











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introducción
Simojovel de Allende, Chiapas, constituye uno de los depósitos de ámbar más importantes del Cenozoico (Solórzano-Kraemer, 2007, 2010). En él se han conservado de manera excepcional organismos completos o fragmentos de ellos, como las plantas, cuyo registro fósil es relativamente más escaso en la zona, en comparación con otros grupos biológicos, como el de los insectos (Avendaño-Gil et al., 2012). Dentro del registro de plantas fósiles el grupo de las angiospermas es el mejor representado, en su mayoría por flores. éstas constituyen un caso especial, ya que se tratan de órganos delicados y efímeros, con un alto potencial sistemático, al ser fácilmente observables (Bhattacharya, 2005) y más consistentes en sus caracteres, en comparación con órganos vegetativos que al ser sometidos a la presión del medio ambiente fluctuante (Sivarajan, 1991).
Las flores fósiles reportadas en los depósitos de ámbar de Simojovel de Allende se han identificado como miembros de distintas familias. Por ejemplo: Fabaceae con Hymenaeae mexicana (Poinar y Brown, 2002) y H. allendis (Calvillo-Canadell et al., 2010), reconocidas hasta ahora como las dos especies extintas que dieron origen al ámbar; además de Anacardiaceae representada por Tapirira durhamii (Miranda, 1963), Arecaceae por Socratea brownii (Poinar, 2002), Colpothrinax chiapensis (Chambers et al., 2012) y Meliaceae por Swietenia miocenica (Castañeda-Posadas y Cevallos-Ferriz, 2007). En este trabajo se amplía el registro taxonómico de las angiospermas con una nueva especie extinta de Salicaceae sensu lato (s.l. ) perteneciente al género Lunania .
Salicaceae sensu stricto (s.s. ) está constituida por dos géneros Salix y Populus , sin embargo, la actual circunscripción de la familia propuesta por Chase et al. (2002) a través de un análisis filogenético con datos moleculares (rbcL) de Flacourtiaceae, sugiere que la mayoría de los géneros incluidos en Flacourtiaceae forman parte de Salicaceae, ampliando la circunscripción de la familia a ca . 54 - 55 géneros y dividiéndola en nueve tribus: Saliceae, Flacourtieae, Scyphostegieae, Homalieae, Scolopieae, Prockieae, Abatieae, Bembicieae, y Samydeae. Esta última es considerada por algunos taxónomos como una familia (Samydaceae), dejando al resto como Salicaceae sensu medio , pero este enfoque no ha sido ampliamente aceptado (Alford y Belyaeva, 2009).
La tribu Samydeae se encuentra constituida por 12 géneros, incluido Lunania Hook (Chase et al., 2002). Lunania es un género neotropical, constituido por ca . 14 especies (Hooker, 1844; Robyns, 1968; Chase et al ., 2002) de árboles y arbustos, con flores pequeñas, bisexuales, en racimos axilares o terminales y sostenidas por dos bractéolas. Sin embargo, las bases de datos recientes como The Plant List (2013) sólo reconoce a tres especies: L. parviflora , L. mexicana y L. bauchii . Este trabajo representa al primer registro fósil del género y está basado en una flor.
2. Materiales y métodos
2.1. Localidad
El material fósil fue colectado en el municipio de Simojovel de Allende en el Estado de Chiapas (17°08ʼ19"N y 92°43ʼ00"0) a una altitud de 600 msnm (Figura 1). El área está limitada al Norte por los municipios de Huitiupán, Sabanilla y Tila; al Este por el municipio de Chilón; al Sur por los municipios de Pantheló, Chalchihuitán y el Bosque; al Oeste por Jitotol y Nuevo Pueblo Solistahuacán (Centro Estatal de Estudios Municipales [CEEM], 1988). El clima que presenta esta región es cálido-subhúmedo, con lluvias veraniegas en abundancia, y semicálido-húmedo con lluvias todo el año. Los meses más cálidos son abril y mayo.
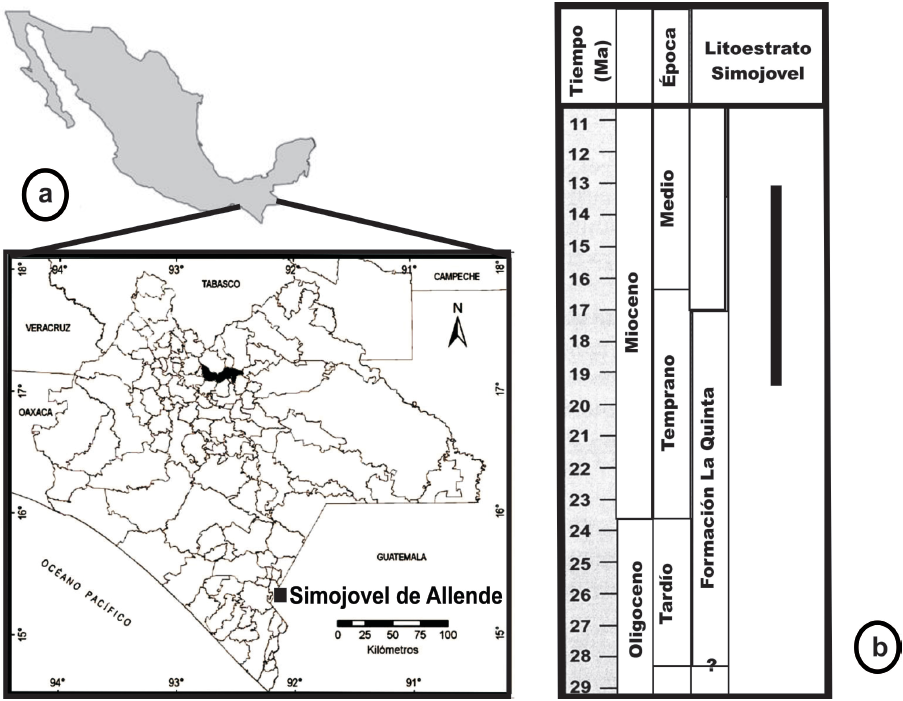
Figura 1 (a) Localización de los depósitos de ámbar de Simojovel de Allende. (b) Edad de los depósitos (Tomado y modificado de Solórzano-Kraemer, 2010).
2.2. Estratigrafía
Böse (1905) realizó una síntesis geográfica y fisiográfica de la zona, dividiendo la secuencia estratigráfica cenozoica en: división Eoceno, división Simojovel (Oligoceno), división Tenejaoa (Plioceno) y división Cuaternario. Posteriormente, Frost y Langenheim (1974) dividieron al Grupo Simojovel en Arenisca La Trinidad, Arenisca Rancho Berlín y Formación La Quinta; ésta última es donde se encuentra asociado el ámbar, aparentemente incluido en una secuencia marina del Oligoceno como lo sugieren foraminíferos planctónicos identificados, como Globigerina ciperoensis Bolli y Globerotalia kugleri Bolli reportados por los mismos autores. Sin embargo, Grimaldi (1996) mencionó que muchos depósitos de ámbar se encuentran asociados con lignitas, pizarras friables y arcillas deltaicas del Mioceno temprano. La última interpretación es consistente con estudios de nanoplanctón localizados en sedimentos de las biozonas N3 y N4, a las que se les ha asignado una edad radiométrica con rangos de 22.5 - 26.0 Ma (Berggren y Van Couvering, 1974). Sin embargo, Solórzano-Kraemer (2010) asigna el ámbar de Simojovel de Allende al Mioceno medio (15 - 20 Ma), debido al parecido que presenta con los depósitos de ámbar de la República Dominicana, incluyendo la presencia de lechos de lignita, un similar registro fósil de insectos (40 %) y su origen botánico por especies extintas de Hymenaea . Sin embargo, Vega et al. (2009) usando 87Sr/86Sr en conchas bien preservadas de Turbinella maya de Los Pocitos, una localidad muy cercana a Simojovel, establecen una edad de 23 Ma.
2.3. Especímenes
El material descrito e identificado es una flor preservada en ámbar del Mioceno de Simojovel de Allende, perteneciente a la colección del Museo de Paleontología "Eliseo Palacios Aguilera", de la Secretaría de Medio Ambiente, Vivienda e Historia Natural (SEMAVIHN), de Tuxtla Gutiérrez, Chiapas, con número de catálogo 5963-IHNFG-2919 (IHNFG-Instituto de Historia Natural-Fósiles Geográficos). La flor fósil, inicialmente fue descrita e identificada por Gómez-Bravo (2005) como un miembro de Salicaceae. Para llevar a cabo su reexaminación, se usó la clave electrónica para los géneros de Flacourtiaceae de Alford (2012), y se ingresaron los siguientes caracteres: sexo de la flor (17), perianto (18), arreglo del perianto (22), largo de los sépalos (23), posición del disco (24), forma de la antera (25), unión de los estambres (39), estaminodios (40) y arreglo de los estambres (42). Así mismo, se llevó a cabo una revisión bibliográfica basada en Sleumer (1980), Alford (2003), Robyns (1968), Wild (1960) y Yang y Zmarzty (2007), en las que se buscaron los taxones actuales con mayor similitud morfológica con la flor fósil. Una vez identificados, se buscaron ejemplares con estructuras reproductoras en los herbarios MEXU y MEX:TUX, para ser examinados y constatar su parecido.
3. Resultados
3.1. Descripción sistemática
Familia: Salicaceae Mirbel, 1815
Tribu: Samydeae (Vent.) Dumort, 1829
Género: Lunania Hooker, 1844
Lunania floresi Hernandez-Damián, Calvillo-Canadell, Cevallos-Ferriz sp. nov.
Diagnosis. Flor bisexual; inconspicua, pedicelada, 2 bractéolas; sépalos 3, parcialmente fusionados hacia la base, cóncavos; pétalos ausentes; disco hipógino, lobado; estambres 15, distribuidos en dos verticilos (el externo más largo que el interno), anteras bitecas, biloculares, basifijas, ovado-elípticas, ligeramente apendiculares; ovario súpero, pubescente y 3 estilos cortos.
Descripción. Flor bisexual de 4.0 mm x 2.0 mm, inconspicua, actinomorfa, pedicelo de 1.0 mm x 0.2 mm, punctato, subglabro, con 2 cicatrices de bractéolas en la base; sépalos 3 (Figura 2, a, b c), parcialmente fusionados hacia la base, cóncavos, ca . 1.5 mm x 0.5 mm, venación reticulada, punctato, subglabros en haz y envés; pétalos 0; disco hipógino, lobado, lóbulos cimbiformes (Figura 2, c); estambres 15, libres, distribuidos en dos verticilos, estambres del verticilo exterior de ca . 1.5 mm x 0.1 mm, estambres del verticilo interior de ca . 1.0 mm x 0.1 mm, filiformes (Figura 2, d); anteras bitecas, biloculares, ovado-elípticas, basifijas 0.5 mm x 0.4 mm, ligeramente apendiculares (Figura 2, e); ovario súpero, ovado y pubescente; 3 estilos terminales y cortos (Figura 2, f).

Figura 2 Lunania floresi sp. no v. (a) Flor bisexual, actinomorfa, pedicelada, con 3 sépalos cóncavos (escala = 1.0 mm). (b) Pedicelo. Flecha señalando 2 cicatrices de bractéolas (escala = 1.0 mm). (c) Flecha señalando disco hipógino con lóbulos (escala = 1.0 mm). (d) Flecha señalando estambres libres distribuidos en dos verticilos (escala = 1.0 mm). (e) Flecha señalando antera basifija y apendicular (escala = 0.5 mm). (f) Ovario súpero, pubescente. Flecha señalando tres estilos cortos (escala = 0.5 mm).
Epíteto específico. El nombre hace referencia a la Dra. Hilda Flores Olvera, botánica mexicana que ha tenido un enorme interés en el desarrollo de la paleobotánica en el país.
Ejemplar examinado. 5963-IHNFG-2919 (IHNFG-Instituto de Historia Natural-Fósiles Geográficos).
Repositorio. Colección del Museo de Paleontología "Eliseo Palacios Aguilera", de la Secretaria de Medio Ambiente, Vivienda e Historia Natural (SEMAVIHN) de Tuxtla Gutiérrez, Chiapas.
4. Discusión y conclusión
Un nuevo miembro de la familia Salicaceae s.l . fue identificado en el ámbar del Mioceno de Simojovel de Allende, con base en una flor bisexual, pedicelada e inconspicua, que presenta 2 cicatrices de bractéolas, 3 sépalos, pétalos ausentes, disco hipógino, lobado y ovario súpero con 3 estilos. Estos caracteres han permitido ubicarla en el género Lunania . Al comparar la morfología de la flor fósil con la de especies actuales de Lunania se reconoció un mayor parecido con L. mexicana y L. parviflora (Tabla 1), destacando el tamaño de los filamentos del fósil con el de L. mexicana (~1.5 mm), así como las anteras apiculadas presentes en L. parviflora (Vázquez-Torres et al., 2010; Robyns, 1968). A pesar del enorme parecido que presentan entre ellas existen también diferencias importantes como el número de estambres (15 vs. 10 - 12), la vestidura de la superficie del ovario pubescente en el fósil, subglabro en L. mexicana y tomentoso en L. parviflora y el tamaño de los estilos (0.3 mm vs. 1.0 mm) (Figura 3). Las observaciones y comparaciones morfológicas realizadas indican que el fósil presenta un mosaico de caracteres entre L. mexicana y L. parviflora así como caracteres únicos, lo que sugiere que se trata de una especie nueva y extinta de Lunania , constituyendo el primer registro del género y de Salicaceae para la zona.
Tabla 1 Comparación de caracteres florales entre Lumania floresi , L. mexicana y L. parviflora (Sleumer, 1980; Vázquez-Torres et al., 2010; Robyns, 1968).
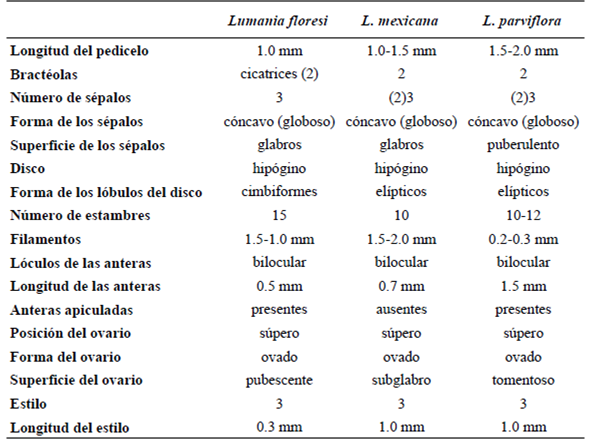
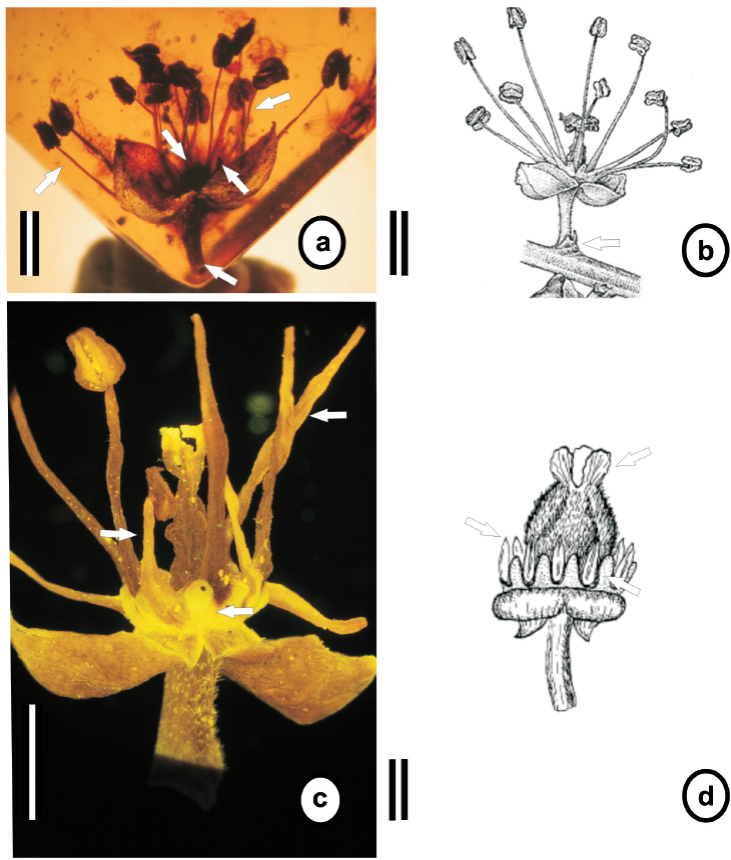
Figura 3 Comparación entre Lunania floresi , L. mexicana y L. parviflora . (a) L. floresi flor bisexual, flecha inferior derecha señalando cicatrices de bractéolas, flecha superior izquierda señalando estambres, flecha media derecha señalando lóbulos del disco, flecha media izquierda señalando ovario súpero con 3 estilos (escala = 1.0 mm). (b) L. mexicana . Flecha inferior derecha señalando bractéolas, flecha superior señalando estambres (escala = 1.0 mm) (tomado y modificado de Nee, 1999). (c) L. mexicana flor bisexual, pequeña pedicelada, flecha inferior derecha señalando lóbulos del disco, flecha superior izquierda y derecha señalando dos verticilos de estambres (escala = 0.5 mm) (Ejemplar examinado y muestreado: Lunania mexicana Brandegee VERACRUZ: Mpio. San Andrés Tuxtla, Loc. Estación Biológica Tropical Los Tuxtlas. 95°04ʼ y 95°09ʼ E y 18°34ʼ y 18°36ʼ. Lote 67 Alt. 200 msnm Selva alta perennifolia. 13 feb. 1984. G. Ibarra 1257. MEXU:TUX.) (d) L. parviflora , flecha inferior derecha señalando lóbulos del disco, flecha superior izquierda señalando estambres y flecha superior derecha señalando ovario tomentoso y 3 estigmas pequeños (escala= 0.5 mm) (tomado y modificado de Robyns, 1968).
En México el registro fósil de la familia se encuentra constituido por hojas de Populus , provenientes del Eoceno de Nuevo León (Pérez-Maussan, 2013), así como del Oligoceno de Puebla, donde además se ha reportado Salix (Ramírez y Cevallos-Ferriz, 2000). La presencia de Lunania en el Mioceno de Chiapas, amplía el registro de la familia espacial y temporalmente en México, esto en su actual circunscripción al haber desaparecido la familia Flacourtiaceae (Chase et al., 2002).
El registro fósil de Salicaceae se extiende desde el Paleoceno, con hojas de Populus del norte de los Estados Unidos de Norte América (Wing, 1981; Collinson, 1992), aunque el primer registro inequívoco de la familia está representado por frutos y hojas del Eoceno, de la Formación Green River, Utah y Colorado (Manchester et al, 1986; Boucher et al., 2003; Manchester et al ., 2006). Sin embargo, cabe señalar que el registro fósil de Salicaceae s.l . está constituido por órganos vegetativos y reproductivos de distintas edades, que incluyen por ejemplo maderas de Aphloioxylon del Paleoceno de Groenlandia (Mathiesen, 1961), Flacourtioxylon del Eoceno de Europa (Greguss,1969; Trivedi y Srivastava, 1986; Crawley, 2001), Homalioxylon del Mioceno de Europa y Asia (Prakash y Tripathi, 1972; Bande, 1974; Gottwald, 1997), Hydnocarpoxylon del Mioceno-Plioceno de Asia (Bande y Khatri, 1980; Awasthi y Srivastava, 1989); hojas de Idesia del Mioceno de California (MacGinitie, 1937), semillas y frutos de Poliothyrsis del Oligoceno-Mioceno de Alemania (Mai, 1980), así como Saxifragispermum del Eoceno de Inglaterra (Chandler, 1961) y Oregon (Manchester, 1994). Así mismo, se han reportado casos excepcionales de órganos en conexión orgánica como lo demuestra Pseudosalix handleyi del Eoceno de Estados Unidos (Colorado) (Boucher et al ., 2003).
Actualmente en México el género Lunania se encuentra representado por la especie Lunania mexicana , la cual crece en selvas altas y medianas de Veracruz, Oaxaca y Chiapas, llegando a Guatemala y Panamá (Nee, 1999; Vázquez-Torres et al., 2010). La presencia de Lunania floresi , en el Mioceno de Chiapas soporta la idea del desarrollo de una selva tropical durante el tiempo de depósito del ámbar, tal y como lo sugiere el registro fósil de plantas y animales incluidos en el ámbar de Simojovel de Allende (e.g . Miranda, 1963; Poinar y Brown, 2002; Poinar, 2002; Castañeda-Posadas y Cevallos-Ferriz, 2007; Calvillo-Canadell et al., 2010; Solórzano-Kraemer, 2010; Chambers et al., 2012), constatando a su vez la larga historia de la integración de la vegetación en México.
La distribución de fósiles de Salicaceae en el tiempo sugiere el desplazamiento del grupo de latitudes altas a latitudes bajas de América del Norte, reforzando así la propuesta de la expansión de algunos elementos de la llamada Flora Boreotropical (e.g. , Boucher, 2003; Calvillo-Canadell, 2010; Collinson, 1992; Gómez-Bravo, 2005; Pérez-Maussán, 2013) a las zonas tropicales de América, especialmente durante el Eoceno-Mioceno. En tiempos más recientes el área de distribución de algunos taxa de ésta flora se amplió a Centro y Sur América. Aunque esta interpretación de la contribución de taxa de latitudes altas de Norte América a la flora y vegetación de la zona tropical de América es atractiva, la influencia de los elementos septentrionales es indudable y el entendimiento de la forma en que plantas de los hemisferios Norte y Sur se sumaron para formar la vegetación actual de la zona Neotropical constituye un tema de álgida discusión, que sólo a través de la suma de estudios detallados se logrará integrar una propuesta más sólida sobre su origen a través del tiempo.

