











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En México, el género Pinus tiene gran importancia ecológica, económica y social, su valor económico es alto porque es fuente de madera, leña, pulpa, resina y semillas, sustenta la industria forestal y proporciona servicios ambientales, ya que influyen en el clima regional (Ramírez-Herrera et al., 2005: Sánchez-González, 2008). La mayoría de sus especies se restringen a determinados rangos geográficos (Woodward, 1987), en los que el clima y el suelo son los factores principales que delimitan su distribución (Dawson y Spannagle, 2009). El cambio en el clima y la actividad antropogénica (Peterson et al., 2006), inducen modificaciones en la fenología, el crecimiento, e incluso en la dinámica poblacional (Parmesan, 2006), así como en los intervalos de distribución de muchas especies (Walther, 2010).
En la literatura científica existen modelos para predecir la distribución de especies (Elith et al., 2006), y son representaciones cartográficas de la capacidad de una especie para ocupar un espacio geográfico, determinado en función de variables continuas o categóricas (Guisan y Zimmermann, 2000) también llamadas bioclimáticas. Las más utilizadas son climáticas (Felicísimo et al., 2012), edafológicas (Cruz-Cárdenas et al., 2016; Ramos-Dorantes et al., 2017) y variables topográficas como altitud, pendiente y exposición (Bradley y Fleishman, 2008; Cruz-Cárdenas et al., 2016). MaxEnt es un software basado en la aproximación estadística Máxima Entropía (MaxEnt) que predice la distribución potencial o hábitat de una especie utilizando datos de presencia (Phillips et al., 2006), es considerado el mejor método (Guisan y Zimmermann, 2000; Benito de Pando y Peñas de Giles, 2007; Kumar y Stohlgren, 2009) al estimar la probabilidad (de 0 a 1) de ocurrencia de las especies (Phillips et al., 2006), con un alto grado de precisión. Esta técnica se aplica para priorizar zonas de conservación biológica (Aguirre y Duivenvoorden, 2010; Ávila et al., 2014), evaluar efectos del cambio climático (González et al., 2010; Felicísimo et al., 2012), analizar patrones de propagación de especies invasoras (Morales, 2012) y modelar la distribución potencial de especies de pinos (e.g. Cruz-Cárdenas et al., 2016; Ramos-Dorantes et al., 2017).
Los estudios de distribución potencial de coníferas en México están restringidos a zonas específicas por la dificultad de contar con registros confiables de la especie; la calidad de los modelos generados depende del número de datos (Stockwell y Peterson, 2002), de su distribución geográfica y calidad (confiabilidad) de la información. Es probable que las áreas donde se realiza el manejo de especies de coníferas (e.g. plantaciones forestales, reforestación, migración asistida, colecta de germoplasma, etc.) no sean las que cumplan con el mejor perfil bioclimático.
En este estudio se usa una de las bases de datos más extensa y actual, e incluye toda la distribución posible de la especie en el territorio Mexicano; así, los modelos generados son una valiosa herramienta para delinear la distribución de la especie y con esto a la toma de decisiones para el manejo de las especies, así como para predecir escenarios futuros, con lo cual es más eficiente el uso de los recursos. Por lo anterior, los objetivos de este estudio fueron modelar la distribución potencial de veinte especies de pinos de importancia económica, ecológica y social en México, evaluar las variables climáticas que determinan su distribución y conocer el perfil bioclimático de cada especie. La hipótesis fue que las variables bioclimáticas y topográficas predicen adecuadamente la distribución potencial de las especies de coníferas en México.
Materiales y Métodos
Área de estudio y especies estudiadas
México se ubica en el hemisferio norte, posee una superficie de 1 964 375 km2 (INEGI, 2009), se distribuye a ambos lados del Trópico de Cáncer, con elevaciones oscilan entre 1500 y 5000 msnm; predomina el clima cálido subhúmedo ((Am(f)) con temperaturas desde 2 a 30 °C, y precipitación anual de 0 a 4 000 mm (García, 1998).
Las especies seleccionadas (símbolo - número de registros) fueron (Cuadro 1): Pinus arizonica Engelm. (Par - 541), P. ayacahuite Ehren. (Pay - 738), P. cembroides Zucc. (Pce - 1,244), P. devoniana Lindl. (Pde - 239), P. douglasiana Martínez (Pdo - 299), P. durangensis Martínez (Pdu - 1,217), P. engelmannii Carr. (Pen - 990), P. greggii Engelm. (Pgr - 33), P. hartwegii Lindl. (Pha - 64), P. herrerae Martínez (Phe - 338), P. lawsonii Roezl. (Pla - 61), P. leiophylla Schltdl. et Cham. (Ple - 906), P. lumholtzii Robins & Fern. (Plu - 812), P. maximinoi H. E. Moore (Pma - 76), P. montezumae Lamb. (Pmo - 127), P. oocarpa Schiede (Poo - 1,028), P. patula Schl. et Cham. (Ppa - 130), P. pringlei Shaw. (Ppr - 76), P. pseudostrobus Lindl. (Pps - 473) y P. teocote Schltdl. et Cham. (Pte - 830). La distribución altitudinal de muchas de estas especies varía entre 50 a 3000 msnm (Eguiluz, 1982), incluso algunas pueden alcanzar el límite superior de la vegetación arbórea de hasta 4,000 msnm (Yeaton, 1982). En los pinares la temperatura media anual fluctúa entre 6 y 28 °C y la precipitación desde 350 a más de 1000 mm año-1 (García, 1988).
Cuadro 1 Distribución de registros de especies en los estados de México.
| Estado | Par | Pay | Pce | Pde | Pdo | Pdu | Pen | Pgr | Pha | Phe | Pla | Ple | Plu | Pma | Pmo | Poo | Ppa | Ppr | Pps | Pte | Total |
| Aguascalientes | 2 | 2 | |||||||||||||||||||
| Baja California | 5 | 5 | |||||||||||||||||||
| Baja California Sur | 3 | 3 | |||||||||||||||||||
| México | 2 | 2 | 1 | 1 | 2 | 1 | 9 | ||||||||||||||
| Chiapas | 3 | 14 | 1 | 3 | 22 | 10 | 186 | 5 | 19 | 12 | 275 | ||||||||||
| Chihuahua | 343 | 526 | 657 | 50 | 711 | 538 | 7 | 99 | 390 | 317 | 1 | 57 | 2 | 119 | 3817 | ||||||
| Coahuila | 8 | 2 | 52 | 1 | 3 | 8 | 1 | 2 | 1 | 2 | 1 | 81 | |||||||||
| Colima | 2 | 1 | 3 | ||||||||||||||||||
| Durango | 173 | 143 | 270 | 16 | 97 | 482 | 368 | 167 | 326 | 352 | 106 | 15 | 451 | 2966 | |||||||
| Estado de México | 2 | 2 | 9 | 1 | 9 | 1 | 10 | 13 | 12 | 5 | 8 | 22 | 13 | 107 | |||||||
| Guanajuato | 1 | 30 | 3 | 4 | 5 | 43 | |||||||||||||||
| Guerrero | 14 | 10 | 3 | 18 | 12 | 3 | 17 | 7 | 162 | 3 | 18 | 37 | 27 | 331 | |||||||
| Hidalgo | 6 | 1 | 6 | 2 | 1 | 2 | 3 | 5 | 15 | 4 | 4 | 49 | |||||||||
| Jalisco | 3 | 7 | 50 | 58 | 3 | 2 | 1 | 10 | 4 | 28 | 70 | 9 | 21 | 147 | 2 | 13 | 12 | 440 | |||
| Michoacán | 1 | 1 | 33 | 21 | 5 | 9 | 10 | 25 | 1 | 3 | 13 | 70 | 2 | 6 | 77 | 25 | 302 | ||||
| Morelos | 1 | 2 | 3 | 5 | 1 | 1 | 3 | 1 | 17 | ||||||||||||
| Nayarit | 1 | 25 | 30 | 1 | 7 | 2 | 2 | 12 | 29 | 1 | 3 | 52 | 7 | 2 | 174 | ||||||
| Nuevo León | 5 | 3 | 53 | 2 | 2 | 2 | 3 | 1 | 1 | 30 | 13 | 115 | |||||||||
| Oaxaca | 15 | 71 | 19 | 1 | 5 | 9 | 28 | 31 | 23 | 26 | 153 | 30 | 39 | 158 | 87 | 695 | |||||
| Puebla | 12 | 3 | 1 | 1 | 9 | 3 | 10 | 13 | 4 | 31 | 5 | 34 | 13 | 139 | |||||||
| Querétaro | 9 | 1 | 7 | 1 | 2 | 1 | 21 | ||||||||||||||
| San Luis Potosí | 1 | 14 | 3 | 5 | 3 | 3 | 7 | 36 | |||||||||||||
| Sinaloa | 2 | 8 | 19 | 10 | 9 | 51 | 10 | 1 | 110 | ||||||||||||
| Sonora | 5 | 6 | 18 | 17 | 45 | 8 | 6 | 18 | 1 | 10 | 3 | 137 | |||||||||
| Tamaulipas | 17 | 2 | 1 | 2 | 12 | 17 | 51 | ||||||||||||||
| Tlaxcala | 1 | 1 | 5 | 2 | 2 | 1 | 3 | 1 | 16 | ||||||||||||
| Veracruz | 3 | 1 | 2 | 6 | 1 | 1 | 1 | 1 | 2 | 29 | 16 | 3 | 66 | ||||||||
| Zacatecas | 5 | 1 | 97 | 8 | 2 | 2 | 6 | 5 | 35 | 30 | 2 | 6 | 1 | 12 | 212 | ||||||
| Total | 541 | 738 | 1242 | 239 | 299 | 1217 | 990 | 33 | 64 | 338 | 61 | 906 | 812 | 76 | 127 | 1028 | 130 | 76 | 473 | 830 | 10222 |
Base de datos: climáticas, topográficas y biológicas
Del sitio web WorldClim se obtuvieron 19 cubiertas geográficas de variables climáticas en formato raster de 1 km2 de resolución (Cuadro 2), información derivada de temperatura y precipitación (Hijmans et al., 2005) del periodo 1950 al 2000. La altitud, exposición y pendiente se calcularon en ArcMap 10.2 a partir del Continuo de Elevaciones Mexicano 3.0 (CEM 3.0) de INEGI. Un total de 10 222 registros biológicos de presencia (coordenadas geográficas) de las especies se obtuvieron de la base de datos del Inventario Nacional Forestal y de Suelos (INFyS 2009 - 2014) proporcionados por la Comisión Nacional Forestal (CONAFOR) y antes de su análisis se verificaron para que no existieran registros repetidos y que su distribución coincidiera sobre las áreas boscosas.
Cuadro 2 Variables ambientales Bioclim y topográficas usadas para generar modelos de distribución potencial de coníferas de México.
| Variable | Descripción |
| Bio1 | Temperatura media anual (°C) |
| Bio2 | Rango de temperatura media diurna (°C) |
| Bio3 | Isotermalidad (Bio2/Bio7) (* 100) |
| Bio4 | Estacionalidad de la temperatura (desviación estándar * 100) |
| Bio5 | Temperatura máxima del mes más cálido (°C) |
| Bio6 | Temperatura mínima del mes más frío (°C) |
| Bio7 | Rango de temperatura anual (Bio5-Bio6, °C) |
| Bio8 | Temperatura promedio del trimestre más lluvioso (°C) |
| Bio9 | Temperatura promedio del trimestre más seco (°C) |
| Bio10 | Temperatura promedio del trimestre más cálido (°C) |
| Bio11 | Temperatura promedio del trimestre más frío (°C) |
| Bio12 | Precipitación anual (mm) |
| Bio13 | Precipitación del mes más lluvioso (mm) |
| Bio14 | Precipitación del mes más seco (mm) |
| Bio15 | Estacionalidad de la precipitación (Coeficiente de variación, %) |
| Bio16 | Precipitación del trimestre más lluvioso (mm) |
| Bio17 | Precipitación del trimestre más seco (mm) |
| Bio18 | Precipitación del trimestre más cálido (mm) |
| Bio19 | Precipitación del trimestre más frio (mm) |
| Expos | Exposición (grados) |
| Pend | Pendiente (grados) |
| Altitud | Altitud (msnm) |
Modelación y validación de la distribución de especies
Para generar los modelos de distribución potencial se utilizó MaxEnt versión 3.3.3k (Maximum Entropy Species Distribution Modeling) (Phillips et al., 2006), el cual solo requiere datos de la presencia de la especie. Para generar el modelo de cada especie se utilizaron todas las capas Bioclim y las topográficas, solicitando al software 500 interaciones y salida de tipo logística, este formato permite interpretarse como la estimación de probabilidad (de 0 a 1) de especies condicionada a variables ambientales (Phillips y Dudík, 2008), el 50 % del total de registros se usó para entrenamiento y el resto para prueba. Los modelos fueron evaluados acorde con pruebas curva de respuesta (análisis de omisión/comisión) y sensibilidad Curva Operada por el Receptor (ROC) - Área bajo la curva (AUC) (Elith et al., 2006; Aguirre y Duivenvoorden, 2010) y pruebas Jackknife para medir el efecto de cada variable en el modelo (Hijmans et al., 2005; Ramos-Dorantes et al., 2017). Los modelos obtenidos en MaxEnt se reclasificaron en ArcMap 10.2, para obtener superficies en los diferentes niveles de probabilidad y con ello obtener el perfil bioclimático de cada especie. Un análisis de componentes principales se realizó usando la matriz estandarizada de contribución de cada variable bioclimática y topográfica para conocer su comportamiento en función a variables Bioclim.
Resultados y Discusión
Evaluación de los modelos de predicción
La prueba estadística área bajo la curva ROC muestra que los modelos de predicción son sensibles para aquellas especies con menos de 340 registros (Figuras 1A, B, C, E, F, G, H, K, L y O), al observar que las curvas de entrenamiento y de prueba se encuentran muy dispersas del umbral de predicción, es decir, sobrestimando o subestimando las proyecciones. Para especies con más de 470 registros el ajuste fue muy bueno, puesto que en el análisis de omisión/comisión (Figuras 1D, I, J, M, N, Ñ, P, Q, R y S) la tasa de omisión es cercana a la omisión estimada (línea diagonal), tanto de la muestra de entrenamiento como la de prueba. La evaluación de la predicción de los modelos mediante la técnica ROC ha sido aceptada como un método estándar y proporciona una medida simple del desempeño del modelo (Benito de Pando y Peñas de Giles, 2007; Aguirre y Duivenvoorden, 2010).

Figura 1 Análisis del ajuste de modelos de distribución potencial para 20 especies de pino en México, mediante la técnica ROC.
El análisis de resultados muestra que tanto el valor de AUC como el valor relativo de probabilidad (VRP) disminuyen significativamente (p≤0.001) conforme aumenta el número de registros de especies (Figura 2A y 2B). Stockwell y Peterson (2002) demostraron que la modelación de cualquier especie depende de la cantidad y calidad de los datos; de esta última, la precisión geográfica de su ubicación, incluso sugieren que para que la predicción sea robusta, se utilice un mínimo de 50 registros. Aunque se ha realizado modelación de especies con menos de 6 registros (Aguirre y Duivenvoorden, 2010), ésta se ha reportado como no significativa. En nuestro estudio los registros variaron desde 33 en P. greggii hasta 1,244 en P. cembroides (Figura 1), y es claro que la modelación de la primera especie pudo no ser robusta por tener pocos registros, y el resto de las especies presenta más de 60 datos. De aquí se deriva que el ajuste del modelo aumenta con menor número de registros, pero subestimado y sobreestimado las predicciones (Figura 2A). Elith et al. (2006) demostraron que la estimación de parámetros en MaxEnt es crítica cuando el tamaño de muestra es pequeño, y viceversa.
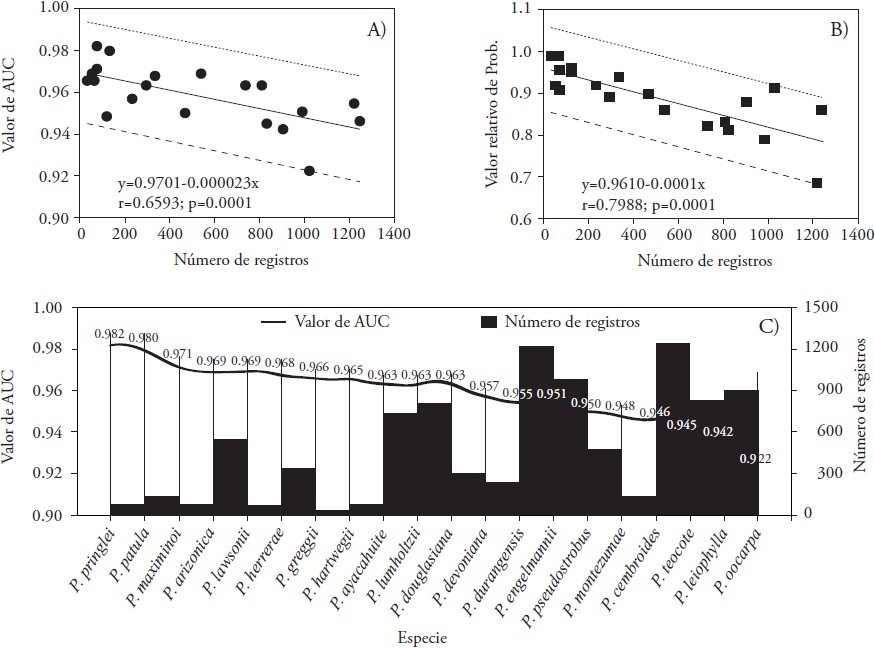
Figura 2 Relación entre valores del área bajo la curva, AUC (A), valor relativo de probabilidad (B) y número de registros. Tendencia de valores de AUC con respecto al número de registros de cada especie (C).
El promedio de los AUC de todas las especies fue 0.9589, con P. pringlei (0.982), P. patula (0.980) y P. maximinoi (0.971) como las especies con mayor ajuste, contrario a lo observado con P. oocarpa (0.922), P. leiophylla (0.942) y P. teocote (0.945) (Figura 2C), incluso estos últimos muy aceptables ya que Phillips y Dudík (2008) establecen que cuanto más próximo a 1 esté el valor de AUC mejor es el modelo. Además, este promedio supera el valor (0.75) que sugieren Elith et al. (2006) y Aguirre y Duivenvoorden (2010), quienes aseguran que valores de AUC mayores a 0.75 indican que las predicciones basadas en solo datos de presencia son lo suficientemente precisas para establecer planes de manejo.
La modelación de la distribución potencial de coníferas de nuestro estudio mediante MaxEnt se puede considerar confiable. Aguirre y Duivenvoorden, (2010) registraron un promedio de AUC de 0.9238, menor al promedio de nuestro estudio (0.9589) pero variando de 0.9994 a 0.7660. En P. pringlei, P. patula, P. maximinoi, P. teocote, P. leiophylla y P. oocarpa estudiadas por estos autores, la modelación de todas ellas se mejoró en nuestro estudio con AUC desde 0.928 hasta 0.967. Con 391 registros de P. herrerae, Ávila et al. (2014) reportaron un valor de AUC de 0.9730, similar a nuestro estudio (Figura 1). Estas variaciones en ajuste del modelo y de zonas de predicción se deben básicamente al número de registros usados y su ubicación geográfica. En 11 especies modeladas por Ramos-Dorantes et al. (2017) y consideradas en nuestro estudio, el AUC promedio fue 0.9660, levemente superior al de nuestro estudio (0.9589). En el 54.5 % de ellas, nuestra investigación mejoró la modelación (P. ayacahuite, P. greggii, P. hartwegii, P. patula, P. pseudostrobus y P. teocote). El ajuste de los modelos no siempre es bueno, y en Pinus oocarpa Schiede ex Schltdl. el valor de AUC fue solo 0.548 y 0.445 en Abies guatemalensis Rehd. (Cruz-Cárdenas et al., 2016), pero usando otras variables bioclimáticas; un valor cercano a 0.5 es realmente un valor al azar (Felicísimo et al., 2012).
Variables ambientales que modelan la distribución de las especies
Al considerar las primeras tres variables en el modelo de cada especie, las que más contribuyeron a predecir su distribución son altitud, Bio10 y Bio1, en 10, 9 y 6 especies, respectivamente (Cuadro 3). Sin embargo, una sola variable (Bio4) puede explicar el 57.4 % de la distribución potencial de P. lawsonii. El perfil bioclimático es diferente entre especies (Cuadro 3) y depende de su distribución geográfica en el país. Al respecto, Eguiluz (1982) asoció la distribución de 77 taxas del género Pinus en México, y mostró que varias de ellas dependen de temperatura máxima (11 especies donde se registran hasta 45 °C, entre ellas P. pinceana, P. pseudostrobus var. estevezii Mart., P. greggii y P. oocarpa), mínima (hasta -23 °C, donde crecen P. ayacahuite, var. brachyptera, P. lumholtzii, P. engelmannii y Pinus arizonica), precipitación (P. radiata var. binata Lem y P. greggii en 100 y 2900 mm de precipitación anual), pero también de altitud (P. muricata D. Don y P. hartwegii Lind. crecen entre 120 hasta 4000 msnm, respectivamente).
Cuadro 3 Variables ambientales con mayor porcentaje de contribución en la predicción de la distribución potencial de pinos en México y su perfil bioclimático.
| Especie | Var | % | Intervalo | Var | % | Intervalo | Var | % | Intervalo |
| Ppr | Bio4 | 57.4 | 1974 - 669 | Altitud | 27.1 | 2959 - 885 | Bio7 | 4.5 | 23.7 - 15.9 |
| Ppa | Bio10 | 36.5 | 24.2 - 11.0 | Bio7 | 13.3 | 23.8 - 15.7 | Bio5 | 12.3 | 31.8 - 19.3 |
| Pma | Bio4 | 33.1 | 2878 - 437 | Bio12 | 28.2 | 2870 - 761 | Altitud | 20.9 | 3001 - 428 |
| Par | Bio1 | 45.8 | 20.3 - 9.5 | Altitud | 14.4 | 3079 - 1051 | Bio11 | 9.9 | 15.3 - 2.5 |
| Pla | Bio4 | 46.4 | 2850 - 505 | Altitud | 28.9 | 3052 - 124 | Pendiente | 14.2 | 18.52 - 1.282 |
| Phe | Bio18 | 16.5 | 828 - 95 | Bio10 | 15.8 | 25.4 - 14.4 | Altitud | 15.0 | 2882 - 1025 |
| Pgr | Altitud | 22.2 | 2870 - 181 | Pendiente | 19.4 | 16.05 - 0.852 | Bio2 | 11.8 | 16.2 - 11.3 |
| Pha | Bio10 | 40.3 | 25.7 - 8.1 | Altitud | 22.9 | 3902 - 790 | Bio12 | 11.1 | 2363 - 692 |
| Pay | Bio1 | 33.1 | 23.6 - 9.1 | Bio10 | 27.3 | 24.9 - 11.4 | Bio17 | 5.2 | 151 - 10 |
| Plu | Bio5 | 52.6 | 33.4 - 22.1 | Bio3 | 13.0 | 68 - 51 | Bio19 | 8.2 | 274 - 32 |
| Pdo | Bio10 | 21.4 | 25.3 - 12.1 | Bio13 | 15.2 | 431 - 128 | Bio16 | 9.8 | 1218 - 311 |
| Pde | Bio7 | 22.4 | 26.8 - 15.0 | Bio10 | 15.3 | 25.9 - 12.6 | Bio8 | 12.5 | 24.8 - 11.7 |
| Pdu | Bio1 | 42.4 | 21.6 - 9.8 | Altitud | 13.1 | 3079 - 921 | Bio18 | 10.6 | 788 - 165 |
| Pen | Bio1 | 24.9 | 23.2 - 10.1 | Bio11 | 16.9 | 19.3 - 2.8 | Bio18 | 15.4 | 788 - 125 |
| Pps | Bio5 | 33.0 | 33.6 - 15.9 | Bio10 | 20.7 | 25.5 - 8.4 | Bio12 | 18.0 | 3546 - 487 |
| Pmo | Bio4 | 20.6 | 5469 - 575 | Pendiente | 18.2 | 25.5 - 0.7 | Altitud | 16.8 | 3686 -664 |
| Pce | Bio1 | 37.0 | 20.1 - 10.3 | Altitud | 27.2 | 3132 - 1068 | Bio12 | 10.3 | 1253 - 191 |
| Pte | Bio5 | 45.3 | 33.9 - 17.5 | Bio10 | 11.1 | 26.7 - 9.7 | Bio8 | 7.5 | 26.2 - 8.7 |
| Ple | Bio1 | 35.1 | 23.4 - 9.9 | Bio10 | 28.8 | 25.1 - 11.7 | Bio18 | 11.7 | 793 - 125 |
| Poo | Bio16 | 29.3 | 1670 - 207 | Bio8 | 25.4 | 26.5 - 11.3 | Bio13 | 15.9 | 635 - 72 |
Var: variables bioclimáticas y topográficas; %: porcentaje de contribución según la prueba de Jackknife. Ppr = P. pringlei; Ppa = P. patula; Pma = P. maximinoi; Par = Pinus arizonica; Pla = P. lawsonii; Phe = P. herrerae; Pgr = P. greggii; Pha = P. hartwegii; Pay = P. ayacahuite; Plu = P. lumholtzii; Pdo = P. douglasiana; Pde = P. devoniana; Pdu = P. durangensis; Pen = P. engelmannii; Pps = P. pseudostrobus; Pmo = P. montezumae; Pce = P. cembroides; Pte = P. teocote; Ple = P. leiophylla; Poo = P. oocarpa. Bio1,…, Bio19 están definidas en el Cuadro 2.
El análisis de compontes principales muestra la existencia de dos grupos claramente definidos: un grupo compacto de especies (Pinus cembroides, P. durangensis, P. arizonica, P. lumholtzii, P. leiophylla, P. herrerae, P. leiophylla y P. ayacahuite) distribuidas especialmente a Bio1, Bio11, Bio18 y Bio19; y un segundo grupo destacando P. devoniana, P. douglasiana y P. oocarpa, dependiendo de variables Bioclim derivadas de precipitación, Bio13, Bio15 y Bio16 (Figura 3A). Pinus greggii demostró una respuesta diferente las otrasespecies, y su distribución depende de precipitación del mes más seco (Bio14) y exposición. Al considerar una contribución mayor al 10 %, la altitud aparece en la estructura del modelo en más de 55 % de las especies, le siguen Bio10 (50 %) y Bio1 (35 %); las variables que no explican más de ese valor son Bio6, Bio9, Bio15, Bio17, Bio19 y exposición (Figura 3B).
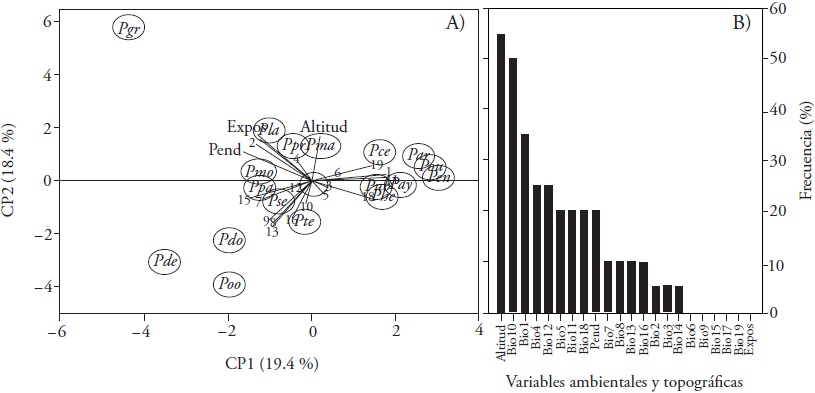
Figura 3 Análisis de componentes principales de la contribución de variables bioclimáticas y topográficas a la modelación de 20 especies de coníferas en México (A) y frecuencia en los modelos (B). La simbología se explica en Materiales y Métodos.
Similar a nuestro estudio y derivado de la prueba de Jacknife, Téllez et al. (2005) reportan que las variables que explican la distribución de P. arizonica, P. devoniana, P. durangensis y P. pseudostrobus son Bio1 y Bio12. En 12 especies de coníferas evaluadas por Ramos-Dorantes et al. (2017), Bio1 fue la variable más importante, y 11 de estas especies se incluyeron en este mismo estudio respondiendo de esta misma variable bioclimática. En Pinus herrerae Martínez, la temperatura máxima y altitud (1985 - 2227 msnm) determinan su distribución (Ávila et al., 2014), y concuerda con nuestro estudio de que la exposición no explica la distribución de especies de coníferas. Una investigación realizada por Cruz-Cárdenas et al. (2016) a escala regional en ocho especies incluidas en nuestro estudio, contrasta con lo reportado aquí: Bio6 es la variable más importante para P. douglasiana (92.3 %), P. lawsonii (76.7 %), P. pseudostrobus (92.4 %) y P. teocote (92.3 %); la concordancia existe solo con P. montezumae (48.9 %) al observar que la altitud determina su distribución.
Superficie y mapas de distribución potencial
Las tres especies con mayor superficie potencial estimada por MaxEnt acorde a una probabilidad mayor o igual que 0.70 son: P. montezumae, P. devoniana y P. pseudostrobus con 14,744.8, 14,436.1 y 11,594.8 km2 (Figuras 4 y 5O, L y Ñ),, mientras que las de menor superficie fueron P. durangensis, P. lumholtzii y P. engelmannii con 0.1, 275.9 y 819.1 km2 (Figuras 4 y 5N, J y M). Oaxaca (+) y Durango (*) son los estados que registran superficie idónea para nueve especies (Figura 4), Chihuahua para siete, Guerrero para seis, Michoacán y Puebla para cinco (Figura 4), siete estados solo registran condiciones ideales para una especie, pero diferente. El 50 % de las especies registraron un valor de probabilidad mayor a 0.9 (Figuras 5A, B, C, E, F, G, H, L, O y S), y solo P. durangensis no posee superficie dentro de este rango (Figura 5M).

Figura 4 Superficie estimada por especie en un umbral de probabilidad mayor a 0.70 y los tres estados del país con mejores condiciones de idoneidad de ocurrencia de cada especie.

La comparación de las superficies potenciales de especies de coníferas en México, entre estudios, resultaría ambigua porque los estudios pueden estar representados con diferente escala de información y análisis espacial (Ávila et al., 2014; Cruz-Cárdenas et al., 2016; Ramos-Dorantes et al., 2017); además, Téllez et al. (2005) y Aguirre y Duivenvoorden (2010) proporcionan superficies a nivel de especie y taxa sin indicar número de registros, y presentan superficies en diferentes rangos de probabilidad. De estos últimos autores, el perfil bioclimático y distribución geográfica es similar al encontrado aquí en P. arizonica, P. devoniana, P. durangensis y P. pseudostrobus.Stockwell y Peterson (2002) coinciden que el número de registros es relevante en la calidad del modelo de predicción y por ende en la estimación de superficies la cual es importante, ya que Cruz-Cárdenas et al. (2016) indican que especies como P. leiophylla y P. teocote bajo diferentes escenarios de cambio climático reducirán su superficie. La delineación de la distribución geográfica de las especies estudiadas aquí, incluye perfectamente las cinco zonas geográficas reportadas por Sánchez-González (2008), y son útiles en la evaluación de la distribución de las especies por efectos del cambio climático (Phillips y Dudík, 2008).
Conclusiones
Los modelos generados se pueden considerar confiables, ya que los valores de AUC fueron superiores a 0.90. La delineación de la distribución potencial de veinte especies de coníferas en México se ha actualizado, en niveles diferentes de probabilidad. Las variables que conforman los modelos son especialmente las derivadas de temperatura máxima, altitud y precipitación; sin embargo, cada especie tiene requerimientos bioclimáticos específicos. Oaxaca y Durango poseen las características idóneas para soportar el crecimiento para nueve especies de coníferas. El perfil bioclimático determinado para cada especie es relevante para establecer planes de manejo e incrementar su potencial productivo. La hipótesis planteada se acepta porque las variables bioclimáticas predicen adecuadamente la distribución potencial de las especies estudiadas.

