











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Los frutales caducifolios se caracterizan por un periodo de baja actividad hormonal y metabólica en los meristemos, el cual es denominado “reposo”. Este periodo es una adaptación a los inviernos fríos (Horvath, 2009) que permite desarrollar mecanismos de resistencia a los daños por frío (Elle y Sauter, 2000; Campoy et al., 2011). La aclimatación al frío de las yemas florales en varias especies de Prunus L. está determinada por factores genéticos (Arora y Wisniewski, 1994; Yamane et al., 2011), fisiológicos, bioquímicos (González-Rossia et al., 2008; Die y Rowland, 2014), morfológicos y anatómicos (Ashworth, 1982; 1984). Las temperaturas bajas se requieren para satisfacer las necesidades fisiológicas de las yemas florales (Andreini et al., 2014). Una deficiencia en horas frío provoca floración deficiente, malformaciones en los verticilos vegetativos y reproductivos (Nava et al., 2009; Reig et al., 2013). Al final del “reposo”, la yema floral del duraznero tiene cambios bioquímicos, fisiológicos y anatómicos relacionados con el establecimiento de la conexión vascular, lo cual aumenta el riesgo de congelación (Ashworth, 1982; 1984; Wisniewski y Davis, 1989), pero no se conoce con precisión cuando y en que tejidos de la yema sucede la diferenciación del xilema, tejido responsable de la acumulación de agua en los tejidos. En el “reposo” la actividad enzimática aumenta y la concentración de azúcares disminuye, lo cual aumenta la susceptibilidad al frío de las yemas florales de duraznero (Palonen, 1999) y la concentración baja del almidón en el duraznero y peral (Pyrus communis L.) (Ito et al., 2002 y González-Rossia et al., 2008), pero no se conoce la secuencia de estos eventos en relación con el estado fenológico y fecha de floración. Respecto al desarrollo de la antera, en chabacano (Prunus armeniaca L.) el cambio de color de las anteras se asocia a la meiosis y al fin del “reposo” de las yemas (Julian et al., 2014). Reinoso et al. (2002) describieron el desarrollo anatómico de las yemas florales de duraznero desde un punto de vista botánico, pero no revisaron la diferenciación del tejido vascular ni la presencia de granos de almidón. El xilema es el responsable de conducir el agua hacia las yemas y el almidón es una importante fuente de energía para la morfogénesis; la descripción minuciosa de estos eventos en duraznero es una herramienta eficaz en el manejo agronómico y reducción de daño por heladas. El objetivo del presente estudio fue caracterizar el desarrollo del xilema y la distribución de granos de almidón, en cinco estados fenológicos (EF), desde el EF “reposo” hasta el EF “estambres visibles” en tres cultivares de P. persica con diferentes fechas de floración. En el supuesto de que la diferenciación del xilema esté relacionada con un estado fenológico particular de la yema floral, esto permitirá predecir el grado de vascularización y la susceptibilidad al frío en cultivares con diferente fecha de floración.
Materiales y métodos
Material vegetal y sitio experimental
Los cultivares de Prunus persica usados y fechas de antesis fueron: Puebla (12 enero, precoz), CP Precoz (20 febrero, intermedio) y Zacatecas (17 marzo, tardío) durante el invierno 2011-2012. Los árboles de cinco años de edad estaban injertados en duraznero criollo. La huerta está en el Campo Experimental Valle de México (CEVAMEX), el Horno, Chapingo, Texcoco, Estado de México, (19° 29’ 17” N y 98° 53’ 41” O, altitud de 2280 m), con suelo fluvisol móllico (Govaerts et al., 2008) clima templado subhúmedo C (W0), temperatura y precipitación media anual de 15.5 °C y 644 mm (García, 1987).). Los árboles se estructuraron según el método Tatura Modificado y la huerta se plantó en el sistema milpa intercalada con árboles frutales (MIAF) (Santiago-Mejía et al., 2015). Las prácticas culturales fueron: fertilización 60-15-60 (g de N-P2O5-K2O) y 3 kg de pollinaza por árbol, en abril (1/3; 1-1-0 de N, P, K, pollinaza) y agosto (2/3; 0-0-1 de N, P, K, pollinaza); riego por gravedad sin restricción de humedad.
Muestreo y diseño experimental
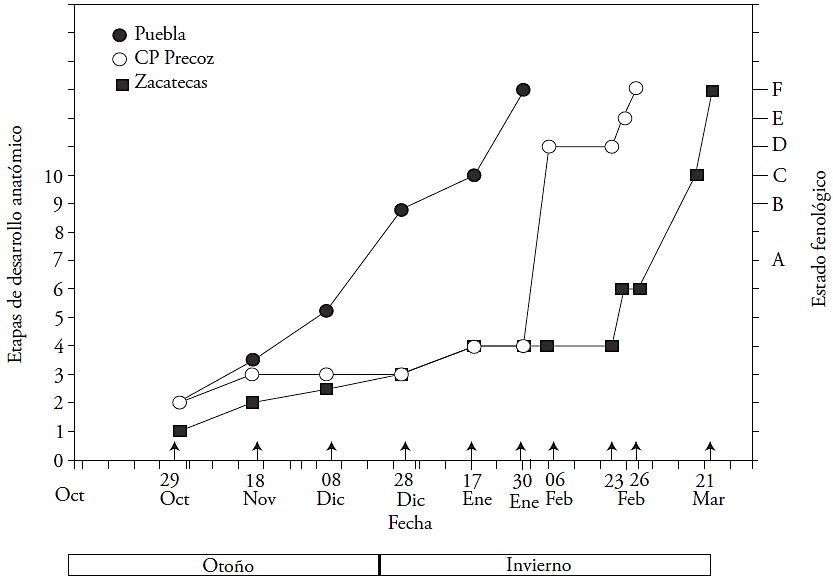
El muestreo se realizó completamente al azar y se tomaron dos yemas florales centrales de cinco ramillas mixtas de un año, de la parte media del dosel de 10 árboles por cultivar. Con la finalidad de cubrir la fenología de los tres cultivares se realizaron siguientes muestreos: 29 de octubre, 18 de noviembre, 08 de diciembre y 28 de diciembre de 2011 y 17 de enero, 30 de enero, 06 de febrero, 23 de febrero, 26 de febrero, 01, 17 y 21 de marzo de 2012. Los caracteres anatómicos revisados fueron cualitativos, por lo cual no se realizó un análisis estadístico. Los resultados son representativos de todas las yemas analizadas en cada estado fenológico.
Estados fenológicos y etapas anatómicas
Las yemas florales se clasificaron en los EF de acuerdo con la nomenclatura de Baggiolini (1952): A: “yema en reposo”; B: “yema hinchada”; C: “cáliz visible”; D: “punta rosada”; y E: “estambres visibles”. Para cada EF se describieron los cambios anatómicos, los cuales se agruparon en “etapas anatómicas” de acuerdo con la nomenclatura sugerida por Reinoso et al. (2002). Siete etapas anatómicas (1-7) se describen en el EF “A”, dos etapas anatómica (8 y 9) en el EF “B”, una etapa anatómica (10) en el EF “C”, y en la EF “D” y ”F” solo se describen los cambios morfológicos, por tratarse de la emergencia de las anteras del botón floral y de la apertura floral o antesis. Las características morfológicas de estos estados fenológicos se ilustran en la Figura 1.

Microtecnia para la anatomía de las yemas
La morfología externa de las yemas, en cada estado fenológico, se documentó con una cámara Moticam 2000® (Motic, China), acoplada a un estéreo-microscopio (Nikon SMZ800®, Japón). Las yemas se fijaron en FAA (10 % formalina, 50 % etanol, 5 % ácido acético glacial, 35 % agua desionizada) durante 48 h y se procesaron para inclusión en Paraplast. Cortes longitudinales (micrótomo rotatorio, American Optical, USA) se tiñeron con Safranina O y Verde fijo FCF. La Safranina O es un colorante catiónico (básico) que tiñe en rojo las paredes lignificadas y ácidos nucleicos, por lo cual se usó para identificar los vasos maduros del xilema. El Verde fijo FCF es un colorante aniónico (ácido) verde que tiñe la celulosa, citoplasma, elementos de tubo criboso, parénquima y células meristemáticas, que en combinación con la Safranina, produce una tinción metacromatica (Baker, 1966). La anatomía de las yemas se estudió con un microscopio Axioskop 2 plus® (Carl Zeiss, Alemania) y una cámara Axiocam 5 MRc (Carl Zeiss, Alemania).
Vascularización y distribución de granos de almidón
La aparición de vasos de xilema en el receptáculo, cáliz, corola, anteras y gineceo se registró con un microscopio Axioskop 2 plus® (Carl Zeiss; Alemania) en campo claro. Los granos de almidón se visualizaron con epifluorescencia (excitación BP 450490 nm y emisión LP 515 nm) y las imágenes se capturaron con la cámara Axiocam 5 MRc (Carl Zeiss, Alemania).
Resultados y discusión
Fenología de la floración
El primer cultivar en salir del “reposo” (A) fue Puebla (finales de diciembre), después CP Precoz (a finales de enero) y al final Zacatecas (finales de febrero). El desarrollo de la yema fue rápido en el cv. Puebla y en enero presento sépalos visibles (EF “C”) (Figura 2). El desarrollo de la yema del cv. CP Precoz fue más lento, el cáliz fue visible (EF “D”) posterior al del cv. Puebla. El cultivar más tardío fue el Zacatecas; la yema permaneció en reposo (EF “A”) hasta febrero, luego se desarrolló rápidamente presentando la punta rosada (EF“D”) en marzo.
Vascularización de la yema floral
La diferenciación del xilema, en la yema floral, fue acrópeta (la maduración de los elementos de vaso inició en la base y avanzó hacia la punta de la yema) en los tres cultivares. Durante el reposo (A1 a A5), la yema de los tres cultivares no está vascularizada. El cv. Puebla fue el primero (28 diciembre) en presentar xilema en el receptáculo, cáliz y corola (A6 y A7) (Figura 2 y 3). En contraste, la ausencia de vascularización se prolongó en CP precoz (30 de enero) y en Zacatecas (23 febrero) (Figura 2 y 3A). La vascularización de la yema inició a los 2 meses en el cv. Puebla, a los 3 meses en el cv. CP Precoz y a los 4 meses en el cv. Zacatecas (Figura 2 y 3B). En el receptáculo, solo células procambiales vasculares fueron observadas (Figura 4A). En las etapas A3, A 4 y A5, el xilema se observó en las escamas basales (Figura 3A). La vascularización de las anteras y ovario se presentó primero en el cv. Puebla (yema hinchada B8 y B9) el 17 enero, y el más tardío (14 marzo) fue Zacatecas (Figura 2 y Figura 3B). La vascularización del estilo presentó la mayor variación entre cultivares: en el cv. Puebla sucedió en 15 d (del 17 al 30 de enero), en CP Precoz en 27 d (del 30 enero al 26 de febrero), y en el cv. Zacatecas en 4 d (del 17 al 21 de marzo).

Figura 3 Distribución y abundancia de los granos de almidón y xilema en los verticilos vegetativos y reproductivos de la yema floral en los estados fenológicos: reposo A); yema hinchada B); cáliz visible C); punta rosada D); cáliz visible E). Los puntos rojos indican los granos de almidón y las líneas azules punteadas indican vasos del xilema.
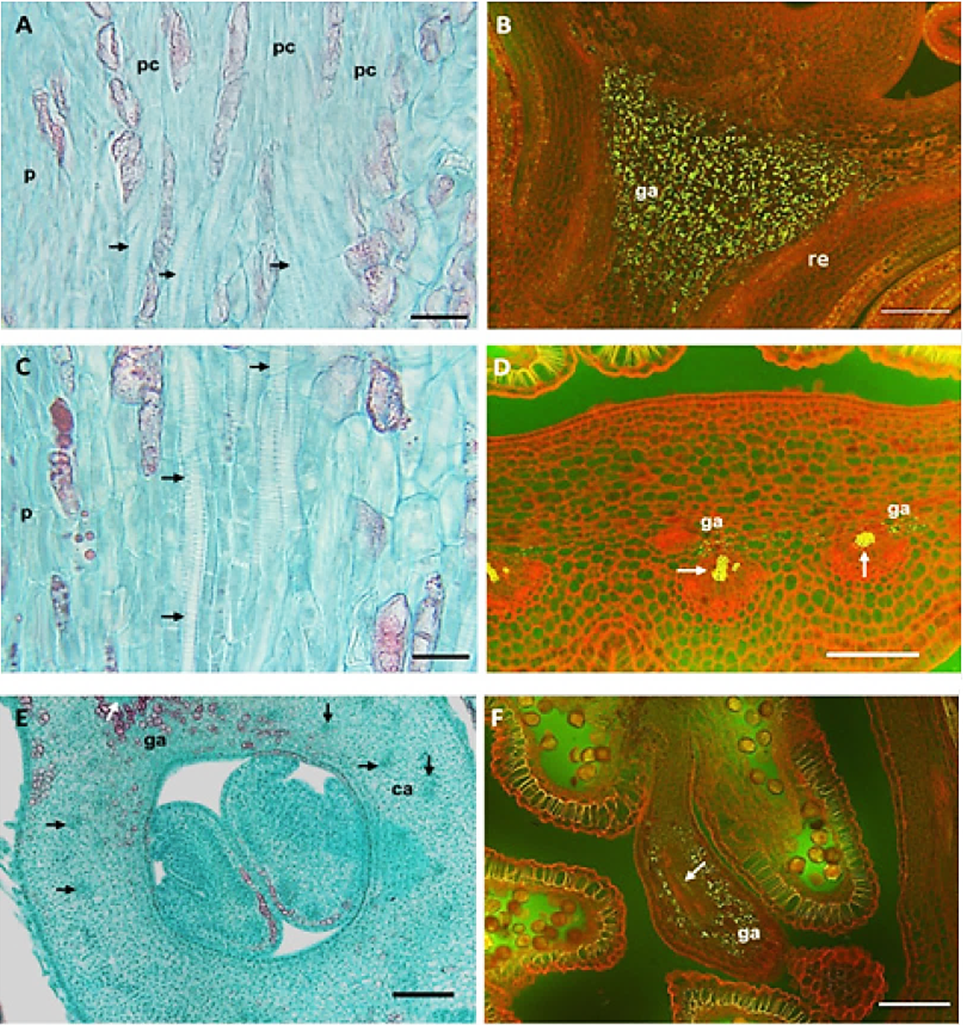
Figura 4 Anatomía de la vascularización y presencia de granos de almidón en yemas florales de duraznero. A) Procambium en el receptáculo, ausencia de xilema en reposo (etapas 3 a 5, cv. Zacatecas); B) granos de almidón en el receptáculo en reposo (etapas 6 a 7, cv. Puebla); C) vasos de xilema en los verticilos vegetativos de yema hinchada (etapa 8, cv CP Precoz); D) xilema y almidón en la base de anteras de yema hinchada (etapa 8, CP Precoz); E) xilema en la pared del carpelo, yema cáliz visible (etapa 10, cv. Puebla); F) vasos y granos de almidón en la antera (etapa 10, cv. Puebla). A, C y E) campo claro; B, D y F) epifluorescencia. Las flechas indican haces vasculares (ga: granos de almidón; re: receptáculo; pc: procambium; ca: carpelo). Barras: A y C = 20 µm; B, D y F = 100 µm; E = 500 µm.
La vascularización de la yema a finales de reposo se reportó para P. armeniaca, P. avium, P. ceratus, P. saliciana y P. sargentii (Ashworth, 1984; Reinoso et al., 2002). Pero nuestro estudio es el primero en describir el avance de la vascularización en las partes de la yema, así como la comparación entre cultivares con diferentes fechas de floración o precocidad.
La vascularización de los verticilos vegetativos, estuvo asociada con un aumento del almidón en el receptáculo (Figura3) en los tres cultivares. Los granos de almidón fueron visibles con epifluorescencia (Figura 4B) y los elementos de vaso del xilema presentaron engrosamiento y placas de perforación escalariformes (Figura 4C). Según Ashworth (1984), en chabacano la conexión vascular entre la ramilla y toda la yema ocurre al final del reposo. En contraste, en los cv. Mexicanos de duraznero este evento fue gradual como se muestra en la Figura 3: el xilema aparece primero en el receptáculo (A-5), luego en el cáliz y la corola (A-7), después el xilema se extendió a las anteras en la yema hinchada (B-8), continuó hacia el ovario (B-9) y alcanzó el estilo y el estigma hasta el estado de cáliz visible (C-10).
Reinoso et al. (2002) solo mencionaron la presencia de tejido vascular en todos los verticilos al final del reposo sin incluir los detalles anatómicos y fenológicos. La Figura 4D muestra haces vasculares y granos de almidón en la base de anteras de yema hinchada (B-8). La pared del ovario se vascularizó (B-9) a finales de yema hinchada (Figura 3). Estos eventos sucedieron primero (inicio de enero) en el cv. Puebla, luego el CP Precoz (febrero) y finalmente en el cv. Zacatecas (principios de marzo). En contraste, en chabacano este evento sucedió antes, a finales del reposo (Julian et al., 2011). En cáliz visible (C-10) los haces de xilema avanzaron hacia el estilo hasta llegar al estigma (Figura 3). Los filamentos de las anteras presentaron un haz central anficribal. Los haces vasculares fueron de tipo colateral y abundantes en la pared del carpelo (Figura 4 E). La vascularización completa del pistilo (C-10), sucedió primero en el cv. Puebla (mediados de enero), luego en cv. CP Precoz (inicios de febrero) y al final en Zacatecas (mediados de marzo).
El evento más tardío fue la vascularización del óvulo, lo que explica porque el primordio de la semilla es la estructura que requiere protección del frío, por razones reproductivas. Durante el año del experimento las heladas se presentaron cuando las yemas florales se encontraban en reposo (Santiago-Mejía et al., 2015), y nuestro estudio evidenció la ausencia de conexión vascular en este estado.
Distribución de granos de almidón
La presencia y distribución de los granos de almidón estuvo asociada con la diferenciación del xilema y desarrollo de los verticilos florales. En el cv. Puebla los granos de almidón aparecieron primero en la base del receptáculo (08 diciembre) y luego aumentaron en número cuando las anteras iniciaron su desarrollo (Figura 4B). En el cv. CP Precoz este evento sucedió hasta el 23 febrero, después el almidón se presentó en la base de las anteras, mientras los granos de polen formaron la célula generativa y la vegetativa (Figura 5G) (Cuadro 1). En yema hinchada (B-8 y B-9) granos de almidón aparecen en el estilo y las anteras; lo cual coincidió con la diferenciación del xilema en el ovario, el inicio de la nucela, y la formación de la pared del polen (Cuadro 1 y Figura 5I). La diferenciación de xilema en los estambres y durante la formación de los granos de polen en los cv. Puebla y CP Precoz fue similar a lo observado en P. armeniaca, aunque la presencia del almidón en las células intermedias ocurrió antes de la formación de microsporas (Julian et al., 2011).
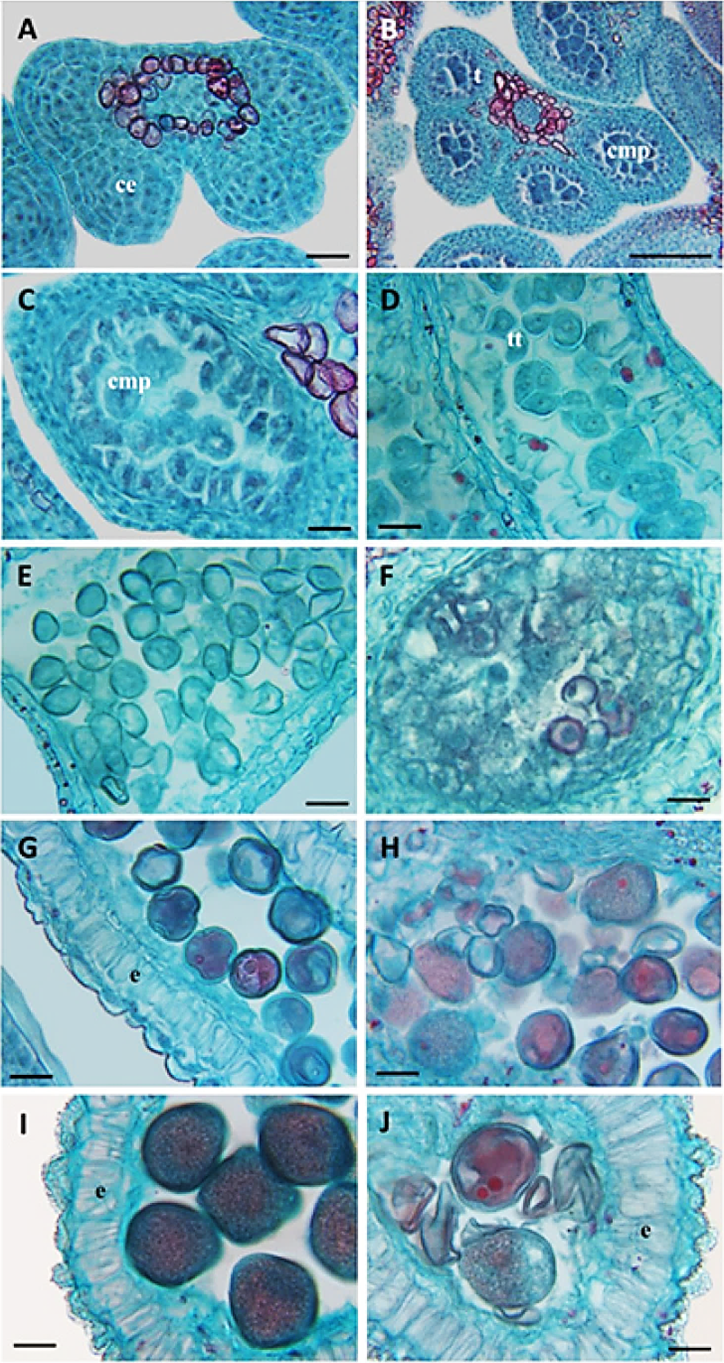
Figura 5 Desarrollo de la antera en tres cultivares de Prunus persica. A) Antera tetraesporangiada con células esporógenas (2n) cv. Puebla; B) células madres del polen (CMP) y tapete diferenciado cv. CP Precoz; C) meiosis de las células madres del polen (1n) Zacatecas; D) formación de tétradas con arreglo tetraédrico cv. Puebla; E) liberación de las microsporas (1n) cv. CP Precoz; F) defecto en la liberación de las microsporas y aborto cv. Zacatecas; G) alargamiento del endotecio y primera mitosis del grano de polen cv. Puebla; H) aborto de granos de polen cv. Zacatecas; I) engrosamiento del endotecio y de la pared del polen cv. Puebla; J) colapso de granos de polen, falta de engrosamiento de la pared del polen cv. Zacatecas. ce: células esporógenas; e: endotecio; cmp: células madres de polen; tt: tétradas. Barras: A, C-J = 20 µm, B = 100 µm.
Cuadro 1 Caracterización anatómica de los estados fenológicos de las yemas florales de duraznero (Prunus pérsica), desde “yema en reposo” (A) hasta “estambres visibles” (E), observados en los cultivares Puebla, CP Precoz y Zacatecas.
| Estado fenológico† | Etapa anatómica ¶ | Caracterización Anatómica |
| A: Yema en “reposo” | 1 | No existe vascularización de la yema (Figura 3 A), presencia de algunos granos de almidón en la base del receptáculo. Anteras jóvenes tretra-esporangiada con dos lóculos en cada uno de los lóbulos unidos por el tejido conectivo (Figura 5A). En el gineceo solo se observa la cavidad locular. |
| 2 | No hay vascularización de la yema. Epidermis diferenciada en la pared del microesporangio y en el interior células esporógenas grandes con citoplasma denso, núcleos grandes y nucléolos prominentes (Figura 5B). Algunos granos de almidón en el receptáculo (Figura 3 A 1-2). | |
| 3 | No hay vascularización de la yema, se identifican algunos elementos de vaso en la base del receptáculo, y aumentan los granos de almidón en la base del receptáculo (Figura 3 A 3-5). Estratos identificables del microesporangio, epidermis, endotecio, tres estratos de células parietales, y el tapete. Inicia la diferenciación de las células madre de polen (CMP, 2n) (Figura 5 C). | |
| 4 | Presencia de tejido vascular llegando al centro del receptáculo (Figura 3 A 3-5). La pared interna del microesporangio se diferencia en tapete, con células de citoplasma denso que rodean a las CMP en desarrollo. Inicio de protuberancias meristemáticas del óvulo en la cavidad locular. | |
| 5 | Tejido vascular en el receptáculo (Figura 3A 3-5). Células del tapete con grandes vacuolas, algunas binucleadas. Las CMP agrandadas comienzan a separarse una de otra. | |
| 6 | El tejido vascular se comienza a diferenciar en los verticilos vegetativos, aumentan los granos de almidón en el receptáculo y éste crece arriba del ovario (flor periginia) (Figura 3 C), En el ovario se diferencia el obturador sobre cada óvulo (sólo ‘Puebla’). Meiosis asincrónica, algunas CMP inician la meiosis (Figura 5 C), pero en otras anteras las CMP forman tétradas (1n) (Figura 5 D). | |
| 7 | El tejido vascular avanza hacia los verticilos de cáliz y corola (Figura 3 A 6-7). Las microsporas (1n) recién se liberan de las tétradas (Figura 5 E). El tapete degenera gradualmente, las células parietales comienzan a comprimir al endotecio. | |
| B: Yema hinchada | 8 | Los granos de almidón aparecen en la base de las anteras y el tejido vascular se extiende a las anteras (Figura 4 F) y aparece almidón cerca del óvulo (Figura 3 B 8). Expansión de las células del endotecio y granos de polen en mitosis (Fig. 5 G), una célula generativa y una célula vegetativa. Se observan nucela del óvulo y obturador en CP Precoz y Zacatecas.. |
| 9 | Aumentan los granos de almidón en el pistilo y anteras, pero disminuyen en la base del receptáculo (Figura 3 B-9). La vascularización abarca el ovario (Figura 3 E). La nucela de los óvulos es prominente. Estratos intermedios de la antera comprimidos y los granos de polen forman una exina gruesa (Figura 5 I) | |
| C: Cáliz visible | 10 | Los granos de almidón disminuyen en toda la yema, pero su vascularización es completa, el xilema avanza hasta el estigma (Figura 3 C-10). En el ovario, los óvulos continúan su desarrollo. La pared del endotecio desarrolla bandas de engrosamiento de pared secundaria en la parte anticlinal y tangencial. En el estigma se inicia la diferenciación de una epidermis con papilas secretoras. |
| D: Punta Rosada | Granos de almidón ausente en las anteras y persistente en el pistilo (Figura 3 D). Grano de polen maduro. El ovulo continua su desarrollo. La corola se observa como un círculo rosa pequeño en el ápice de la yema (Figura 1 Punta rosada), los sépalos adquirieren su color verde-marrón. Adelgazamiento en el estomio. | |
| E: Estambres visibles | Granos de almidón muy escasos en el pistilo y base del receptáculo (Figura 3 E). Los pétalos duplican su longitud, las flores jóvenes se abren parcialmente, los estambres se hacen visibles hacia el exterior de la flor (Figura 1 E), el estomio se rompe. El estigma ha desarrollado una cubierta con papilas secretoras. |
No se incluye el estado fenológico de antesis por no presentar cambios anatómicos relevantes para este estudio.
† La nomenclatura usada de los estados fenológicos es según Baggliolini (1952).
¶ La nomenclatura usada de las etapas anatómicas es según Reinoso et al. (2002).
En el estado de cáliz visible (C-10), de los tres cultivares, hubo disminución de los granos de almidón en la base de la yema y aumento en el pistilo, evento coordinado con la diferenciación del xilema en el estilo. Sin embargo, en punta rosada (D), el almidón desapareció de las anteras, aunque persistió en el ovario (Figura 4). Este patrón fue similar al observado en inflorescencias de olivo (De la Rosa y Rallo, 2000). Los cambios en la partición de almidón durante el desarrollo de las yemas florales se han demostrado en Prunus persica (Bonhomme et al., 2005; González-Rossia et al., 2008), Rubus idaeus (Palonen, 1999), Pyrus pyrifolia (Ito et al., 2002), Populus x canadensis (Elle y Sauter, 2000) y Olea europea (De la Rosa y Rallo, 2000), pero nuestro reporte es el primero sobre la localización histológica del almidón en la yema floral asociada con la morfogénesis y diferenciación del xilema en duraznero.
Caracterización anatómica con énfasis en desarrollo de antera
Los cambios anatómicos observados en la yema en el estado fenológico de reposo (A) se agruparon en siete etapas anatómicas (1-7) las cuales se resumen en el Cuadro 1. Estos eventos sucedieron de manera desfasada en el tiempo entre cultivares (Figura 2). Los cambios anatómicos observados durante las primeras cuatro etapas anatómicas del reposo (A-1 a A-4), sucedieron lentamente en la yema floral del cv Zacatecas (4 meses), fueron menos lentos en el cv. CP Precoz (3 meses) y rápidos en el cv. Puebla en 20 d (Figura 2 y Cuadro 1).
En los tres cultivares, durante el “reposo” (A), el desarrollo temprano del polen, fue normal y similar; el microesporangio presentó células esporógenas (2n) (Cuadro 1 y Figura 5A); en la etapa A-2 se diferenciaron las células madres de polen (2n) (Cuadro 1); en las etapas A-3 y A-4 el tapete fue prominente (Cuadro 1 y Figura 5C); en la etapa A-6 se observaron las células madres del polen (CMP) en meiosis y las microesporas (1n) se organizaron en un arreglo tetraédrico (Figura 5D). En yema hinchada (B8 y B-9), del cv Puebla y CP Precoz, las microsporas se separaron en granos de polen jóvenes (1n) (Cuadro 1 y Figura 5E), pero en el cv. Zacatecas las microsporas no se separaron, y algunas se obliteraron y desintegraron (Figura 5F). Esto significó un defecto en la formación del grano de polen en el cv. Zacatecas. Después, los cv Puebla y CP Precoz continuaron su desarrollo normal, aunque desfasado en el tiempo; el cv Puebla presentó primero la primera mitosis del grano de polen joven y el engrosamiento de la pared del polen (Figura 5G), y el cv. Zacatecas mostró granos de polen obliterados con deficiente engrosamiento de la exina (Figura 5H).
En el siguiente estado, cáliz visible (10-C), las paredes anticlinales del endotecio se lignificaron de manera normal en los tres cultivares, pero la maduración del grano de polen solo se presentó en los cv. Puebla y CP Precoz (Figura 5I).
En el estado de punta rosada (D) varias microsporas detuvieron su desarrollo y abortaron en el cv. Zacatecas (Figura 5H). El cv. Zacatecas presentó la mayoría de los granos de polen abortivos y con defectos en la exina (Figura 5J).
Las anormalidades y aborto del polen se han observado cuando los requerimientos de frío son insatisfechos (Bonhomme et al., 2005; Nava et al., 2009; Reig et al., 2013). En el contexto del calentamiento global los frutales de clima templado enfrentarán reducción en horas frío, problemas durante el “reposo” asociados con aborto y reducción en el flujo de carbohidratos hacia las yemas (Campoy et al., 2011; Marafon et al., 2011). De acuerdo con el método de Richardson et al. (1974), el cv. Zacatecas requiere 550 Unidades Frío (UF), el CP Precoz 400 UF y el Puebla 350 UF (Santiago-Mejía et al., 2015). El frío acumulado promedio del 17 noviembre al 31 de marzo en la región de estudio, fue menor a las 550 UF (Santiago-Mejía et al., 2015). En forma natural el cv. Zacatecas se encuentra a mayor latitud y acumula más horas frío (Rumayor et al., 2009). La deficiencia de frío provoca anomalías en el gametofito masculino como; aborto de polen, reducción de la producción y problemas de germinación del polen (Nava et al., 2009), además de una apertura floral anormal (Bonhomme et al., 2005). Lo anterior explica las anomalías observadas en el desarrollo del polen de Zacatecas.
Conclusiones
La diferenciación del xilema, en la yema floral del duraznero es acrópeta (la maduración de los elementos de vaso inicia en la base y avanza hacia la punta de la yema) y gradual. No hay indicios de vascularización durante el reposo temprano en los tres cultivares estudiados (Puebla, CP Precóz y Zacatecas). La presencia de almidón precedió a la aparición de xilema. La vascularización de los verticilos vegetativos inició en el reposo tardío: a los dos meses en el cv. Puebla, a los tres meses en el cv. CP Precoz y a los cuatro meses y medio en el cv. Zacatecas. La vascularización avanza hasta el ovario en el estado de yema hinchada y continúa hacia el estigma hasta cáliz visible. La vascularización del estilo fue el evento que presentó la mayor variación entre cultivares: en el cv. Puebla15 días, en CP Precoz 27 días, y en el cv. Zacatecas 4 días. La vascularización fue postergada de acuerdo con la precocidad del cultivar. La vascularización tardía de la parte femenina, sería una estrategia para proteger su función reproductiva en presencia de heladas. Esta información se puede usar en programas de manejo agronómico y reducir los daños por heladas en durazno.

