











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Existen alrededor de 200 especies anuales y perennes del género Lotus. El mediterráneo europeo es la zona con mayor diversidad (Lagler, 2003) y L. corniculatus, la especie con importancia forrajera mayor (Castro, 2006), está en regiones templadas de Europa, Asia Menor y norte de Sudamérica (Gunn et al., 1992). Esta leguminosa es de clima templado, su rendimiento y calidad nutricional (18 a 22 % de proteína cruda) se compara con Medicago sativa L., y Trifolium repens L., (Marley et al., 2006); además, contiene menos celulosa y más carbohidratos no estructurales (Grant, 2009) y no produce timpanismo en los animales en pastoreo (Beuselinck y Grant, 1995).
La productividad y persistencia de las praderas está en función de su capacidad para recuperar y mantener el área foliar remanente, por lo cual, diferentes estrategias de cosecha tienen impacto sobre las plantas y determinan la velocidad de crecimiento, la productividad y la persistencia (Nabinger y Pontes, 2001). Por lo tanto, la eficiencia de la cosecha es uno de los principales factores que definen la productividad de una pradera (Giacomini et al., 2009), en la cual influyen las condiciones ambientales como la cantidad de luz (Lemare y Agnusdei, 2000) que aumenta la competencia entre plantas. En los periodos de rebrote se reduce la cantidad y calidad luminosa en la pradera (Da Silva y Nascimento, 2007), por lo que un criterio para estimar el punto óptimo de cosecha es cuando la pradera alcanza 95 % de luz interceptada (Montagner et al., 2012); en consecuencia, un buen manejo de la cantidad de luz interceptada por la pradera asegura una mejor productividad (Difante et al., 2008). La información para L. cornicultatus es escasa, mientras que en otras leguminosas de clima templado hay correlaciones altas entre el rendimiento de forraje con la cantidad interceptada de luz (Rojas et al., 2016). Por lo tanto, el objetivo del presente estudio fue evaluar el comportamiento productivo del genotipo 202700 de L. corniculatus en función de la cantidad de luz interceptada y un intervalo de corte fijo definido estacionalmente.
Materiales y métodos
Condiciones de campo
El experimento se realizó en el Colegio de Postgraduados, Campus Montecillo, Texcoco, México (19° 29’ N y 98° 53’ O), a 2,250 msnm, en los ciclos agrícolas de 2014-2015 y 2015-2016. El clima es templado sub-húmedo, con temperatura media de 15 °C y precipitación anual acumulada de 636 mm (García, 2004). El suelo es de textura franco arenosa, con pH de 7.1. Los datos climáticos se obtuvieron de la estación meteorológica de la Universidad Autónoma Chapingo, a 2 km del sitio de estudio. La precipitación anual acumulada fue 1,043 mm en el primer año y 877 mm en el segundo; durante primavera y verano se registraron las máximas precipitaciones y temperaturas, respecto a otoño e invierno, en ambos años (Figura 1).
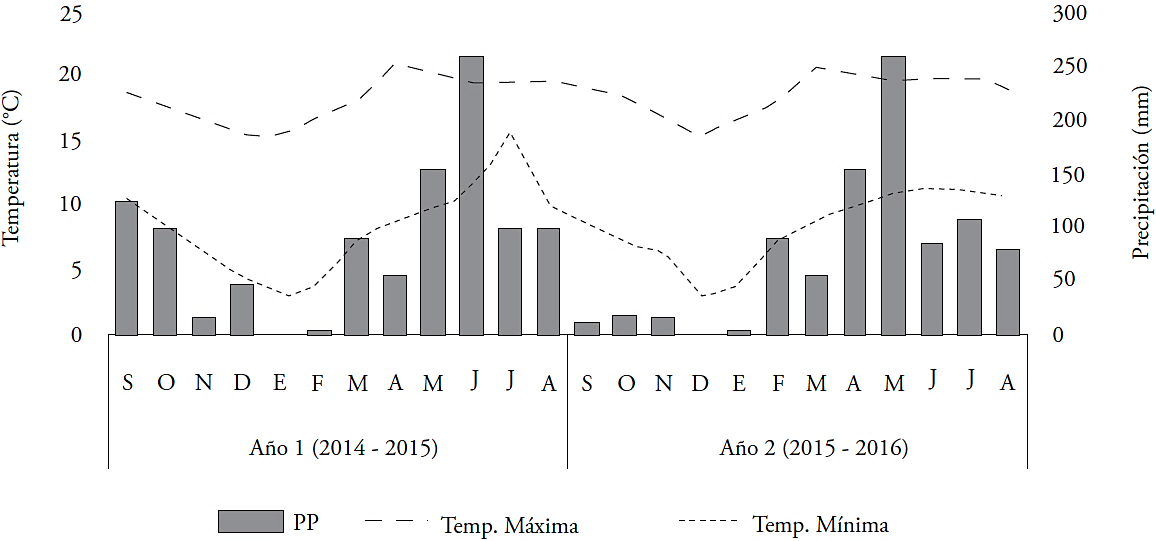
Figura 1 Temperatura media mensual máxima, mínima y precipitación pluvial (PP) acumulada mensual en los dos ciclos agrículas.
La pradera de L. corniculatus se estableció mediante trasplante el 14 de marzo de 2014, la densidad fue 9 plantas m-2 y la distancia entre plantas fue 33 cm. Las plantas se obtuvieron de plántulas ya establecidas en invernadero, en sustrato de suelo típico del lugar, sin fertilizar. Antes de iniciar el experimento, hubo un pastoreo con ovinos a las 13 semanas de su establecimiento y un corte de uniformización al inicio del experimento (5 de septiembre de 2014) a 7 cm sobre el nivel del suelo. Las parcelas se regaron cada 15 d a capacidad de campo en época de estiaje; no se aplicó fertilizante.
El área experimental se dividió en 12 sub parcelas de 4 m2 de área, con 9 plantas por m2 y 36 plantas por unidad experimental. Los cortes fueron los tratamientos y se determinaron cuando las praderas alcanzaron porcentajes de luz interceptada de 90, 95 y 100 %, más un corte fijo definido estacionalmente (primavera - verano 28 d, otoño 35 d, e invierno cada 42 d). La LI fue monitoreada a nivel del suelo (seis mediciones) y se usó un analizador de dosel AccuPAR Linear PAR/LAI ceptometer Modelo PAR 80 (Decagon devices, USA).
El rendimiento de forraje (kg MS ha-1) se evaluó al cosechar la biomasa en dos cuadros fijos (0.25 m2) por repetición, establecidos al inicio del experimento y el forraje se secó a 60 °C, hasta peso constante, en una estufa de aire forzado (Felisa, Mod. FE243A). La composición botánica y morfológica (CBM) se calculó al tomar una muestra de 10 % del total del forraje cosechado, separado en hoja, tallo, material muerto (material senescente) y maleza; se calculó el aporte al rendimiento total de cada componente (kg MS ha-1). La altura de la planta (cm) se midió en 12 lecturas al azar por repetición y se usó una regla graduada de 50 cm. La equivalencia en rendimiento por cm de altura de la planta se calculó dividiendo el rendimiento entre los cm de altura de la planta entre el número de cortes. Los datos obtenidos de la CBM de hoja y tallo se usaron para determinar la relación hoja/tallo (R:H/T: 1), dividiendo el peso de la hoja entre el tallo.
Los tratamientos se asignaron a unidades experimentales de acuerdo con un diseño de bloques al azar en parcelas divididas con tres repeticiones, donde la parcela grande fue el genotipo y la chica el tratamiento. Los datos obtenidos por corte se organizaron por estación y se analizaron con PROC GLM de SAS (SAS Institute, 2009); los promedios se compararon con la prueba de Tukey (p≤0.05).
Resultados y discusión
Rendimiento de forraje
Solo en el segundo año hubo diferencias (p≤0.05) en la acumulación anual de forraje (Cuadro 1) y el rendimiento menor correspondió al corte fijo con 29 % menos respecto al promedio de los tres porcentajes de luz interceptada (32,173 kg MS ha-1), lo que indica menor adaptación de la especie al manejo de corte fijo, respecto a cortes dependientes del porcentaje de luz interceptada. Independiente del tratamiento, los mayores rendimientos medios estacionales, en ambos años, se presentaron en primavera con 10,461 kg MS ha-1 y los menores en otoño (5,915 kg MS ha-1) e invierno (5,585 kg MS ha-1). Tal comportamiento puede explicarse por la relación directa y estrecha entre el rendimiento del forraje y las temperaturas óptimas (22 °C) para el crecimiento de la especie en condiciones de campo (García et al., 2014). Las temperaturas óptimas se presentaron en primavera (Figura 1), lo que benefició el crecimiento y productividad de L. corniculatus. En nuestro estudio, el comportamiento estacional es similar al rendimiento de forraje reportado por Scheffer et al. (2011) de cinco poblaciones de L. corniculatus, pero a diferencia en el comportamiento del corte fijo establecido en nuestro estudio, esos autores observaron que una alta frecuencia de corte aumenta el rendimiento. Además, Sbrissia et al. (2010) atribuyen estos cambios en el rendimiento a las condiciones ambientales en cada estación del año. Díaz et al. (1996) encontraron una distribución estacional en la producción de forraje de L. corniculatus de 49, 25, 14 y 12 % para primavera, verano, invierno y otoño, respectivamente, con 72 % en primavera-verano. Según Santiñaque y Battista (2003), hay un aumento de 45 % en los intervalos de cosecha de 44 d, comparado con 22 d. Para un intervalo de corte de 45 d, García et al. (2014) observaron un rendimiento promedio de forraje de 7700 kg MS ha-1, y el genotipo 202700 fue el más productivo de los 12 genotipos de L. corniculatus evaluados; también señalaron que la producción acumulada puede estar influenciada por el clima, manejo y el hábito de crecimiento del genotipo.
Cuadro 1 Rendimiento acumulado de forraje de L. corniculatus, genotipo 202700, en función del porcentaje de luz interceptada (LI) y un corte fijo definido estacionalmente.
| LI (%) | Otoño | Invierno | Primavera | Verano | Anual acumulado | EEM† |
| Año 1 (2014 - 2015) | ||||||
| 90 | 4881 Ad | 6447 Ac | 10396 ABa | 8593 ABb | 30317 A | 516 |
| 95 | 5533 Ad | 6757 Ac | 10806 Aa | 9177 Ab | 32274 A | 393 |
| 100 | 5785 Ab | 6453 Ab | 11116 Aa | 9880 Aa | 33234 A | 823 |
| Corte fijo§ | 4896 Ac | 6568 Ab | 8708 Ba | 7509 Bb | 27680 A | 373 |
| Promedio estacional | 5274 d | 6556 c | 10256 a | 8790 b | 30876 | 369 |
| EEM† | 721 | 864 | 673 | 546 | 2303 | |
| Año 2 (2015 - 2016) | ||||||
| 90 | 5695 ABc | 5729 Ac | 11063 Aa | 8797 Ab | 31283 A | 549 |
| 95 | 6567 Ac | 6293 Ac | 12068 Aa | 8773 Ab | 33701 A | 265 |
| 100 | 5398 ABc | 5688 Ac | 11433 Aa | 9015 Ab | 31534 A | 762 |
| Corte fijo§ | 4934 Bb | 4377 Bb | 8222 Ba | 5311 Bb | 22844 B | 562 |
| Promedio estacional | 5648 c | 5522 c | 10696 a | 7974 b | 29841 | 259 |
| EEM† | 531 | 421 | 694 | 555 | 1003 | |
AB,abcd Promedios con letra mayúscula diferente en una columna y letra minúscula diferente en una hilera, son estadísticamente diferentes (p≤0.05).
†EEM=error estándar de la media. §Corte fijo=otoño: 35, invierno: 42, y primavera-verano 28 días entre corte.
El rendimiento acumulado de forraje de L. corniculatus en función de los porcentajes de LI no tuvo diferencias (p>0.05), en contraste, Da Silva y Nascimento, (2007), Da Silva y Hernández, (2010); Mattera et al. (2013) indicaron que las especies forrajeras se deben cosechar al 95 % de intercepción de luz, siempre y cuando la densidad de plantas sea competitiva. Al respecto, Rojas et al. (2016) en alfalfa (M. sativa L.) registraron el 95 % de luz interceptada sólo en verano, explicada por la baja densidad de plantas, debido al tiempo de establecimiento de la pradera. Además, Mattera et al. (2013) encontraron que el 95 % de intercepción de luz en alfalfase alcanza en primavera al combinar distancias entre plantas de 10, 15, 20, 25 y 30 cm, mientras que en verano e invierno solo se presenta en densidades de 10 y 15 cm.
Composición botánica y morfológica
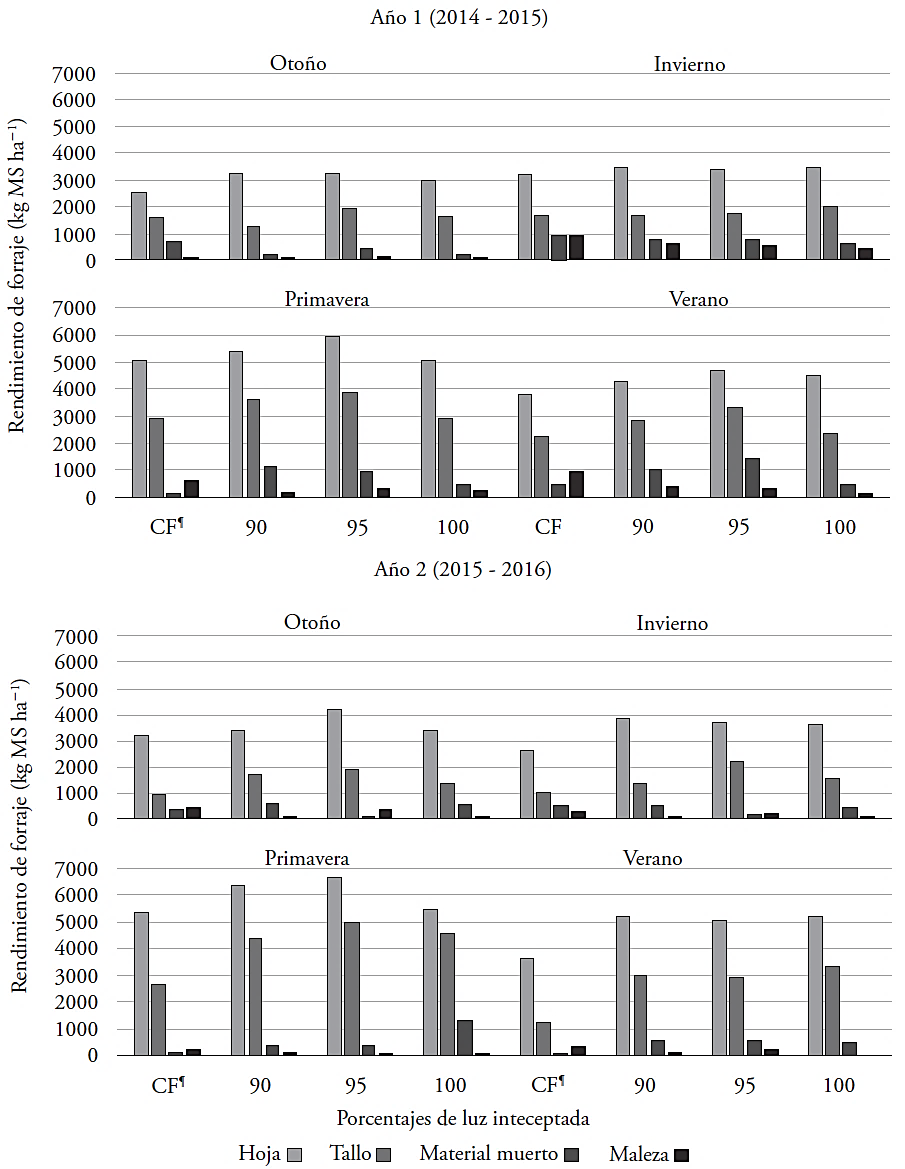
La pradera tuvo un porcentaje mínimo de malezas con promedio anual de 4 % en ambos años (Cuadro 2). Al respecto, Teixeira et al. (2007) mencionaron que la presencia de malezas en una pradera puede aumentar con el tiempo, por efecto de la frecuencia de corte. Dentro de los componentes morfológicos, la hoja aporto más al rendimiento con promedio anual de 17247 kg MS ha-1 (58 %), seguida por el tallo (31 %) y el material muerto (7 %). Tal comportamiento puede resultar del acelerado ritmo de crecimiento de la hoja y el tallo, favorecidos por las condiciones ambientales como el fotoperiodo, la temperatura y la humedad (Difante et al., 2009).
Cuadro 2 Rendimiento anual de forraje, por componente botánico y morfológico, de L. corniculatus, genotipo 202700, en función del porcentaje de luz interceptada (LI) y un corte fijo definido estacionalmente.
| LI (%) | Hoja | Tallo | Material muerto | Maleza | EEM† |
| Año 1 (2014 - 2015) | |||||
| 90 | 16435 BCa | 9389 ABb | 3161 Ab | 1333 Ad | 477 |
| 95 | 18858 Aa | 10418 Ab | 1977 Ac | 1020 Ac | 691 |
| 100 | 17335 Aba | 10919 Ab | 3598 Ac | 1383 Ad | 749 |
| Corte fijo § | 14504 Ca | 8386 Bb | 2208 Ac | 2582 Ac | 1042 |
| Promedio | 16783 a | 9778 b | 2736 c | 1579 c | 430 |
| EEM† | 847 | 669 | 790 | 593 | |
| Año 2 (2015 - 2016) | |||||
| 90 | 18788 Aba | 10387 Ab | 1976 ABc | 132 Ad | 458 |
| 95 | 19622 Aa | 12091 Ab | 1178 Bc | 809 Ac | 1010 |
| 100 | 17674 Ba | 10816 Ab | 2833 Ac | 211 Ad | 477 |
| Corte fijo§ | 14763 Ca | 5794 Bb | 979 Bc | 1308 Ac | 574 |
| Promedio | 17712 a | 9772 b | 1741 c | 615 d | |
| EEM† | 470 | 762 | 379 | 724 | |
ABC,abc Promedios con letra mayúscula diferente en una columna y letra minúscula diferente en una hilera son estadísticamente diferentes (p≤0.05). †EEM=error estándar de la media. §Corte fijo=otoño: 35, invierno: 42, y primavera-verano. 28 d entre corte.
En ambos años, el tratamiento con 95 % de luz interceptada tuvo mayor cantidad de hoja, 19 240 kg MS ha-1 en promedio, y el corte fijo con la menor cantidad de hojas y también de tallos con 14 634 y 7090 kg MS ha-1, respectivamente (p≤0.05) en relación a los otros tratamientos (Cuadro 2). Estos resultados son similares a los indicados por Giacomini et al. (2009), quienes evaluaron B. brizantha cv. Marandu a intercepciones de luz de 95 y 100 % y dos intensidades de corte (10 y 15 cm) y encontraron una mayor cantidad de hoja a 95 % de intercepción luminosa, a una altura de forraje residual de 10 cm. En nuestro estudio, el forraje muerto no fue diferente (p>0.05) entre tratamientos durante el primer año; en el segundo año los valores mayores se registraron a 100 % de luz interceptada, y el menor a 95 % de luz interceptada y corte fijo. Lo anterior indica que el momento óptimo de cosecha es a 95 % de radiación interceptada, por registrar mejor calidad de forraje, basada en una menor cantidad de material muerto, mayor rendimiento (Cuadro 1, año 2) y mayor cantidad de hoja (Cuadro 2), similar a lo reportado por Montagner et al. (2012). Además, la presencia mínima de material muerto o senescente en el corte fijo se puede deber a que la planta no alcanzó su momento fisiológico óptimo de cosecha, y una mayor cantidad de luz penetró al interior de la pradera (Matthew et al., 2000), lo que causó una mayor producción de material verde, hojas y tallos, pero con menor peso de estos últimos (Villegas et al., 2004).
Según la época del año, la hoja fue el componente que más aporto al rendimiento de forraje (Figura 2). El rendimiento estacional mayor y menor se registró durante primavera y otoño-invierno con 5813 y 3,378 kg MS ha-1, el tratamiento con 95 % de luz interceptada y el de corte fijo tuvieron rendimientos de 6480 y 5177 kg MS ha-1. Estos resultados indican efectos de estación y de los tratamientos sobre la producción de hoja, influenciados por las condiciones ambientales durante el estudio (Figura 1). Además, la cantidad de tallos mayor y menor se presentó durante primavera e invierno, con promedios estacionales de 3814 y 1612 kg MS ha-1. Al respecto, Sbrissia et al. (2010) mencionaron que la producción de tallos cambia en función de las variaciones estacionales en la cantidad y calidad de luz que inciden sobre la pradera, la precipitación y la temperatura. En cuanto al material muerto y malezas, no hubo diferencias estacionales en ambos años.
Altura de la planta
Hubo diferencias entre tratamientos aplicados y las estaciones del año (p≤0.05), excepto en el verano del segundo año (p>0.05) (Cuadro 3). Las alturas promedio anuales mayores se registraron en 100 y 95 % de luz interceptada con 23 y 24 cm en el primer y segundo año, mientras que las menores alturas fueron en el corte fijo con 19 cm, en promedio, de ambos años. Estos resultados están relacionados con la edad de la planta porque a mayor porcentaje de luz interceptada la planta tuvo más días de rebrote (51 d, promedio de ambos años1), respecto al corte fijo (33 d, ambos años4 ), el cual siempre fue de edad menor, con cortes más frecuentes y afectó el rendimiento de forraje (Villegas et al., 2006). Al respecto, Santos et al. (2004) señalaron que a una altura mayor corresponde un rendimiento de forraje mayor, pero Scheffer et al. (2011) relacionaron la altura y el rendimiento de L. corniculatus, con el hábito de crecimiento (erectos y postrados).
Cuadro 3 Altura media de la planta de L. corniculatus, genotipo 202700, en función del porcentaje de luz interceptada (LI) y un corte fijo definido estacionalmente.
| LI (%) | Otoño | Invierno | Primavera | Verano | Promedio anual | EEM† | |
| Año 1 (2014 - 2015) | |||||||
| 90 | 22 Ab | 17 Cc | 23 BCab | 25 Aa | 22 B | 0.7 | |
| 95 | 21 Ab | 19 Bb | 25 Ba | 25 Aa | 22 AB | 0.8 | |
| 100 | 16 Bc | 21 Ab | 28 Aa | 26 Aa | 23 A | 0.6 | |
| Corte fijo§ | 22 Aa | 15 Db | 22 Ca | 22 Ba | 20 C | 1.0 | |
| Promedio estacional | 20 b | 18 c | 24 a | 24 a | 22 | 22 | |
| EEM† | 0.6 | 0.3 | 0.7 | 0.6 | 0.2 | ||
| Año 2 (2015 - 2016) | |||||||
| 90 | 21 Aab | 16 Bb | 24 Ba | 23 Aa | 21 B | 1.5 | |
| 95 | 19 ABc | 23 Ab | 30 Aa | 23 Ab | 24 A | 0.7 | |
| 100 | 18 BCc | 23 Ab | 28 ABa | 22 Ab | 23 AB | 0.8 | |
| Corte fijo§ | 17 Cb | 13 Bc | 19 Cb | 23 Aa | 18 C | 1.1 | |
| Promedio estacional | 19 c | 19 c | 25 a | 23 b | 21 | 21 | |
| EEM† | 0.7 | 1.6 | 1.4 | 0.8 | 0.7 | ||
ABC,abc Promedios con letra mayúscula diferente en una columna y letra minúscula diferente en una hilera, son estadísticamente diferentes (p≤0.05). EEM†=error estándar de la media. §Corte fijo=otoño: 35, invierno: 42, y primavera-verano 28 d entre corte.
En el primer año las alturas mayores se registraron en primavera y verano con 24 cm y las menores en invierno con 18 cm; mientras que, en el segundo año, en primavera se presentó la altura mayor con 25 cm, y en otoño e invierno la menor, con 19 cm. Las alturas mayores de la planta se encontraron en las estaciones con las condiciones óptimas de humedad y temperatura para el mayor crecimiento y rendimiento de la especie (Figura 1). Estos resultados son similares a los reportados por García et al. (2014), al evaluar 12 genotipos de L. curniculatus. Además, según Marcelino et al. (2006) con condiciones adecuadas de fotoperiodo, temperatura y humedad, las plantas forrajeras aceleran su crecimiento y presentan cambio en la altura en las diferentes estaciones (Difante et al., 2009), lo cual tiene una correlación positiva con el rendimiento de forraje (Da Silva y Nacimiento Jr., 2007), como sucedió en nuestra investigación.
La altura de la planta puede ser asociada con la producción de forraje (Hakl et al., 2010). Además, la producción de forraje de L. corniculatus se relaciona con la altura, pero también con el hábito de crecimiento del genotipo, con los rendimientos y altura más bajas en genotipos postrados (García et al., 2014). La altura de la pradera da una idea del forraje producido; por lo tanto, al calcular la equivalencia de los mayores rendimientos estacionales promedios y de los tratamientos, cada cm de altura correspondió a 178 kg MS ha-1 para primavera y 184 kg MS ha-1 para el 95 % LI, promedios de ambos años.
Relación hoja: tallo
La relación promedio anual mayor de hoja: tallo se presentó en el corte fijo (Cuadro 4), seguido de 90, 95 y 100 % de intercepción luminosa, en ambos años. El corte fijo superó en 26 % (2.3 vs 1.7: 1) al promedio de los tratamientos con 95 y 100 % de LI, en el primer año y en el segundo año con 39 % a 100% de LI (2.8 vs 1.7:1). En el corte fijo, la relación hoja: tallo mayor fue el resultado de cortes más frecuentes (33 d promedio anual5), lo cual no permitió cosechar la especie en su momento óptimo, sino en la fase de crecimiento acelerado, donde se encuentra el mayor porcentaje de hojas jóvenes y poca cantidad de tallo; en consecuencia, hubo una mayor penetración de luz al interior de la pradera (Barbosa et al., 2011). Además, Montagner et al. (2012) mencionan que, posterior al corte, la calidad y cantidad de luz incidente al interior de la pradera, es alterada por el intervalo de corte que causa variaciones en la producción de hojas y tallos y, por lo tanto, la relación hoja: tallo.
Cuadro 4 Relación hoja: tallo de L. corniculatus, genotipo 202700, en función del porcentaje de luz interceptada y un corte fijo definido estacionalmente.
| LI (%) | Otoño | Invierno | Primavera | Verano | Promedio anual | EEM† | |
| Año 1 (2014 - 2015) | |||||||
| 90 | 2.0 Aa | 2.2 Aa | 1.8 Aa | 2.0 Aa | 2.0 AB | 0.30 | |
| 95 | 1.9 Aab | 2.0 Aa | 1.6 Abc | 1.4 Ac | 1.7 B | 0.14 | |
| 100 | 1.9 Aa | 1.9 Aa | 1.5 Ab | 1.5 Ab | 1.7 B | 0.32 | |
| Corte fijo§ | 2.7 Aa | 2.4 Aa | 2.0 Aa | 2.1 Aa | 2.3 A | 0.47 | |
| Promedio estacional | 2.1 a | 2.1 a | 1.7 b | 1.8 b | 1.9 | 0.09 | |
| EEM† | 0.28 | 0.52 | 0.26 | 0.37 | 0.20 | ||
| Año 2 (2015 - 2016) | |||||||
| 90 | 2.6 Aba | 2.9 Aa | 1.5 Bb | 1.8 Bb | 2.2 B | 0.28 | |
| 95 | 2.4 Ba | 2.3 ABa | 1.3 Bb | 1.7 Bb | 2.0 BC | 0.23 | |
| 100 | 2.1 Ba | 1.7 Bab | 1.2 Bb | 1.6 Bab | 1.7 C | 0.24 | |
| Corte fijo§ | 3.3 Aa | 2.7 Abc | 2.2 Ac | 2.9 Aab | 2.8 A | 0.20 | |
| Promedio estacional | 2.6 a | 2.4 a | 1.6 c | 2.0 b | 2.1 | 0.12 | |
| EEM† | 0.27 | 0.25 | 0.15 | 0.20 | 0.11 | ||
AB,abc Promedios con letra mayúscula diferente en una columna y letra minúscula diferente en un reglón son estadísticamente diferentes (p≤0.05). †EEM=error estándar de la media. §Corte fijo=otoño: 35, invierno: 42, y primavera-verano. 28 d entre corte.
La relación hoja: tallo, entre estaciones, en ambos años, fue diferente (p≤0.05; Cuadro 4). La relación hoja: tallo fue mayor en otoño e invierno con 2.1:1 y 2.5:1 para el primer y segundo años. En primavera se registraron los menores valores de la relación hoja: tallo, en ambos años, y fue diferente al verano solo en el segundo año (p≤0.05). Estos resultados son similares a los reportados por Barbosa et al. (2011), quienes encontraron una relación hoja: tallo mayor en estaciones donde el crecimiento en las plantas es menor (otoño e invierno), debido a una mayor densidad de tallos, pero de menor peso; además, una relación hoja: tallo menor en primavera y verano resulta de un peso individual de tallos mayor (Villegas et al., 2004). Al respecto, Giacomini et al. (2009) mencionan que la translocación de asimilados, de las hojas hacia los tallos, es mayor en primavera y verano, lo cual resulta en un mayor peso de estos. La proporción de hoja respecto al tallo fue mayor en todas las estaciones y tratamientos, con un promedio de 1.9:1 (primer año) y 2.1:1 (segundo año).
Conclusiones
El mejor comportamiento productivo del genotipo 202700 de L. corniculatus se presentó en los porcentajes de luz interceptada, marcando diferencias con el corte fijo, no obstante, con 95 % LI se produjo la mayor cantidad de hoja, principalmente en primavera, por lo que se puede definir este último como la mejor estrategia de manejo. Pero en un corte fijo la especie registró menos rendimiento. La producción de forraje en nuestro estudio es diferente a la observada en otras partes del mundo y se debe investigar bajo los distintos tipos de ambientes en México.

