










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletín de la Sociedad Geológica Mexicana
versão impressa ISSN 1405-3322
Bol. Soc. Geol. Mex vol.66 no.1 Ciudad de México Abr. 2014
Artículos
Nuevos registros del género Tempskya CORDA, 1845 (Pteridophyta) en depósitos del Hauteriviense Superior-Barremiense Inferior del norte de España
New records of the genus Tempskya CORDA, 1845 (Pteridophyta) in late Hauterivian-early Barremian deposits in northern Spain
Estefanía Puente-Arauzo1, Luis Miguel Sender1,*, Uxue Villanueva-Amadoz2, José Bienvenido-Diez3, Fidel Torcida Fernández-Baldor4
1 Área de Paleontología, Universidad de Zaragoza, C/Pedro Cerbuna, 12, 50009, Zaragoza, España. *lmsender@unizar.es
2 Estación Regional del Noroeste, Instituto de Geología, UNAM, L.D. Colosio y Madrid S/N, Campus Unison, Apartado Postal 1039, 83000, Hermosillo (Sonora), México.
3 Departamento de Xeociencias Mariñas e Ordenacion do Territorio, Universidade do Vigo, Campus Lagoas-Marcosende, 36200, Vigo (Pontevedra), España.
4 Colectivo Arqueológico-Paleontológico Salense, C.A.S, Museo de Dinosaurios de Salas de los Infantes, Pza. Jesús Aparicio, 9. 09600 Salas de los Infantes (Burgos), España.
Manuscrito recibido: Febrero 1, 2013.
Manuscrito corregido recibido: Abril 9, 2013.
Manuscrito aceptado: Mayo 30, 2013.
Resumen
En este estudio se presentan nuevos registros de helechos arborescentes fósiles del género Tempskya Corda, 1845, pertenecientes a la Formación Pinilla de los Moros, de edad Hauteriviense Superior - Barremiense Inferior, en la localidad de Salas de los Infantes (provincia de Burgos, norte de España). Estos registros poseen características que coinciden con aquellas presentes en la especie T. riojana Barale y Viera, 1989, pero también poseen otras características, como la presencia de islotes de esclerénquima con formas irregulares en la capa parenquimática de la corteza interna de los tallos, así como una capa de corteza intermedia que se introduce hacia el interior de la capa interna. Estos caracteres se identifican por primera vez en este taxón y podrían servir como criterio de determinación de esta especie frente a otras del género Tempskya Corda, 1845, y por lo tanto, constituye una nueva aportación al conocimiento de este taxón.
Palabras clave: Paleobotánica, helechos arborescentes fósiles, Tempskya, Cretácico Inferior, España.
Abstract
In this study we present new records of the tree fern fossil genus Tempskya Corda, 1845 from the Pinilla de los Moros Formation, late Hauterivian - early Barremian in age, in the surroundings of the village Salas de los Infantes (Burgos province, northern Spain). The studied samples exhibit similar anatomical features to those of the species T. riojana Barale y Viera, 1989 , even though they present some extra characters. These additional features include the presence of sclerenchyma islets with irregularly shaped parenchymal layer in the inner bark of stems and also an intermediate bark layer which invaginates into the inner layer. These characters are identified for the first time in this taxon, and they could serve as a criterion for differentiating this species from other Tempskya Corda, 1845 species, constituting a new contribution to the knowledge of this taxon.
Keywords: palaeobotany, fossil tree ferns, Tempskya, Early Cretaceous, Spain.
1. Introducción
El género Tempskya Corda, 1845 pertenece a un tipo de helecho arborescente que desarrolló una estructura de sostén, de varios metros de altura, en forma de "falso tronco", con tallos dispuestos en un sistema dicotómico que estaban sustentados por un empaquetamiento compacto de raíces adventicias (Tidwell et al., 2010), y cuyos restos se han encontrado exclusivamente hasta el momento en depósitos continentales del periodo Cretácico. Su origen es incierto, aunque existen formas descritas con características muy similares ya desde el Carbonífero (Galtier y Hueber, 2001). De igual modo, su evolución y relación con los helechos actuales aún se encuentra por determinar. Existe un gran número de especies descritas para este género, la mayoría de ellas encontradas en yacimientos en el Hemisferio Norte. Un hecho notable reside en que las diferencias en la morfología entre las diferentes especies descritas parecen ser mínimas. A pesar de ello, tanto las características externas del "falso tronco" como las estructuras internas de la anatomía de tallos y peciolos han servido como base para el establecimiento de las diferencias anatómicas entre ellas. Sin embargo, se ha constatado que los procesos tafonómicos tienen un papel muy importante en la fosilización de los restos, ya que la actuación de dichos procesos habría podido alterar las estructuras por lo que, según algunos autores, todas las especies definidas hasta el momento podrían englobarse en una sola (Taylor et al., 2009).
En su descripción de los registros del género Tempskya Corda, 1845 en el Albiense de Argentina. Tidwell y Wright (2003) realizan una amplia revisión del género y de sus diferentes especies en depósitos del Cretácico a nivel mundial, aunque esta revisión no es del todo completa, ya que no mencionan los ejemplares españoles. De entre estas especies, destacan por su singularidad los registros del Albiense del centro-oeste de los Estados Unidos (ver en tabla 1), así como las especies Tempskya uemurae Nishida, 2001 del Cretácico Superior (Santoniense) de Japón (Nishida, 1986; 2001), que es la especie más moderna del género encontrada hasta el momento, T. varians Mikuláš y Dvořák, 2003 del Cenomaniense Superior de la República Checa (Velenovsky, 1888; Mikuláš y Dvořák, 2003), T. dernbachii Tidwell y Wright, 2003 del Albiense de la Patagonia en Argentina (Tidwell y Wright, 2003) y T. judithae Clifford y Dettmann, 2005 del Albiense Superior de Australia (Clifford y Dettmann, 2005).
Un dato muy significativo respecto al conocimiento de este taxón a nivel mundial, reside en que los registros más abundantes y con mayor número de especies de este género corresponden a los hallados en los depósitos del centro y oeste de los Estados Unidos. Este hecho puede ser debido, probablemente, a las excepcionales condiciones de afloramiento y a los numerosos investigadores que han estudiado los registros paleobotánicos de esta zona a lo largo de la historia (ver tabla 1). Dichos yacimientos han sido considerados históricamente como de edad Albiense, pero estudios radiométricos recientes parecen indicar que las edades de algunos de estos registros serían más modernas, llegando incluso al Cenomaniense Inferior (Tidwell et al., 2010).
En España, los únicos registros del género Tempskya Corda, 1845 conocidos hasta el momento pertenecen a la especie Tempskya riojana Barale y Viera, 1989 procedente de los depósitos del Valanginiense-Hauteriviense de la región de La Rioja (Barale y Viera, 1989; 1991) y del Cretácico Inferior indeterminado en la región de Soria (Crisafulli et al., 2008). Sin embargo, la edad de este último registro presenta ciertas reservas ya que se encontró rodado como un clasto en depósitos fluviales paleógenos, argumentando los autores que la supuesta área fuente de los restos vegetales se ubicaría en afloramientos de rocas del Cretácico Inferior que habrían estado situados aguas arriba de la terraza fluvial en donde fueron encontrados.
En el presente trabajo, se estudian nuevos hallazgos del género Tempskya Corda, 1845 en depósitos continentales del Cretácico Inferior de España, datados con una edad de Hauteriviense Superior - Barremiense Inferior, realizando un estudio sistemático completo de los mismos y comparando sus características con las de otras especies definidas previamente.
2. Localización del área de trabajo y estratigrafía
Los restos estudiados se localizaron en el yacimiento de Horcajuelos, que se encuentra en las inmediaciones de la localidad Salas de los Infantes, situada al sureste de la provincia de Burgos (norte de España). Geográficamente, la zona de estudio se dispone entre la Cuenca del Duero al sur y la Sierra de la Demanda al norte, y geológicamente se ubica dentro del sector noroccidental de la Cuenca de Cameros (Figura 1), que es una de las cuencas formada por depósitos mesozoicos que se desarrolló a principios del Cretácico Inferior en el este de Iberia. Esta cuenca se sitúa en la parte Noroccidental de la Cordillera Ibérica y limita al norte con los materiales cenozoicos de la depresión del Ebro, al sur con la cuenca terciaria de Almazán, al sureste con el macizo paleozoico del Moncayo y al noroeste con el macizo de materiales paleozoicos de la Demanda.
El yacimiento estudiado se encuentra en materiales pertenecientes a la Formación Pinilla de los Moros, dentro del Grupo Urbión, con una edad Hauteriviense Superior - Barremiense Inferior a partir de su datación por carofitas (Martín-Closas y Alonso-Millán, 1998) (Figura 1). Esta formación, presenta en este sector occidental de la cuenca de Cameros una potencia de hasta 450 m y está formada por areniscas en cuerpos canalizados alternando con lutitas rojas. Lateralmente, la litología de los materiales cambia a calizas oncolíticas, margas, lutitas y areniscas en cuerpos lenticulares de pequeño desarrollo (Martín Closas y Alonso-Millán, op. cit.).
La serie estratigráfica de la Formación Pinilla de los Moros en el yacimiento de Horcajuelos (Figura 2), consiste en amplios y potentes niveles, de espesor métrico, de lutitas de color rojo muy oscuro, con intercalaciones de paquetes de potencia decimétrica a métrica de areniscas blancas, grises y ocres de grano fino a medio, a veces en cuerpos con geometría canaliforme de hasta 2 m de espesor y con lags basales conglomeráticos. Algunos de estos cuerpos presentan superficies de acreción lateral, aunque generalmente el relleno está formado por sets de estratificación cruzada en surco. A techo de las areniscas, se encuentran esporádicas intercalaciones de niveles de lutitas arenosas que presentan bioturbación de hábito vertical y esporádicos niveles de lutitas carbonatadas laminadas. El medio de depósito de estos materiales correspondería a un medio fluvial de cauces anastomosados, separados por amplias llanuras aluviales, con desarrollo de llanuras de inundación entre los canales, con depósitos proximales de levee y crevasse-splay y con desarrollo de paleosuelos en algunos puntos (Clemente et al., 1991, Clemente y Pérez-Arlucea, 1993; Martín-Closas y Alonso-Millán, 1998).
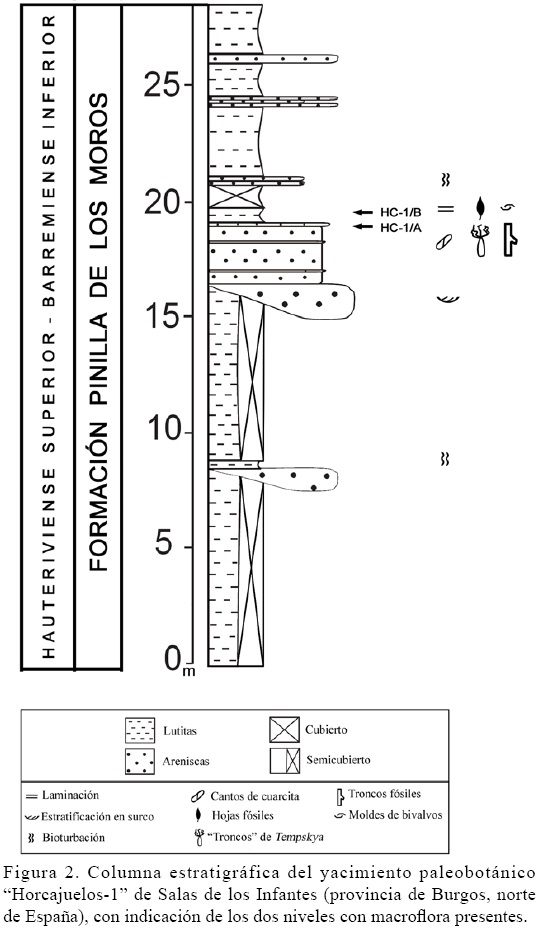
El yacimiento está constituido por dos niveles (HC-1/A y HC-1/B), diferenciados por sus tipos de facies. El nivel inferior (HC-1/A), está formado por areniscas anaranjadas de grano fino, con intercalaciones de niveles de arenas blancas compactadas de grano fino de potencia decimétrica con abundantes costras ferruginosas. A techo de la última capa de areniscas compactadas es en donde aparecen los restos de "falsos troncos" del género Tempskya Corda, 1845 estudiados en este trabajo (Figura 3A), así como fragmentos de troncos de coníferas de longitud decimétrica. Estos niveles, se corresponderían con depósitos de relleno arenoso de canales fluviales anastomosados.
El nivel superior (HC-1/B), está formado por un conjunto de paquetes de lutitas laminadas más arenosas a la base y que se van haciendo más carbonatadas hacia techo. Estas capas son ricas en moldes de bivalvos y conchas de gasterópodos turbinados ambos de pequeño tamaño, y en ellas se ha encontrado una asociación vegetal conservada compuesta por pteridofitas, con pinnas de helechos correspondientes a los taxones Onychiopsis cf. psilotoides (Stokes y Web) Ward, 1905, Ruffordia sp. (Dunker) Seward, 1894 y Cladophlebis sp. Brongniart, 1849 y pinnulas pertenecientes al taxón Regnellites sp. Yamada y Kato, 2002 así como fragmentos de hojas de gimnospermas indeterminadas pertenecientes al morfogénero Desmiophyllum Lesquereux, 1878 (Sender et al., 2011). Estos depósitos, con abundantes restos de flora y conchas de moluscos, fueron acumulados en un pequeño cuerpo de agua situado en las cercanías de un canal, relacionado con un episodio de inundación.
3. Métodos y materiales estudiados
Se han realizado varias secciones pulidas así como preparaciones en lámina delgada de los restos de "falsos troncos" (Figura 3D, E), para lo cual se han seleccionado los cuatro mejores especímenes, correspondientes a fragmentos del eje de crecimiento, de entre los 79 restos recogidos en el yacimiento. Para el estudio de las láminas delgadas, se ha utilizado un microscopio en luz transmitida del modelo Olympus B×41 con una cámara acoplada de modelo Olympus C5060-ADU trabajando a una resolución de 1600×1200H.
Los restos estudiados se recogieron en varias campañas de prospecciones entre los años 1999 y 2009. La variedad morfológica de los restos encontrados, así como el hallazgo de varios fragmentos apicales, indicarían la presencia de varios ejemplares individualizados de "falsos troncos" en el yacimiento. Todos los restos se presentan conservados como permineralizaciones en sílice, en algunos casos con un elevado grado de recristalización (Figura 3B). Entre ellos, destaca un ejemplar de un fragmento de "falso tronco" de 89 cm de longitud y 17,5 cm de anchura (Figura 3C), referenciado por Huerta-Hurtado y Torcida Fernández-Baldor (1999).
Todos los restos estudiados se encuentran depositados en el Museo de Dinosaurios de Salas de los Infantes, provincia de Burgos (España).
4. Paleontología sistemática
División Pteridophyta Schimper, 1879
Clase Polypodiopsida Cronquist, Takht. y Zimmerm., 1966
Familia Tempskyaceae Read y Brown, 1937.
Género Tempskya Corda, 1845.
Especie tipo: Tempskya pulchra Corda, 1845.
Tempskya riojana Barale y Viera, 1989, p. 110, Text-Figs.
1–6; Láms. 1–5.
Tempskya aff. riojana
Figura 3 (b-e), Figura 4, Figura 5
Los ejemplares estudiados, se componen de fragmentos de "falsos troncos" formados por tallos solenostélicos individuales, inmersos en una densa matriz de raíces.
Los tallos solenostélicos o estípites presentan en su mayoría una forma ovalada (posiblemente debido a procesos de deformación). El diámetro de los estípites se encuentra entre 1.8-3.5 mm. En los ejemplares estudiados, se ha observado que la corteza mantiene una división en tres partes: corteza externa, corteza intermedia y corteza interna (Figura 4A, B).
La corteza externa posee un grosor de entre 200-250 μm, aunque en la mayoría de los casos presenta un considerable grado de recristalización, en algunos ejemplares todavía es posible apreciar la zona más interna de esta capa externa, en parte debido a su mejor preservación. Justo en esta zona limítrofe con la capa intermedia, se presentan de dos a tres capas de células alargadas de tipo parenquimatoso (Figura 4C).
La corteza intermedia presenta un mayor desarrollo (250 a 500 μm de espesor), y se encuentra formada por células esclerenquimatosas de pequeño tamaño y de formas variables, desde poligonales a ovaladas (con una longitud de entre 25 a 35 μm), separadas por espacios intercelulares que tienden a ser equidimensionales. Estas células se encuentran organizadas formando capas concéntricas, presentándose de 10 a 15 capas de células (Figura 4C). Su límite externo se encuentra bien delimitado pero su zona más interna, limítrofe con la corteza interna, se encuentra más precariamente definida, ya que parte del esclerénquima de la capa intermedia se introduce a través de la corteza interna por medio de entrantes que penetran en esta última (Figura 4C).
La corteza interna tiene un espesor de entre 200 a 300 μm, y está formada por células parenquimatosas de tamaño variable (de 40 a 60 μm de longitud). Estas células poseen una forma morfología poligonal y se disponen formando entre 6 a 8 capas, dispuestas concéntricamente, y con espacios intercelulares de dimensiones variables. Esta capa presenta islotes de células de esclerénquima de pequeño tamaño (25-35 μm), inmersos entre las células parenquimatosas predominantes y que aparecen distribuidas por toda la corteza interna (Figura 4C).
El endodermo, el periciclo y el floema que se dispondrían entre la corteza interna y el xilema, no aparecen diferenciados en los ejemplares estudiados, debido a las condiciones de preservación de los restos, observándose una zona hueca o masivamente recristalizada rodeando externamente al xilema (Figura 4D).
El xilema está formado por células de gran tamaño (de entre 50 a 70 μm de longitud) que presentan una morfología ovalada a poligonal. Estas células se organizan en grupos de capas concéntricas, de entre 5 a 8 capas, que presentan espacios intercelulares de gran tamaño, en proporción, y con morfología irregulares (Figura 4B, C). La médula, se encuentra homogéneamente compuesta por células de esclerénquima de pequeño tamaño (de 25 a 35 μm de longitud), sin presentar ninguna zonación interna basada en tamaño, o forma de las células que lo componen (Figura 4D).
Las inserciones de los peciolos en los tallos se encuentran organizadas en dos filas, alineadas y situadas en el mismo lado del tallo Los internodos son largos, distinguiéndose, en sección, entre una, y dos emisiones peciolares. Los peciolos presentan un diámetro que oscila entre 1.3 y 1.6 mm y se encuentran rodeados por completo por una fila de raíces. La epidermis del peciolo se encuentra pobremente preservada. La corteza aparece bien diferenciada, formada por una única capa con una anchura de entre 250 y 300 μm y con una distribución muy ordenada de pequeñas células esclerenquimáticas de 25 μm de diámetro, cuyos espacios intercelulares se encuentran muy reducidos. La parte más interna del peciolo alberga una porción de xilema (Figura 4E) con una característica morfología en forma de "media luna", debido al estrechamiento y posterior estrangulamiento que se produce en el xilema en las últimas fases del proceso de emisión peciolar (Barale y Viera, 1989).
Las raíces muestran, en sección transversal, una orientación vertical predominante que en algunos casos pasa a ser aleatoria e incluso totalmente transversal a la dirección de crecimiento (Figura 5C). Las raíces son muy numerosas y forman una densa matriz que sostiene la estructura de tallos y peciolos. La mayoría de las raíces aparecen orientadas de forma paralela al eje de crecimiento, (Figura 5C) aunque en ocasiones también se pueden encontrar orientadas en dirección oblicua o incluso perpendicular con respecto al conjunto (Figura 5E).
En la corteza que cubre las raíces se diferencian dos capas: corteza externa y corteza interna. La corteza externa de las raíces presenta entre 3 a 4 filas de grandes células parenquimatosas de hasta 120 μm de espesor, y la corteza interna está formada también por 3 a 4 filas de pequeñas células de esclerénquima, pero de menor tamaño que las de la corteza externa, con un espesor que no supera los 100 μm (Figura 5C).
El xilema interno, se encuentra típicamente dividido en protoxilema y metaxilema, en donde ambos se combinan para dar una estructura en forma de cruz (Figura 4D).
5. Discusión y conclusiones
Tras la definición del género por Corda (1845) se han sucedido diversos estudios que han intentado relacionar al género Tempskya Corda, 1845 con alguna de las familias de helechos conocidas, así como otros que han aportado datos sobre el desarrollo y el crecimiento de sus diferentes partes dentro de la estructura de "falso tronco".
Seward (1924) señaló la presencia de células engrosadas, organizadas en corona y comparables al mecanismo de anillo de un esporangio, en las raíces de Tempskya knowltoni Seward, 1924 del Cretácico Inferior de los Estados Unidos, lo que le sirvió para relacionar el género con otros con similares estructuras fértiles dentro la Familia Schizaeaceae.
Más tarde, Andrews y Kern (1947) descartaron la pertenencia a la Familia Schizaeaceae de los esporangios presentes en los ejemplares estudiados por Seward (1924), y propusieron una nueva interpretación del crecimiento de las hojas del género Tempskya Corda, 1845. En este estudio, sobre ejemplares del Albiense de Idaho (Estados Unidos), estos autores pudieron constatar el crecimiento de las hojas de este taxón a lo largo de todo el eje principal de la planta, y no solo en la parte media-superior del tallo, como se creía hasta entonces.
Ash y Read (1976) propusieron la diferenciación de dos grandes grupos de especies dentro del género Tempskya Corda, 1845, descritos a continuación, en función de la organización interna de la corteza de los tallos:
Especies del Grupo I o especies con corteza simple: corteza interna y externa constituida por células parenquimatosas, y corteza intermedia formada por células esclerenquimatosas. A este grupo corresponderían las especies Tempskya minor Read y Brown, 1937, T. knowltonii Seward, 1924, T. rossica Kidston y Gwynne-Vaughan, 1911, T. zellerii Ash y Read, 1976.
Especies del Grupo II o especies con corteza mixta: corteza interna formada por células parenquimatosas con formas continuas o discontinuas de células esclerenquimatosas. A este grupo corresponderían las especies Tempskya grandis Read y Brown, 1937, T. reesidei Ash y Read, 1976, T.superba Arnold, 1958, T. wesselii Arnold, 1945, T. wyomingensis Arnold, 1945.
Tidwell y Hebbert (1992) publicaron una nueva y más completa clave de clasificación de los caracteres, que les permitió agrupar las principales especies del género Tempskya Corda, 1845 en dos tipos, según sus características:
Especies tipo 1. Presentan un falso tronco con simetría radial, tallos individuales de 5 a 15 mm de diámetro y corteza interna parenquimatosa con bandas continuas y/o irregulares de esclerénquima. Especies: T. grandis Read y Brown, 1937, T. superba Arnold, 1958 T. reesidei Ash y Read, 1976, T. wesselii Arnold, 1945, T. wyomingensis Arnold, 1945, T. stichkae Tidwell y Hebbert, 1992, T. readii Tidwell y Hebbert, 1992.
Especies tipo 2. Presentan un falso tronco con simetría radial o bien dorsiventral, con tallos individuales de diámetro entre 2.5 y 8.0 mm y corteza interna parenquimatosa con ausencia de bandas continuas y/o irregulares de esclerénquima. Especies: T. zellerii Ash y Read, 1976, T. minor Read y Brown, 1937, T. jonesii Tidwell y Hebbert, 1992, T. knowltonii Seward, 1924, T. dernbachii Tidwell y Wright, 2003, T. iwatensis Nishida, 1986, T. rossica, Kidston y Gwynne Vaughan, 1911.
Las características de los ejemplares estudiados en este trabajo se ajustan a las presentes en las especies tipo 1 de Tidwell y Hebbert (1992), ya que presentan largos internodos (sólo con 2 o 3 peciolos por sección) y poseen islotes de esclerénquima dentro de la corteza interna predominantemente parenquimatosa. Estas características, presentes también en la especie Tempskya riojana Barale y Viera, 1989 (Tabla 2) procedente de los depósitos del Cretácico Inferior continental de Igea y Cornago en La Rioja, España (Barale y Viera, 1989; 1991), relacionan los ejemplares estudiados de la provincia de Burgos con dicha especie. Sin embargo, la presencia de islotes de esclerénquima aislados dentro de la corteza parenquimatosa interna, es una característica adicional que no se había observado hasta el momento en esta especie, con lo que se ha optado por asignar los ejemplares estudiados a Tempskya aff. riojana.
Las características anatómicas de los ejemplares de Tempskya aff. riojana del yacimiento de Horcajuelos, también son muy similares a las encontradas en las especies Tempskya superba Arnold, 1958 y T. grandis Read y Brown, 1937, tal como ya expusieron Barale y Viera (1989) en el estudio de sus ejemplares de La Rioja. Sin embargo, Tempskya riojana Barale y Viera, 1989 difiere de estas dos especies ya que T. superba Arnold, 1958 presenta una banda de parénquima externa (Read y Brown, 1937) y T. grandis Read y Brown, 1937 muestra una banda interna de esclerénquima en la estructura de la médula (Arnold, 1958).
Andrews y Kern (1947) observaron como el proceso de dicotomización de los tallos permitía una diversificación de los mismos dentro del "falso tronco". Según su modelo, el proceso comenzaría desde un tallo inicial situado en la zona más basal que, por medio de sucesivas divisiones, desarrollaría un número elevado de tallos en la zona más apical. Estos autores describieron el proceso de dicotomización desde las fases más juveniles hasta las más adultas, como un desarrollo de numerosas dicotomizaciones sucesivas en dirección adapical, a la vez que la anchura del "falso tronco" también disminuiría adapicalmente. De este modo, al llevarse a cabo una disminución progresiva del espacio disponible para el crecimiento de los tallos hacia el ápice del "falso tronco", se produciría la saturación de los mismos y la imposibilidad de que pudieran dividirse nuevamente debido a la falta de espacio, por lo que únicamente se producirían un número reducido de tallos. Proporcionalmente, y al mismo tiempo que el helecho fuera creciendo en altura y disminuyendo su anchura hacia el ápice, la parte basal del "falso tronco" crecería en anchura, provocando el colapso de los tallos en esta zona basal debido al masivo crecimiento de raíces necesarias para permitir la sustentación vertical de la planta.
En los ejemplares estudiados, se aprecian algunas evidencias de las etapas de dicho proceso de dicotomización de los tallos. Así, en la Figura 5A se muestra una sección de tallo con la corteza formando todavía una estructura común, presentando una invaginación en la zona de división a partir de la cual se desarrollará el proceso de dicotomización. En esta sección se aprecia también cómo el conjunto de xilema y corteza interna ya se encuentran divididos en dos partes, en un estadio de desarrollo del proceso de división previo a la etapa que produciría, posteriormente, el estrangulamiento de la médula interna. La Figura 5B muestra otra sección de un tallo que presenta el siguiente paso correspondiente con el proceso de división, en donde se aprecia cómo el xilema y la corteza interna ya aparecen individualizados albergando la médula interna, aunque la corteza intermedia-externa todavía es común a las dos partes a dividir.
En la Figura 5B se aprecia la disposición ya individualizada de dos tallos con respecto a una zona común de corteza, presente en la parte más central del conjunto, la cual se encuentra atravesada por tres raíces (Figura 5B, flecha). Este hecho representa una etapa del proceso inmediatamente anterior a la completa división de la corteza, la cual tendría lugar a partir de un movimiento centrípeto del conjunto.
En la actualidad, no existe ningún criterio que relacione directamente el número de tallos con el estadio de desarrollo de las diferentes especies del género Tempskya Corda, 1845. Sin embargo, atendiendo al modelo de crecimiento descrito por Andrews y Kern (1947), a medida que el "falso tronco" crece en altura, su anchura disminuye, produciéndose a la vez un aumento en el número de tallos hacia el ápice. En este modelo, la proporción de tallos respecto al de raíces aumenta hasta un punto en el que la dicotomización de los estípites se satura adapicalmente, debido a la disminución del espacio disponible. Este hecho implicaría que la proporción de tallos respecto a la de raíces sería mínima en las partes más basales y máxima en las zonas apicales.
De este modo, aplicando el modelo de Andrews y Kern (1947) a los restos estudiados en este trabajo, se deduce que estos pertenecerían a zonas relativamente cercanas a la base de la planta, ya que contienen entre 5 a 25 tallos por sección transversal, a diferencia de los ejemplares de Tempskya riojana Barale y Viera,1989, que contenían una media 200 tallos en sección por lo que, según dicho modelo, se corresponderían con zonas intermedias y apicales del "falso tronco".
Otra diferencia con los registros anteriormente mencionados, reside en que, además de la presencia de islotes de esclerénquima aislados dentro de la corteza parenquimatosa interna, también aparecen entrantes de esclerénquima que se adentran desde la corteza intermedia hacia la capa interna. Este tipo de estructura no ha sido descrito, hasta el momento, en ninguna de las especies identificadas del género Tempskya Corda, 1845, y podría constituir una nueva característica diferenciadora presente en la estructura interna de la especie T. riojana Barale y Viera, 1989.
Los hallazgos de "falsos troncos" de helechos, pertenecientes al género Tempskya Corda, 1845 en Salas de los Infantes (provincia de Burgos, España) de edad Hauteriviense Superior-Barremiense Inferior, aportan una nueva cita de este género en el sur de Europa occidental en una zona en donde sus registros son muy escasos, además de constituirse en unos de los registros más modernos de este género, hasta el momento, tanto en España como en el conjunto del sur europeo. El estudio en lámina delgada de las características anatómicas de los ejemplares, indica que éstas coinciden con las definidas para la especie Tempskya riojana Barale y Viera, 1989. Las secciones de los ejemplares estudiados de esta especie presentan, además de las características propias de la especie, islotes de esclerénquima y entrantes de esclerénquima desde la corteza intermedia hacia la capa interna de la estructura, los cuales constituyen dos nuevos caracteres no observados con anterioridad en estudios precedentes, que permitirían identificar esta especie frente a otras especies de este género.
Agradecimientos
Los autores quieren expresar su agradecimiento a los miembros del Colectivo Arqueológico – Paleontológico Salense (CAS) y demás colaboradores de la localidad de Salas de los Infantes por su inestimable ayuda e interés en los trabajos paleobotánicos realizados, así como a la Fundación para el estudio de los Dinosaurios en Castilla y León que financió una campaña de excavación realizada en el yacimiento Horcajuelos. Nuestro agradecimiento también a la Dra. Marta Pérez Arlucea por su ayuda en las labores de campo y sus aportaciones a la estratigrafía y sedimentología del yacimiento. Agradecemos también a la Comandancia militar de Burgos, Soria y Cantabria del Ejército de Tierra español, el habernos proporcionado los recursos materiales necesarios para el desarrollo de las tareas de campo durante la campaña de excavaciones del año 2010. Por último, los autores quieren agradecer la colaboración de todos los integrantes del equipo de excavación (Raquel Sánchez Pellicer, Borja Cascales Miñana, Paula Baltasar, Manuel García, Manuel Juncal, Fernando Mesías, Gonzalo Rial, Asier Pascual y Antoine Bercovici) sin cuya ayuda y dedicación no podría haberse llevado a cabo este trabajo. Este estudio es una contribución al Proyecto de Investigación y Difusión: "Paleoflora de los ecosistemas cretácicos en la provincia de Burgos" (Expte. 171/09-BU), financiado por la Junta de Castilla y León.
Referencias
Andrews, H.N., Kern, E. M., 1947, The Idaho tempskyas and associated fossil plants: Annals of the Missouri Botanical Garden, 34(2), 119-183. [ Links ]
Arnold, C.A., 1945, Silicified plant remains from the Mesozoic and Tertiary of western North America: Michigan Academy of Science, Arts and Letters, 30, 3-34. [ Links ]
Arnold, C.A., 1958, A new Tempskya: Contributions from the Museum of Paleontology. University of Michigan, 14(8), 133-142. [ Links ]
Ash, S.R., Read, C.B., 1976, North American species of Tempskya and their stratigraphic significance: United States Geological Survey professional paper, 847, 1-42. [ Links ]
Barale, G., Viera, L.I., 1989, Tempskya riojana nov. sp., fougère arborescente du Crétacé inférieur du Nord-Ouest de L'Espagne: Palaeontographica Abteilung B, 212, 103-122. [ Links ]
Barale, G., Viera, L.I., 1991, Description d'une nouvelle paléoflore dans le Crétacé inférieur du Nord de l'Espagne: Munibe, 43, 21-35. [ Links ]
Clemente, P., Pérez-Arlucea, M., 1993, Depositional architecture of Cuerda del Pozo Formation, Lower Cretaceous of extensional Cameros Basin, North-Central Spain: Journal of Sedimentary Petrology, 63(3), 437-452. [ Links ]
Clemente, P., Alonso, A., Pérez-Arlucea, M., 1991, Secuencias de depósito en la parte occidental de la Cuenca de Los Cameros. Jurásico terminal-Cretácico inferior (resumen), en Tercer Coloquio del Cretácico de España, Morella, 1, 21. [ Links ]
Clifford, H.T., Dettmann, M.E., 2005, First record from Australia of Cretaceous fern genus Tempskya and the description of a new species, T. judithae: Review of Palaeobotany and Palynology, 134, 71-84. [ Links ]
Corda, A.J., 1845, Flora Protogaea, en Calvary, S., Co. (ed.), Beiträge zur Flora der Vorwelt, 60, Tafeln, Berlin, 1-128. [ Links ]
Crisafulli, A., Sanchez-Hernandez, B., Herbst, R., 2008, Filices y Bennettitales del Cretácico Inferior de Carazuelo, provincia de Soria, España: Geobios, 41, 589-604. [ Links ]
Cronquist, A., Takhtajan, A., Zimmermann, W., 1966. On the higher taxa of Embryophyta, Taxon, 14(4), 129-134. [ Links ]
Galtier, J., Hueber, F.M., 2001, How early ferns became trees: Proceedings of the Royal Society of London, B, 268, 1955-1957. [ Links ]
Huerta-Hurtado, P., Torcida Fernández-Baldor, F. (1999), Hallazgos de restos vegetales fósiles en Salas de los Infantes (Burgos, España), en Colectivo Arqueológico- Paleontológico de Salas (ed.), Actas de las I Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno: Salas de los Infantes, Burgos, España, 21-25 de septiembre de 1999, 371-380. [ Links ]
Kidston, R., Gwynne-Vaughan, D.T., 1911, On a new species of Tempskya from Russia: Verhandlungen der Russisch- Kaiserlichen Mineralogischen Gesel, 48, 1-20. [ Links ]
Lesquereux, L., 1878, On the Cordaites and their related generic divisions in the Carboniferous formation of the United States: Proceedings of the American Philisophical Society, 17, 315-355. [ Links ]
Martín-Closas, C., Alonso Millán, A., 1998, Estratigrafía y bioestratigrafía (Charophyta) del Cretácico Inferior en el sector occidental de la Cuenca de Cameros (Cordillera Ibérica): Revista de la Sociedad Geológica de España, 11(3-4), 253-270. [ Links ]
Mikuláš, R., Dvořák Z., 2003, Vrtby v dřevitých tkáních stromových kapradin rodu Tempskya (česká křídová pánev) / Borings in xylic tissues of the tree fern Tempskya (Bohemian Cretaceous Basin, Czech Republic): Zprávy o geologických výzkumech, 2002, 129-131. [ Links ]
Nishida, H., 1986, A new Tempskya stem from Japan: Transactions and proceedings of the Palaeontological Society of Japan, New series, 143, 435-446. [ Links ]
Nishida, H., 2001, A Leptosporangiate Fern Tempskya uemurae, sp. nov. (Tempskyaceae) from the Upper Cretaceous (Santonian) of Iwate Pref., Japan: Acta Phytotaxonomica et Geobotanica, 52, 41-48. [ Links ]
Pereda-Suberbiola, X., Ruiz-Omeñaca, J.I., Torcida Fernández-Baldor, F., Maisch, M.W., Huerta, P., Contreras, R., Izquiedo, L.A., Montero-Huerta, D., Urién-Montero, V., Welle, J., 2011, A tall-spined ornithopod dinosaur from the Early Cretaceous of Salas de los Infantes (Burgos, Spain): Comptes Rendus Palevol, 10, 551-558. [ Links ]
Read, C.B., Brown, R.W., 1937, The American Cretaceous ferns of the genus Tempskya: United States Geological Survey professional paper, 186-F, 105-131. [ Links ]
Schimper, W.P., 1879, Palaeophytologie, en Zittel, K.A. (ed.), Handbuch der Palaeontologie, 2 (1) 1-152. 117 figs. [ Links ]
Sender, L.M., Villanueva-Amadoz, U., Diez, J.B., Torcida Fernández-Baldor, F Sánchez-Pellicer, R., Cascales-Miñana, B., Bercovici, A., Huerta, P., Ferrer, J., 2011. Nuevos registros paleobotánicos del Cretácico Inferior de la cuenca de Cameros occidental (provincia de Burgos): Geogaceta, 50(2), 181-184. [ Links ]
Seward, A.C., 1894, Catalogue of the Mesozoic plants in the Department of Geology, British Museum Natural History: The Wealden Flora, I, Thallophyta-Pteridophyta, London, 179 pp. [ Links ]
Seward, A.C., 1924. On a new species of Tempskya from Montana Tempskya Knowltoni, sp. nov.: Annals of Botany, 38(151), 485-507. [ Links ]
Stopes, M.C., 1915, Lower Greensand (Aptian) plants of Britain, in Catalogue of the Mesozoic plants in the British Museum (Natural History). The Cretaceous Flora. Part II, 9-21. [ Links ]
Taylor, T.N., Taylor, E.L., Krings, M., 2009, Paleobotany. The biology and evolution of fossil plants, 2° Edition: Elsevier, 1252 p. [ Links ]
Tidwell, W.D., Hebbert, N., 1992, Species of the Cretaceous tree fern Tempskya from Utah: International Journal of Plant Sciences, 153, 513-528. [ Links ]
Tidwell, W.D., Wright, W.W., 2003, Tempskya dernbachii sp. nov. from Neuquén Province, Argentina, the first Tempskya species reported from the Southern Hemisphere: Review of Palaeobotany and Palynology, 127, 133-145. [ Links ]
Tidwell, W.D., Ash, S.R., Britt, B.B., 2010, Oldest Known Dicotyledonous Lianas from the Early Late Cretaceous of Utah and New Mexico, U.S.A., en Gee, C.T. (ed.), Plants in Mesozoic Time. Morphological Innovations, Phylogeny, Ecosystems: Bloomington & Indianapolis, Indiana University Press, 271-291. [ Links ]
Velenovsky, J., 1888, Die Farne der Böhemischen Kreideformation: Abhandlungen der Königlichen Böhmischen Gesellschaft der Wissenschaften, 7, 2(8), 1-32. [ Links ]
Ward, L.F., 1905, Status of the Mesozoic Floras of the United States: United States Geological Survevey Monographs, 48, 1-616. [ Links ]
Yamada, T., Kato, M., 2002, Regnellites nagashimae gen. et sp. nov., the oldest macrofossil of Marsileaceae, from the Upper Jurassic to Lower Cretaceous of Western Japan: International Journal of Plant Sciences, 163, 715-722. [ Links ]

