinoides and Elphidium are also present as subordinate components. Mollusks and echinoids are also present. The Aquitanian biota of the studied area represents a foramol assemblage (Lees and Buller, 1972) (Figure 15) (Table 4). These foraminifera indicate a low energy and hypersaline condition and interpreted as a semi-restricted lagoon setting (inner ramp) (Wilson, 1975; Geel, 2000; Corda and Brandano, 2003; Bassi et al ., 2007; Flügel, 2010).
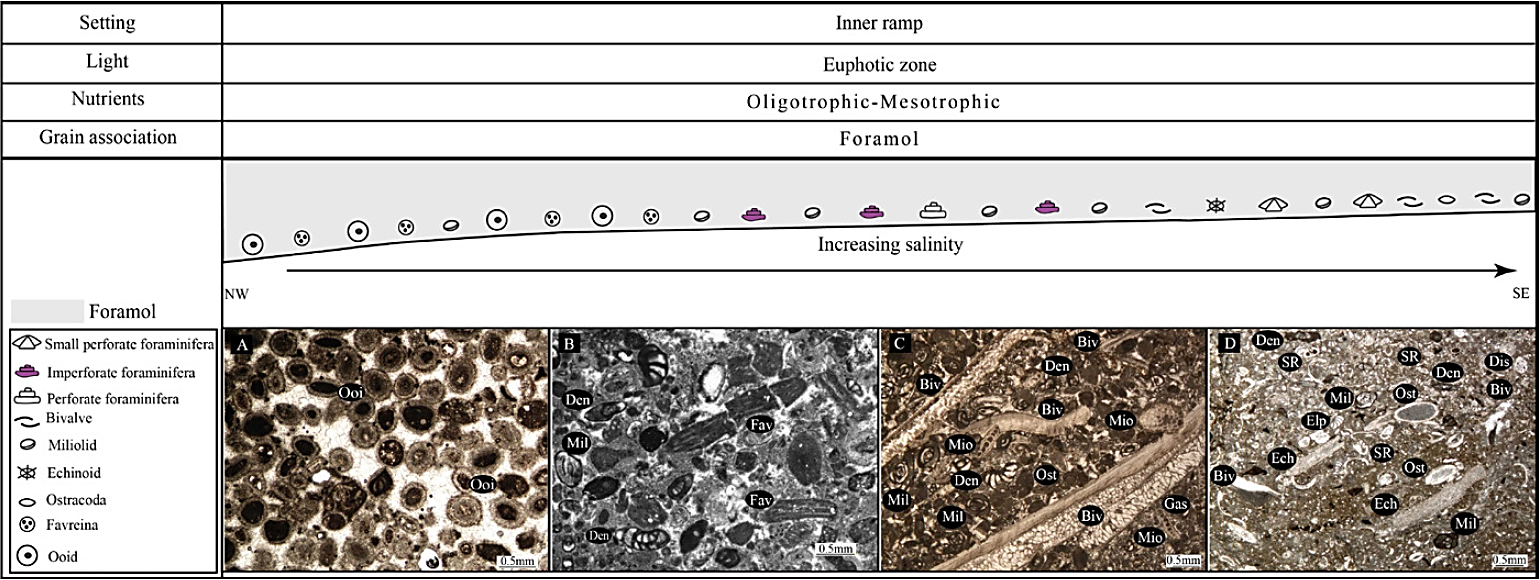
Figure 15 Sedimentary and paleoecological profile of the Asmari Formation during the Aquitanian time. A) Ooid grainstone; B) packstone to grainstone dominated with non-skeletal grains (Ooid and Favreina); C) packstone to grainstone with imperforate benthic foraminifera; D) wackestone to packstone with small benthic foraminifera and Echinoid fragments; Biv. Bivalve fragment; Den. Dendritina ; Dis: Discorbis, Ech. Echinoid fragment; Elp. Elphidium ; Fav. Favreina ; Gas. Gastropod; Mil. Miliolids; Mio: Miogypsinoides, Ooi. Ooid; Ost. Ostracod; Pen. Peneroplis ; SR. Small rotalids.
This biotic association indicates a deposition within the photic zone, in a sea-grass-dominated environment as suggested by the presence of epiphytic porcellaneous foraminifera such as Pen eroplis and miliolids (Wilson, 1975; Langer, 1993; Beavington-Penney and Racey, 2004; Reich et al ., 2015). Further evidence for sea grass can be given by the poorly sorted and micrite-rich textures and absence of sedimentary structures. The associated fauna (Peneroplis, Borelis, discorbids and thick samples of Amphistegina) are also typical for seagrass meadows (Murray, 1973; Brasier, 1975; Beavington-Penney and Racey, 2004; Brandano et al ., 2009a; Reuter et al ., 2010; Pomar et al ., 2014).
One of the most important and excellent markers for the Aquitanian is Favreina asmaricus (coprolitic facies) (Figure 15) (Table 4) with occurrences related to the particular environmental condition (isolated hypersaline basins for part of the time) (Laursen et al ., 2009; Van Buchem et al ., 2010). In the more saline interval Favreina asmaricus is common (Mossadegh et al ., 2009). The presence of porcellaneous benthic foraminifera with Favreina asmaricus, coralline algae, coral, bryozoa and echinoderm fragments are common in lower parts of the Aquitanian sediments, especially in Halayjan section. Favreina asmaricus is rare in Kuh Shur, Kuh-e Bad and Gharibi Ha sections which are situated in shallower parts of the Asmari intrashelf basin.
The Favreina limestone is a well known facies of intrashelf margin of the Aquitanian. It has been interpreted as restricted condition of interashelf basin (Van Buchem et al ., 2010).
4.3. Burdigalian
In the study area, the common biota of the Burdigalian are coral, coralline algae, Amphistegina, Miogypsinoides, Meandropsina, Elphidium, Borelis, Dendritina, Peneroplis, miliolids and mollusc debris (Table 4). A similar assemblage was reported from the inner ramp of the Miocene sediments of the Central Apennines (Corda and Brandano, 2003) and from the Early Oligocene deposits of the Lower Inn Valley (Nebelsick et al ., 2001). The abundance of larger benthic foraminifera (Borelis, Peneroplis, Dendritina, Heterostegina, Operculina) and zooxanthellate corals indicate mesophothic to euphotic condition (Halfar et al ., 2004; Pomar et al ., 2004; Pomar and Hallock, 2008; Brandano et al ., 2009b; Mazzucchi and Tomassetti, 2011; Tomassetti and Brandano, 2013). During the Burdigalian a slightly restricted lagoon became established over the entire width of the platform in the SW (e.g. Dezful Embayment) characterized by proliferation of imperforate foraminifera (Rahmani et al ., 2009).
At Kuh-e Bad section, planktonic foraminifera (rare), coral, corallinacean algae, Heterostegina and Operculina are also present in the Burdigalian assemblage (Table 4). Towards the east of studied area (Figures 1C and 10) in Gharibi Ha section, planktonic foraminifera facies is more common in the upper part of the Burdigalian. These faunal associations represent a nannofor assemblage (Lees and Buller, 1972) (Table 4). The nannofor (aphotic zone-outer shelf) association occurred in a deeper water depositional setting. The existence of abundant planktonic foraminifera indicates that the sediment of the upper part of the Burdigalian was deposited in deeper-water environment in Gharibi Ha section (Figures 16 and 17).

Figure 16 Sedimentary and paleoecological profile of the Asmari Formation during the Burdigalian time. A) wackestone with small benthic foraminifera and Echinoid fragments; B) wackestone to packstone with high diversity of imperforate foraminifera; C) packstone with perforate benthic foraminifera and red-algal fragments; D) packstone dominated with planktonic foraminifera and Echinoid fragments; Biv: Bivalve fragment, Bor: Borelis, Bry: Bryozoan, Den: Dendritina, Ech: Echinoid fragment, Gas: Gastropod, Glo: Globigerinids, Mea: Meandropsina, Mil: Miliolids, Mio: Miogypsinoides, Ope: Operculina, RA: non-articulated red-algal fragments, SR: Small rotalids.
5. Discussion
Our detailed analysis of the stratigraphy and paleoecology show that the Asmari Formation was deposited on a carbonate ramp in tropical to subtropical latitudes (10° to 30°) (Vaziri-Moghaddam et al ., 2006; Heydari, 2008; Allahkarampour Dill et al ., 2010; Vaziri-Moghaddam et al ., 2010; Shabafrooz et al ., 2015a, b). The differentiation between non-tropical and tropical carbonates is mainly based on analysis of skeletal components (Lees and Buller, 1972; Carannante et al ., 1988; Nelson, 1988). The foraminiferal associations such as Dendritina, Archaias, Borelis, Spiroclypeus, Lepidocyc lina, Neorotalia, and Amphistegina within the Asmari Formation suggest that carbonate sedimentation took place in tropical to subtropical latitudes. The symbiont bearing larger foraminifera are known to thrive in tropical waters (Langer and Hottinger, 2000). According to Heydari (2008) the Zagros Basin was located close to 30° N during the Oligocene-Miocene. This idea suggests that carbonate sedimentation of the Asmari Formation took place in tropical to subtropical waters.
Zooxanthellate corals and large benthic foraminifera are the most successful shallow, benthic carbonate producers in clear, tropical-subtropical oceanic waters (Hallock, 2001). Zooxanthellate corals and the LBF (Borelis, Archaias, Peneroplis, Lep idocyclina) assemblages of the Asmari Formation thrive in oligotrophic to possibly slightly mesotrophic waters (Langer and Hottinger, 2000; Halfar et al ., 2004; Pomar et al ., 2004; Bassi and Nebelsick, 2010).
Oligotrophic to mesotrophic conditions in the inner ramp setting (Chattian-Burdigalian) lead to the occurrence of larger imperforate foraminifera such as Archaias, Borelis, Peneroplis and Austrotrillina . These porcellaneous larger foraminifera show well lit, highly translucent, shallowest part of the photic zone. Lens shaped rotalids lived in mesophotic conditions in the middle ramp setting (Chattian), and large and flat lepidocyclinid-nummulitids characterizes the sediments of the deeper oligophotic zone in the middle ramp setting (Chattian).
These assemblages of benthic foraminifera are not identified in the Kuh Shur and Halayjan sections, because of their deep-seated paleogeographical position during the Chattian (Figure 10). However, the base of the Asmari Formation deposits, from southeast to northwest, becomes younger and diachronous. Consequently, this stepwise younging could be expected in series with basinward progradation of the Asmari Formation over the basinal shale and marl of the Pabdeh Formation. Some of these progradational architectures (e.g . Eshgar and Gurguda clinoforms) have been documented in different parts of the Asmari depositional basin (Van Buchem et al ., 2010; Shabafrooz et al ., 2015b).
During the Chattian the thickness of the Asmari Formation increases from northwest to southeast in the study area. Afterward, during the Aquitanian, the thickest interval of the formation is recorded in the Halayjan section. In the Burdigalian the thickness of the formation increases from the Halayjan section to the Kuh-e Bad and decreases towards the Gharibi Ha section (Figure 10).
The upper part of the Asmari Formation in the Gharibi Ha section (Burdigalian) is characterized by abundance of planktonic Foraminifera. This suggests that deeper-water carbonate (nannofor, oligophotic to aphotic, outer ramp) deposition was active in the Asmari Formation, in the Gharibi Ha section. The planktonic foraminifera documented within the upper part of the Asmari Formation in the Gharibi Ha section also indicate a deposition in the tropical waters under oligotrophic conditions as suggested by the occurrence of Globigerinoi des trilobus which proliferate in warm nutrient-poor waters (Tolderlund et al ., 1971; Be, 1977; Hemleben et al ., 1989). In the light of modern analogues, the occurrence of larger benthic foraminifera and zooxanthellate corals in the geological record is interpreted as photozoan assemblages. Nevertheless, the Asmari Formation is dominated by heterozoan assemblages. Heterozoan carbonates can develop on a carbonate ramp (Carannante et al ., 1988; Jones and Desrochers, 1992; Boreen and James, 1995; Feary, 1995; Brachert et al ., 1998). The spread of heterozoan assemblages related to the low capacity of corals to thrive in high-light conditions and to form a wave-resistant reef promoted the distribution of heterozoan assemblages ( Brandano et al ., 2009a; Rahmani et al ., 2009; Allahkarampour Dill et al ., 2010).
Oligo-Miocene carbonates are extensive throughout SW Iran. The Dill Anticline section (Allahkarampour Dill et al ., 2010, 2012) and the Gachsaran and Bibi-Hakimeh oilfields (Shabafrooz et al ., 2015b) were selected for comparison with the studied sections. The biotic associations of the mentioned areas are quite similar to those described in the study area. They were deposited on a carbonate ramp dominated by heterozoan skeletal assemblages, in which most benthic carbonate production occurred in inner-to-middle ramp settings. The main producers were larger benthic foraminifera and red algae, but zooxanthellate corals are rare (Allahkarampour Dill et al ., 2010, 2012; Shabafrooz et al ., 2015b). This biotic association suggests that carbonate sedimentation of the Asmari Formation took place in tropical-subtropical waters under oligotrophic-mesotrophic conditions.
The other examples of the Oligocene shallow-water carbonates have been described in Malta (Brandano et al ., 2009a, b) and in northeastern Italy Lessini Shelf Venetian foreland basin (Bassi and Nebelsick, 2010). At these areas rotalids, miliolids and imperforate foraminifera are frequent in the inner ramp, while red algae and lepidocyclinids are abundant in the middle ramp. The foraminiferal assemblage of the Asmari Formation shows close affinities to that described by Brandano et al . (2009a) of the Malta carbonate platform. Such assemblages suggest that carbonate sedimentation took place in tropical to sub-tropical waters, under oligotrophic to slightly mesotrophic conditions. In the carbonate platform of the Attard Member (Brandano et al ., 2009a, b) and Asmari Formation, zooxanthellate corals did not form a reef framework.
As a result, the lateral facies distribution within the Asmari succession suggests that the sea level in the Chattian was relatively shallow, but deepened toward the NW region. These observations indicate that the Chattian to Aquitanian was a time of relative tectonic quiescence in this area. The lateral distribution of facies during the Late Burdigalian suggests a significant depth range, from the shallowest facies in the NW region to the most distal facies in the SE region. This pattern contrasts with subtle lateral shifts in facies characteristic of the underlying sequence, suggesting increased subsidence during this time in the SE regions (Figure 17). Previous studies (Seyrafian, 2000; Van Buchem et al ., 2010) mentioned that in the north Izeh area, the shallow water deposits of the Early Burdigalian are locally overlain by pelagic marls of the Late Burdigalian (e.g . Ghaleh Madreseh section), while in the Chidan section (Figure 1C), coral, corallinacean algae, benthic imperforate and perforate foraminifera were deposited (Van Buchem et al ., 2010). The deepening in the north is considered a result of a regional tilting that started in the Late Burdigalian (Van Buchem et al ., 2010) (Figure 17).
6. Conclusion
In this research based on the micropaleontological analysis of the larger benthic foraminifera and their distribution, five assemblage zones have been recorded. Three assemblages (1, 3, 4) are present in all sections (Halayjan, Kuh Shur, Kuh-e Bad and Gharibi Ha). Assemblage 2 is recorded in two sections (Kuh-e bad and Gharibi Ha) and assemblage 5 is only occurred in Gharibi Ha section. Assemblages 1 and 2 indicate the Chattian age; assemblage 3 is restricted to the Aquitanian age; and assemblages 4 and 5 suggest the Burdigalian age. According to this study, the age of the Asmari Formation ranges from the Oligocene (Chattian) to Early Miocene (Aquitanian-Burdigalian).
The biotic associations within the Asmari Formation indicate that the lower part of the Asmari Formation (Chattian) was mostly deposited in the oligophotic-mesophotic zone, in normal saline waters. The middle and upper parts of the Asmari Formation is characterized by euphotic-mesophotic biota (except for the Gharibi Ha section, SE of study area, where the sedimentation occur in aphotic situation in Burdigalian). The oligophotic zone is characterized by large and flat lepidocyclinids and nummulitids and corallinacean algae. Lens shaped rotalids abundantly thrived in the mesophotic zone. The euphotic zone is dominated by porcellaneous larger foraminifera and indicates well lit, highly translucent, shallowest part of the photic zone.
The biotic assemblages recognized within the Asmari Formation indicate a deposition in tropical to sub-tropical waters, as suggested by the presence of diverse large benthic foraminifera. Carbonate grain associations suggest a heterozoan assemblage for the Asmari Formation. Deposition of the Asmari Formation took place under oligotrophic to mesotrophic conditions in the study area.
From the Chattian to Aquitanian was a time of relative tectonic stability in the area. The deepening in the north of the study area is considered as a result of a regional tilting that started in the Late Burdigalian.

