











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introducción
La familia Albanerpetontidae Fox y Naylor, 1982, representa la única familia de Allocaudata. Este grupo extinto, junto con Anura, Caudata y Gymnophiona, conforman el clado Lissamphibia, el cual tiene un alcance temporal del Triásico temprano al reciente. El alcance temporal de los albanerpetóntidos va desde el Jurásico medio hasta el Plioceno tardío con una distribución espacial en Laurasia y el norte de áfrica (Gardner y DeMar, 2013; Maddin et al., 2013). Los miembros de la familia Albanerpetontidae se caracterizan por presentar una pequeña talla corporal, similar al de las salamandras, y poseer los siguientes caracteres óseos: frontales fusionados; articulación intermandibular interdigitada; dientes marginales no pedicelados, con coronas en forma de cincel y generalmente tricúspides; y una vértebra "cervical" modificada que recuerda al complejo atlas-axis de mamíferos (Estes y Hoffstetter, 1976; Fox y Naylor, 1982; McGowan y Evans, 1995; Gardner, 2000a, 2001; Ruta y Coates, 2007). La mayor parte del registro de Albanerpetontidae consiste en huesos aislados, representados principalmente por premaxilares, maxilas, dentarios y frontales (Gardner, 2000a, 2000b; Gardner y Böhme, 2008). Estos elementos óseos presentan características diagnósticas que ayudan a una identificación a nivel genérico o inclusive específico (Gardner, 2000a). Cabe destacar que recientemente se encontró y describió un neurocráneo fósil referido a Albanerpeton pannonicum (MáFI V.11.123.1) (Maddin et al ., 2013).
A la fecha se han reconocido cuatro géneros de albanerpetóntidos: Albanerpeton (Estes y Hoffstetter, 1976), con distribución euroamericana; Anoualerpeton (Gardner et al., 2003), registrado para Europa y áfrica; Celtedens (McGowan y Evans, 1995) y Wesserpeton (Sweetman y Gardner, 2013) han sido registrados en Europa. Sin embargo, existen varios especímenes que no han sido nombrados a nivel genérico/específico y han sido registrados en diferentes partes de Laurasia y el norte de áfrica (Gardner, 2002; Gardner et al ., 2003; Sweetman y Gardner, 2013). El género mejor conocido es Albanerpeton el cual incluye siete especies descritas a la fecha (Gardner y Böhme, 2008). Se ha sugerido que este género se originó durante el Aptiano/Albiano temprano de Norte América y que sobrevivió hasta el Plioceno tardío en Europa (Gardner y Böhme, 2008).
Durante el Cretácico y el Paleoceno la diversidad de Albanerpeton fue moderada en Norte América (Gardner y DeMar, 2013), su evolución está relacionada a las planicies costeras que bordearon el margen occidental del Mar Interior del Oeste. Casi todas las localidades donde este género ha sido reportado se encuentran en el lado oriental de Laramida, repartidos en las provincias de Alberta y Saskatchewan (Canadá) y los estados de Montana, Dakota, Wyoming, Utah, Colorado, Nuevo México, Oklahoma y Texas (Estados Unidos), con excepción de dos: Marshalltown (en Nueva Jersey) y Bladen (en Carolina del Norte) que se encuentran en la costa este de Estados Unidos (Gardner y DeMar, 2013).
En México, el registro de Lissamphibia en el Cretácico tardío es prácticamente inexistente. Estes y Sanchíz (1982) mencionan la presencia de Scotiophryne putulosa en Baja California, reporte que ha sido puesto en duda por Gardner y DeMar (2013) al no estar figurado y no tener acceso al material. También existen reportes de restos de anuros indeterminados colectados en afloramientos de la formación El Gallo, Baja California (Lillegraven, 1972, 1976; Estes y Sanchíz, 1982) y en la Formación Cerro del Pueblo, Coahuila (Aguillón-Martínez, 2010).
Durante las décadas de los 60's y 70's se realizaron expediciones paleontológicas por parte del personal del Museo de Historia Natural del Condado de Los ángeles, California, junto con miembros de la Universidad de California y del Instituto de Geología (UNAM), México, en el área de El Rosario, Baja California. Como resultado se obtuvo una colección de restos pertenecientes a numerosos grupos de vertebrados (principalmente de dinosaurios, pero también de anfibios, tortugas, lagartos, cocodrilos, aves y mamíferos) recolectados en depósitos continentales de edad cretácica tardía de las formaciones El Gallo y La Bocana Roja (Morris, 1973; Molnar, 1974; Lillegraven, 1972, 1976; Morris, 1981; Nydam, 1999; entre otros).
Desde 2004 se retomaron los trabajos de campo en el área de El Rosario enfatizando la prospección de nuevas localidades y colecta de microvertebrados, bajo la dirección de Gregory Wilson (University of Washington) y Marisol Montellano (UNAM). Como resultado se han localizado nuevos sitios y recolectado numerosos microvertebrados. En uno de estos sitios, denominado ROS51, se recolectaron los restos de Albanerpetontidae, descritos en este trabajo. Siendo el primer reporte formal del grupo para México y extendiendo su registro geográfico hacia el sur, como era lo esperado.
2. Área de estudio
La localidad ROS51 se encuentra dentro del área de El Rosario, al suroeste de la población Rosario de Arriba, Baja California (Figura 1). Sus coordenadas geográficas son: 30° 03' 12.5'' N, 115° 45' 34.2'' W. En esta área afloran sedimentos continentales, compuestos por conglomerados, arenisca, limolitas, lutitas y tobas que conforman la formación El Gallo. Esta formación está dividida en tres miembros: El Disecado, La Escarpa y El Castillo (Kilmer, 1963). El material proviene del miembro El Disecado, el cual fue interpretado como un sistema fluvial meándrico (Schile, 1974). Se obtuvieron edades radiométricas con 40Ar/39Ar de tobas del miembro El Disecado que oscilan entre 74.87 ± 0.5 Ma a 7.59 ± 0.09 Ma, las cuales corresponden al Campaniano tardío (Renne et al., 1991).
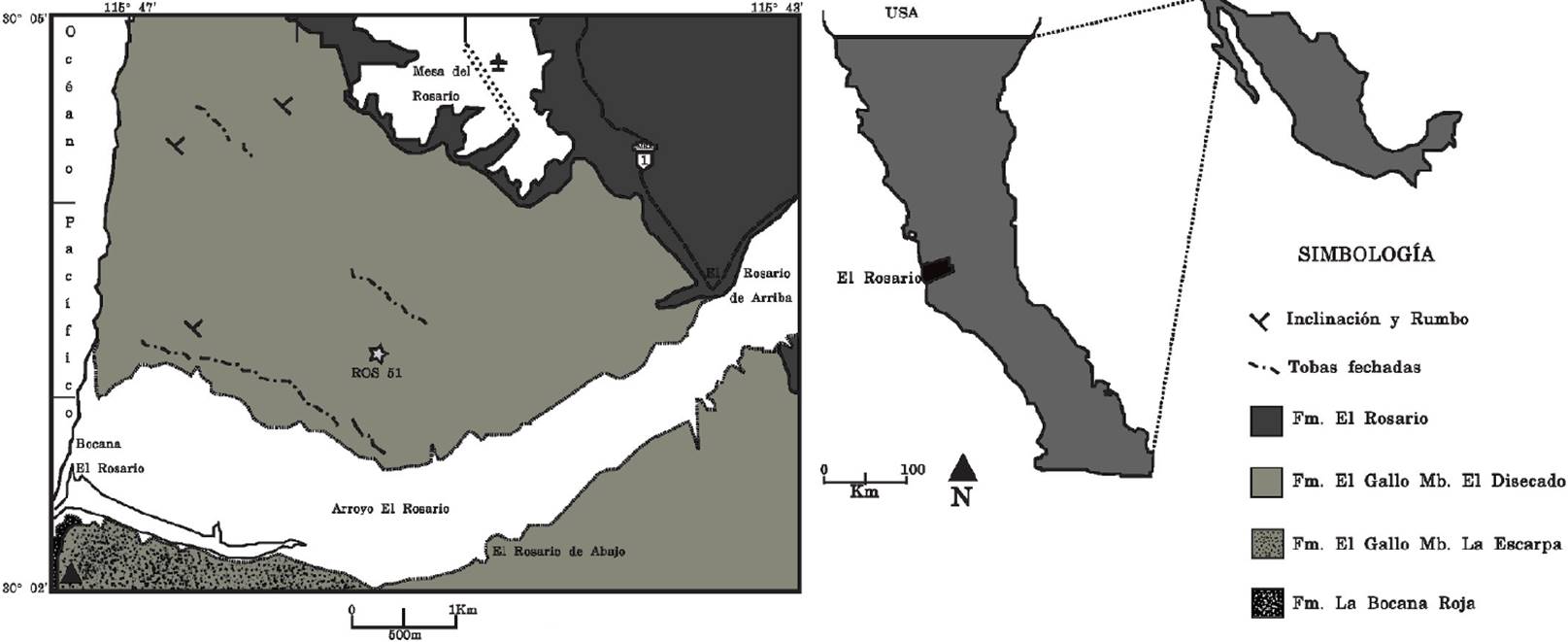
Figura 1 Localización de El Rosario, Baja California, donde aflora la formación El Gallo (modificado de Kilmer, 1963) y mapa geológico del área del Rosario con la ubicación del sitio ROS51 (datos geológicos tomados de Fulbord y Busby 1993).
La sección estratigráfica de la localidad ROS51 está compuesta por estratos de arenisca amarilla de grano medio a grueso con estratificación cruzada y/o horizontal, intercalados con areniscas grisáceas de grano fino y matriz de arcilla, y por limolitas arenosas grisáceas. El material fue hallado en un estrato conformado por arenisca arcillosa de grano fino.
3. Material y metodología
Los especímenes fósiles incluyen un fragmento de un dentario izquierdo con tres dientes incompletos (IGM 4859), y un fragmento de un premaxilar derecho con dos dientes casi completos (IGM 4860) (Figuras 2 y 3). Fueron colectados con el uso de microscopio del sedimento previamente tamizado. Para su descripción se siguió la terminología sugerida por Gardner (2000a).

Figura 2 Albanerpetontidae indet. IGM 4860, fragmento de premaxilar derecho en vista labial (A y B) y vista lingual (C y D). Abreviaturas: pde., pars dentalis .
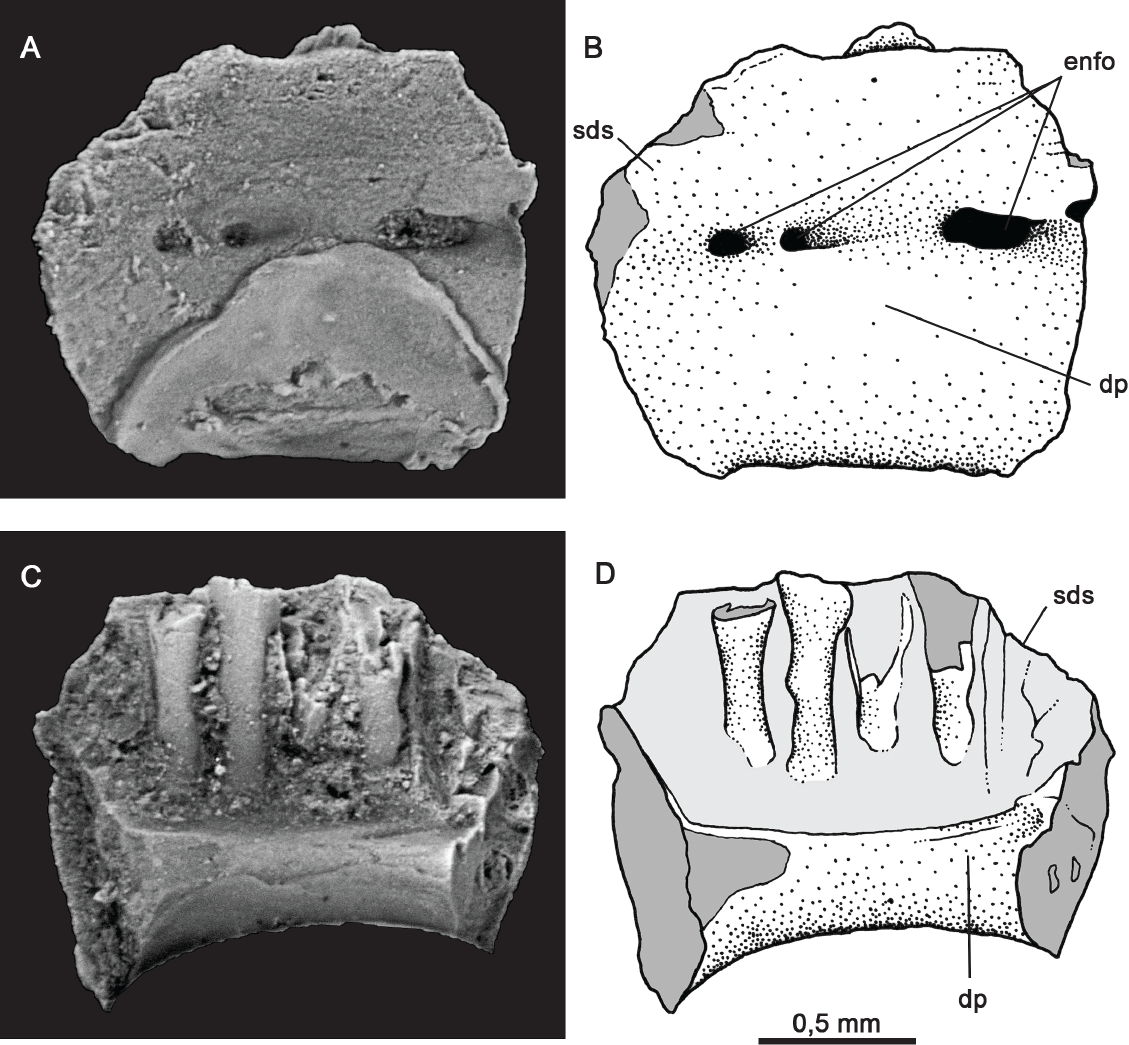
Figura 3 Albanerpetontidae indet. IGM 4859, fragmento de dentario izquierdo en vista labial (A y B) y vista lingual (C y D). Abreviaturas: dp., parapeto dental; enfo., forámenes nutritivos; sds., placa subdental. enfo., forámenes nutritivos.
3.1. Abreviaturas
IGM, acrónimo de la Colección Nacional de Paleontología, Museo María del Carmen Perrilliat, Instituto de Geología, UNAM, México.
Fm. Formación.
FMNH PR, Field Museum of Natural History, Chicago, Illinois, Estados Unidos de América.
MAFI, Magyar AllamiFoldtaniIntezet (Instituto Geológico de Hungría), Budapest, Hungría.
4. Paleontología sistemática
Lissamphibia Haeckel, 1866
Allocaudata Fox y Naylor, 1982
Albanerpetontidae Fox y Naylor, 1982
Género y especie indeterminados
4.1. Descripción
Premaxilar. El ejemplar IGM 4860 es un fragmento de un premaxilar derecho (Figura 2) que corresponde a la porción distal del pars dentalis . En el lado labial se observa la faceta donde se aloja el "flange" de la maxila. Conserva dos dientes pleurodontos no pedicelados y se observa el espacio que debió estar ocupando un tercer diente. Los dientes están casi completos, carecen de las coronas, son robustos y lisos, se encuentran muy próximos entre sí, tal como se observa en los albanerpetóntidos. En la cara labial del premaxilar aparentemente se presenta un foramen.
Dentario. El ejemplar IGM 4859 (Figura 3) corresponde a un fragmento de dentario izquierdo, probablemente es la porción entre la sínfisis y la apertura del canal de Meckel. Conserva tres dientes que carecen de corona, están rotos a la altura del parapeto dental, también se observan tres espacios donde hubieron dientes. Los dientes son pleurodontos y están cercanos uno de otro. Estos se encuentran comprimidos mesiodistalmente. El parapeto dental es moderadamente alto. El borde dorsal en vista labial es horizontal.
La placa subdental es robusta y en forma de canal, en su porción más distal está rota y se observa una apertura amplia para el canal inferior de Meckel. En la terminación mesial se observan dos pequeñas aperturas, una encima de la otra, lo que podría estar indicando un canal meckeliano bifurcado.
En vista labial la superficie del dentario es lisa, salvo por la presencia de dos o tres forámenes nutritivos.
5. Discusión
Durante el Cretácico tardío en Norteamérica, además de los registros de la familia Albanerpetontidae, existen diversos registros de familias de salamandras, teniendo representados los siguientes grupos: sirénidos (Habrosaurus ); anfiúmidos (Proamphiuma ); proteidos (Paranecturus ); batrachosauroideos (Opisthotriton , Parrisia y Prodesmodon ) y scapherpetóntidos (Scapherpeton , Lisserpeton y Piceoerpeton ) (e.g. Estes, 1981; Holman, 2006; Gardner, 2012; DeMar, 2013; Gardner y DeMar, 2013).
Tanto la historia taxonómica como la posición filogenética de las salamandras extintas son complicadas y controversiales. Comenzando porque la mayor parte de los holotipos son vértebras (v.g . FMNH PR 391, holotipo de Prosirenelinorae Goin y Auffenberg, 1958, representado por un conjunto de vértebras dorsales), así como algunas de las formas identificadas se han considerado como neoténicas (v.g. Batrachosauroidae). Inclusive, de manera inicial Albanerpeton fue considerado como una salamandra por Estes y Hoffstetter (1976) y fue incluido dentro de los Prosirenidae, pues compartía caracteres con anfibios pequeños no saltatoriales. Más tarde, Fox y Naylor (1982) reconocieron que este taxón posee caracteres únicos con lo que erigieron la familia Albanerpetontidae y el orden Allocaudata; algo cuestionado por Trueb y Cloutier (1991), Estes y Sanchíz (1982) y apoyado por Milner (1994). Uno de los caracteres que diferencia a los Allocaudata de los Caudata, es la dentición; en los albanerpetóntidos está caracterizada por presentar dientes no-pedicelados generalmente tricúspides con forma de cincel; en cambio los dientes en las salamandras son pedicelados y bicúspides, salvo en grupos neoténicos donde éstos son no-pedicelados y monocúspides (Skutschas, 2013).
Por el estado de preservación del material aquí descrito (IGM 4859 e IGM 4860) la identificación a un nivel taxonómico de género o especie es difícil; sin embargo, los ejemplares presentan características morfológicas que permiten su identificación para incluirlos dentro de la familia Albanerpetontidae. Estas características son: dientes pleurodontos no pedicelados, elongados pero robustos y comprimidos mesiodistalmente, que se encuentran muy próximos entre sí. La presencia de forámenes nutritivos en la superficie labial del dentario también es una característica que de acuerdo a Gardner (2000a, 2000b) y Gardner y Böhme (2008) es diagnóstica para los miembros de esta familia.
Descartamos que los ejemplares IGM 4860 e IGM 4859 pertenezcan a algún grupo de salamandras neoténicas por los siguientes motivos: las dos familias que ocurren durante el Cretácico en Norte América son Prosirenidae y Batrachosauroidae. Los holotipos de la familia Prosirenidae, en Norte América, son vértebras, y las mandíbulas inicialmente referidas a esta familia, posteriormente fueron designadas como Albanerpeton (Fox y Naylor, 1982). Con respecto a la familia Batrachosauroidae, existen tres géneros presentes en el Cretácico de Norte América, Batrachosauroides , Opisthotriton y Parrisia , de estos tres Batrachosauroides y Opisthotriton son considerados por Bonett et al. (2013) formas gigantes y grandes respectivamente y son de mayor talla con respecto las formas de albanerpetóntidos conocidas para Norte América; el género Parrisia es de un tamaño similar al de otros albanerpetóntidos conocidos; sin embargo, los miembros de la familia Batrachosauroidae presentan varios "pits" en la base de los dientes, carácter no presente en la familia Albanerpetontidae (observación personal).
Lillegraven (1972, 1976) ya hacía mención de la presencia de anfibios en la formación El Gallo, Baja California. Por su parte, Estes y Sanchíz (1982) citan que Clemens et al. (1979) hicieron mención de la presencia de un cráneo y parte del postcráneo del anuro Scotiophryme en esta formación. Sin embargo, Gardner y DeMar (2013) lo cuestionan ya que los restos no fueron ni figurados ni descritos, y por lo tanto no se puede verificar la identificación. En la localidad de La Parrita, Formación Cerro del Pueblo, Coahuila, Aguillón-Martínez (2010) mencionó la presencia de elementos esqueléticos de anuros sin llegar a ninguna identificación más específica. De esta forma dentro de los registros existentes de Lissamphibia para México, los especímenes aquí descritos constituyen el primer registro formal de la familia Albanerpetontidae para el país.
La distribución geográfica de la familia en Norte América durante el Cretácico Tardío va desde la provincia de Alberta (en la Formación St. Mary River, Gardner et al., 2003) a Texas (Formación Aguja, Rowe et al., 1992) (Figura 4). Por lo tanto, este registro extiende la distribución geográfica de la familia hacia el suroeste de Norte América.
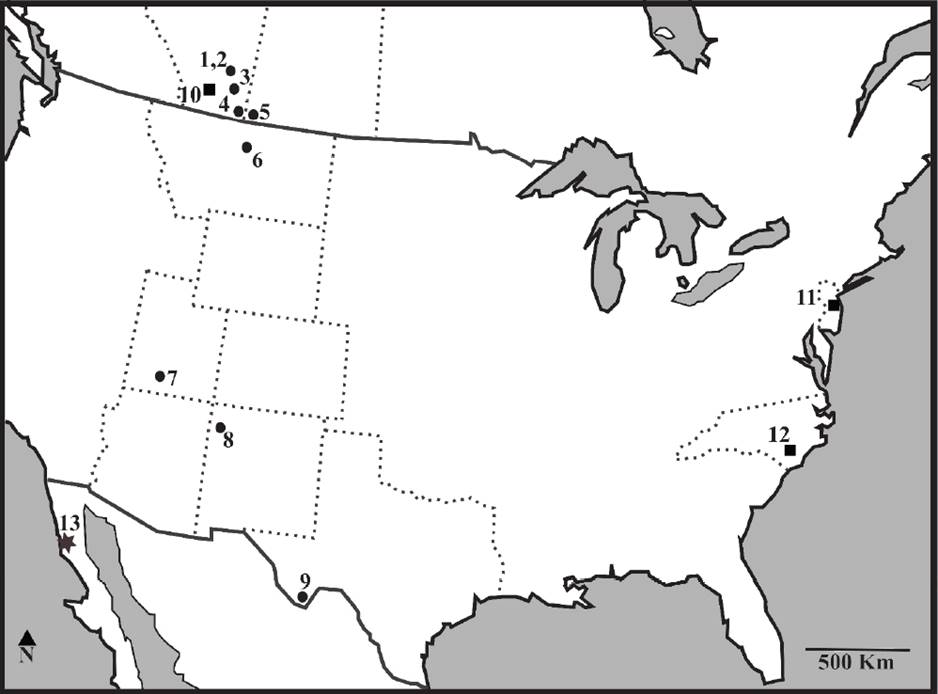
Figura 4 Distribución de la familia Albanerpetontidae en Norte América durante el Campaniano. Mapa modificado de Gardner y DeMar, 2013: 1. Fm. Dinosaur Park (Alberta); 2. Parte superior de la Fm. Oldman (Alberta); 3. Parte superior de la Fm. Foremost (Alberta); 4. Fm. Oldman y Fm. Foremost (Alberta); 5. Fm. Oldman o Dinosaur National Park (Saskatchewan); 6. Fm. Judith River (Montana); 7. Fm. Kaiparowits, (Utah); 8. Fm. Fruitland (Nuevo México); 9. Fm. Aguja (Texas); 10. Fm. St. Mary River (Alberta); 11. Fm. Marshalltown (Nueva Jersey); 12. Fm. Bladen (Carolina del Norte); 13. Fm. El Gallo, con presencia de Albanerpetontidae indet. (Estrella); Círculo: presencia del género Albanerpeton ; Cuadrado: presencia de Albanerpetontidae indet.
Este registro es una contribución al conocimiento de la fauna de vertebrados cretácicos de Norteamérica, en particular a lo que se refiere a anfibios de México. Se propone continuar la búsqueda y recolecta de microvertebrados, lo cual permitirá un mejor entendimiento de la diversidad y evolución de las faunas cretácicas en Norte América.

