










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgrociencia
On-line version ISSN 2521-9766Print version ISSN 1405-3195
Agrociencia vol.52 n.7 Texcoco Oct./Nov. 2018
Crop Science
Phenology of ‘Méndez’ avocado in southern Jalisco, México
1 Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Campo Experimental Santiago Ixcuintla. (salazar.avocado@gmail.com).
2 Investigadores independientes.
In the southern area of the state of Jalisco, Mexico, cultivation of ‘Méndez’ avocado (Persea americana Miller) is important due to its adaptation to the semi-warm climate of the region and because of its early maturation and harvest (summer), in relation to ‘Hass’ (fall), although the lack of knowledge of its phenology makes the effective management of ‘Méndez’ orchards difficult. The objective of this study was to document the phenology of ‘Méndez’ during two production cycles (2014-15 and 2015-16), in two commercial orchards with supplementary irrigation from October to June (annual rain of 728 mm), established in soils of volcanic origin and light texture in southern Jalisco. The maximum temperatures of the soil at 30 cm from the surface were in May (24.5 °C) and the minimum from January to March (15.9 to 14.5 °C). The vegetative flushes (VF) were found in summer (August-September) and winter (February), as well as flowering in summer (September) and winter (February). Flower development (vegetative bud to anthesis) in shoots from the summer and winter VFs lasted 187 and 222 d, respectively. The fruit drop (5 to 6 cm of diameter) was similar in the two flowering seasons (55.4 to 61.8 %). The fruit maturity (≥ 22.7 % d.m.) happened in July and in September-October for fruits from the summer and winter flowering periods, respectively. There were two flushes of root production and the greatest was in summer (June to August). The maximum and minimum average environmental temperatures of the warmest (April to June) and coldest (January to March) months were 31 and 9 °C, respectively. The results from this study are basic to compare the future changes in the phenology of ‘Méndez’ avocado.
Keywords: vegetative flushes; flower development; roots; ecophysiology
En el sur del estado de Jalisco, México, el cultivo del aguacate (Persea americana Miller) ‘Méndez’ es importante por su adaptación al clima semicálido de la región y por su maduración y cosecha anual precoz (verano), respecto a ‘Hass’ (otoño), pero el desconocimiento de su fenología dificulta el manejo efectivo de los huertos de ‘Méndez’. El objetivo de esta investigación fue documentar la fenología de ‘Méndez’ durante dos ciclos de producción (2014-15 y 2015-16), en dos huertos comerciales con riego suplementario de octubre a junio (lluvia anual de 728 mm), establecidos en suelos de origen volcánico y textura ligera en el sur del estado de Jalisco. Las temperaturas máximas del suelo a 30 cm de la superficie fueron en mayo (24.5 °C) y las mínimas de enero a marzo (15.9 a 14.5°C). Los flujos vegetativos (FV) se registraron en verano (agosto-septiembre) e invierno (febrero) así como floraciones en verano (septiembre) e invierno (febrero). El desarrollo floral (yema vegetativa a antesis) en brotes del FV de verano e invierno duró 187 y 222 d, respectivamente. La caída de fruto (5 a 6 cm de diámetro) fue similar en las dos épocas de floración (55.4 a 61.8 %). La madurez del fruto (≥ 22.7 % m.s.) ocurrió en julio y en septiembre-octubre para frutos de las floraciones de verano e invierno, respectivamente. Hubo dos flujos de producción de raíces y el mayor fue en verano (junio a agosto). Las temperaturas ambientales máximas y mínimas promedio de los meses más calientes (abril a junio) y fríos (enero a marzo) fueron de 31 y 9 °C, respectivamente.). Los resultados de este estudio son básicos para comparar los cambios futuros en la fenología de ‘Méndez’.
Palabras clave: flujos vegetativos; desarrollo floral; frutos; raíces; ecofisiología
Introduction
Throughout the year, fruit trees present diverse development or phenological phases, such as vegetative or root growth flushes, and of flower and fruit development (Wolstenholme and Whiley, 1989). The rhythms of annual growth vary with the type of climate and the cultivation conditions, so phenological diagrams were elaborated for ‘Hass’ avocado (Persea americana Miller) with different degree of complexity in production regions like Australia (Whiley et al., 1988), South Africa (Whiley and Wolstenholme, 1990), Chile (Gardiazabal-Irazábal and Rosenberg, 1991), USA (Arpaia et al., 1994-95), and New Zealand (Thorp et al., 1995). In Mexico, ‘Hass’ phenology is described for the production regions of Nayarit (Cossio-Vargas et al., 2008) and Michoacán (Rocha-Arroyo et al., 2011a). The avocado ‘Méndez No. 1’ (Plant Patent 11,173 USA 2002), known in Mexico as ‘Méndez’ is economically important in the southern area of the state of Jalisco, where there are more than 6000 ha planted. Its relevance is because an important proportion of its annual harvest matures earlier (summer) than ‘Hass’ (fall-winter), and this harvest reaches the highest prices of the year.
Due to lack of knowledge of the horticultural aspects of ‘Méndez’, orchard management is done similarly to ‘Hass’. However, given the growing importance of ‘Méndez’ it is necessary to develop appropriate production technology for this avocado cultivar. Therefore, the objective of this study was to document the phenological behavior of ‘Méndez’ avocado in the southern region of the state of Jalisco.
Materials and Methods
The research was performed during the production cycles 2014-15 and 2015-16 in two commercial orchards of ‘Méndez’ avocado from the Agro González, S.P.R. de R.L. company, with fertirrigation, haplic Feozem soil and Semi-warm sub-humid climate [AC(w)] (García-Amaro, 1998) from the south of the state of Jalisco. The Colorín 1 orchard is in Atequizayan, municipality of Zapotlán el Grande, and the Ocote Cuate 2 orchard is in Zapotiltic, municipality of Zapotiltic. For these orchards, the altitude is 1556 m and 1428 m, the distance between trees 7×3.5 m and 5×5 m, and the age at the beginning of the study 4 and 6 years, respectively.
Characteristics of the soil
Five trees were selected randomly in each orchard, and a soil sample was taken made up of four sub-samples from the drip zone of the tree, at 0-30 cm of depth, which is the zone with highest amounts of young roots (Salazar-García and Cortés-Flores, 1986). The analyses were performed in a laboratory accredited by the North American Proficiency Testing (NAPT) program (a program by the Soil Science Society of America). The variables were texture, pH (1:2 water) (McLean, 1982), organic matter (OM) by the Walkley and Black method (Nelson and Sommers, 1982), inorganic-N (Dahnke, 1990), P-Bray (Bray and Kurtz, 1945), K, Ca, Mg and Na with ammonium acetate (Doll and Lucas, 1973), Fe, Zn, Cu, Mn by DTPA method (Lindsay and Norvell, 1978), and B was determined by the hot water and Azomethine-H method (Bingham, 1982). The variables P and B were determined in a Genesys 20 spectrophotometer (Thermo Scientific, Madison, USA); an atomic absorption spectrophotometer (Thermo Series S, Shangai, China) was used for the other nutrients.
Phenological variables
Twenty shoots per tree were tagged in 10 trees per orchard, at the beginning of the vegetative growth flushes of winter and summer 2014 and 2015. From each tree and shoot marked, an apical bud or flower shoot was collected at biweekly or weekly intervals as anthesis was approached. The buds and flower shoots were fixed in FAA (formaldehyde:acetic acid:ethanol, 5:5:90, v:v:v) and then they were introduced into a vacuum bell (Nalgene 8040317, Nalgen Company) at 30 KPa for 5 h. Later they were classified with a stereoscopic microscope (Zeiss Stereomikroskop Mod. Stemi 2000-C, Carl Zeiss, Göttingen, Germany), with the visual scale suggested by Salazar-García et al. (1998) that covers from S-1 (vegetative bud) to S-11 (anthesis).
In each orchard, another group of 10 trees was selected, with a history of annual production of at least 50 kg per tree, and they were used in all the assessments except those of flower development. In February and August 2014 and 2015, 30 shoots were labeled on each tree at the beginning of each vegetative flush (winter and summer, respectively). The types of growth produced (floral, vegetative or inactive shoots) by the tagged shoots were quantified at the end of each flowering period, in October and February 2014, and in November 2015 and March 2016.
The root samples were taken each month from February 2014 to February 2016, alternating odd and even trees, as well as north and south orientation of the trees. The young roots, of light brown color, in a soil volume of 40 ( 40 ( 40 cm in the drip zone of the tree, were extracted, washed and dehydrated in a forced air oven (Binder Mod. ED 240, Binder Corp., Tutllingen, Germany) at 70 °C for 72 h, to obtain their dry weight.
The fruit drop was quantified each year for fruits from the summer and winter flowering periods. In anthesis, five floral shoots were marked in each cardinal point from the medium part of the canopy from every 10 trees selected. For fruit from the summer flowering, the number of fruit were counted per flower shoot, two (November) and six (March) months after anthesis. In fruit from the winter flowering, this was done two (April) and five (July) months after anthesis.
Five fruit were labeled in each tree marked, from each flowering period, when they were 2 cm long. Each month the length and diameter were measured with a digital Vernier scale (Mitutoyo, Mod. CD-6’’ CSX, Mitutoyo Corp., Kawasaki, Japan) from October (fruit from the summer flowering) and April (fruit from the winter flowering).
Climate variables
The data of rainfall were obtained from the National Meteorological Service (Servicio Meteorológico Nacional, SMN, 2016). The environmental and soil (at 30 cm of depth) temperatures were recorded in each orchard with battery-operated automatized thermometers (HOBO H8, Onset Computer, Witzprod, Englewood Cliffs, NJ, USA).
Experimental design an statistical analysis
The experimental design was random blocks with a 2 ( 2 factorial arrangement: orchards (Colorín 1 and Ocote Cuate 2), and vegetative flushes (summer and winter), with 10 replicates (trees); and the experimental unit was 30 shoots per tree. The ANOVA was performed with repeated measurements for each orchard and vegetative flush. Prior to their analysis, the values expressed as percentage were transformed by the arcsine
Results and Discussion
Soil fertility
The characteristics of soil fertility were similar between orchards. The OM, Ca and Mg ranged from moderately low to low and in the Colorín 1 orchard the low levels of Mn, B and Zn were evident (Table 1). The low content of OM in addition to the insufficient levels of Mn, B and Zn are characteristic of soils of volcanic origin (Salazar-García et al., 2016), suggesting the need to incorporate OM and nutrients in moderately low to low concentrations to improve the soil fertility. The pH was neutral in both orchards (Table 1). In other avocado production regions, the soils present similar or more acidic pH (Aguilera-Montañez and Salazar-García, 1991; Salazar-García and Lazcano-Ferrat, 1999). However, avocado can grow in pH of 4.8 or higher than 6.5 (Salazar-García, 2002).
Table 1 Soil characteristics of the orchards at 0-30 cm of depth at the beginning of the study (February 2014).
| Características | Huerto Colorín 1 | Huerto Ocote Cuate 2 |
| pH (1:2 H2O) | 7.1 (Neutro) | 6.6 (Neutro) |
| Materia orgánica (%) | 0.82 (B) | 1.7 (MoB) |
| Conductividad eléctrica (dS/m) | 0.26 | 0.33 |
| mg kg-1 | ||
| N-NO3 | 5.6 (MB) | 6.4 (MB) |
| P-Bray | 22.8 (M) | 66.4 (MA) |
| K | 180 MoB) | 491 (MoA) |
| Ca | 1273 (MoB) | 859 (MoB) |
| Mg | 96.3 (B) | 183 (MoB) |
| Na | 74 (M) | 49.1 (B) |
| Fe | 17.6 (MoA) | 79.4 (MoA) |
| Cu | 2.3 (A) | 4.0 (A) |
| Mn | 1.6 (MB) | 3.9 (B) |
| Zn | 0.54 (B) | 1.2 (MoA) |
| B | 0.22 (MB) | 0.32 (MB) |
Laboratory interpretation: A: high; MoA: moderately high; MA: very high; M: medium; B: low; MoB: moderately low; MB: very low; N: normal.
Type of growth
The production of flower shoots had different behavior between orchards. In Colorín 1, 33.1 % was found and in Ocote Cuate 2, 19.1 % (Tables 2 and 3). The decrease in environmental temperature is the factor that promotes floral development in avocado (Salazar-García et al., 1999, 2006a, 2013). The difference in production of floral shoots between orchards is explained by the lower temperatures in Colorín 1 (annual average 20.5 °C) compared to Ocote Cuate 2 (22.5 °C) (Figures 1a and 1b), which favored a greater exposure of the El Colorín 1 orchard to cooler temperatures. The vegetative flush of origin (winter or summer) did not affect the type of growth produced (vegetative, floral or inactive) (Tables 2 and 3).
Table 2 Variance analysis and orchard interactions x flush of the type of growth produced by shoots from the winter and summer vegetative flushes of ‘Méndez’ avocado. Data from two years from the orchards Colorín 1 and Ocote Cuate 2.
| Tipo de crecimiento | Fuente de variación† | GL | SC | CM | F | P > F |
| Brotes vegetativos | Huerto | 1 | 0.01413000 | 0.01413000 | 0.00 | 0.9525 |
| Flujo | 1 | 3.27166727 | 3.27166727 | 0.83 | 0.3658 | |
| Huerto*Flujo | 1 | 0.42702000 | 0.42702000 | 0.11 | 0.7434 | |
| Brotes florales | Huerto | 1 | 47.17805953 | 47.17805953 | 6.58 | 0.0119 |
| Flujo | 1 | 3.01030617 | 3.01030617 | 0.42 | 0.5185 | |
| Huerto*Flujo | 1 | 14.57155268 | 14.57155268 | 2.03 | 0.1572 | |
| Inactivos | Huerto | 1 | 76.71389754 | 76.71389754 | 14.12 | 0.0003 |
| Flujo | 1 | 3.53008144 | 3.53008144 | 0.65 | 0.4223 | |
| Huerto*Flujo | 1 | 23.07924822 | 23.07924822 | 4.25 | 0.0421 |
†Variance analysis performed with transformed data through the arcsine
Table 3 Effect of the orchard and vegetative flush of origin on the type of growth produced in ‘Méndez’ avocado. Assessments performed in: October 2014 and November 2015 (winter shoots) and February 2015 and March 2016 (summer shoots).
| Fuente de variación | N† | Tipo de crecimiento del total de brotes marcados (%) | ||
| Vegetativo | Floral | Inactivo | ||
| Huerto Colorín 1 | 774 | 53.8 | 33.1 a¶ | 13.0 b |
| Huerto Ocote Cuate 2 | 775 | 53.8 | 19.3 b | 26.7 a |
| Pr > F | 0.9525 | 0.0119 | 0.0003 | |
| Brotes de invierno | 917 | 52.2 | 26.3 | 21.1 |
| Brotes de verano | 634 | 55.8 | 26.3 | 17.8 |
| Pr > F | 0.3658 | 0.5185 | 0.4223 | |
†Number of shoots evaluated.
¶ Means with different letter in a column for each orchard or type of shoot are statistically different (Tukey; p≤0.05).

Vegetative growth
The phenological record began with the budbreak (February 2014) of shoots from the winter vegetative flush. Out of 100 % of winter shoots marked, 25 % flowered in the summer (September 2014) and 6 % flowered the following winter (February 2015). Of the winter shoots, 55 % produced summer shoots in September 2014, and of these, 27 % flowered in winter (February 2015). The winter and summer shoots produced an important number of vegetative shoots (between 50 and 60 %) (Figure 2). In contrast with ‘Méndez’, in ‘Hass’ the winter flush does not produce flowering in the summer; however, it can happen if the environmental conditions are favorable (Rocha-Arroyo et al., 2011a).

In ‘Hass’ the winter vegetative flush (known as spring flush in other regions) is the most important for the winter-spring flowering of the following year, and it is documented in Mexico (Cossio-Vargas et al., 2008; Rocha-Arroyo et al., 2011b) and other countries (Mena-Volker, 2004; Robinson et al., 2002; Salazar-García and Lovatt, 1998; Thorp et al., 1995; Whiley et al., 1990).
Flower development
The duration of flower development (from vegetative shoot to anthesis) showed slight variation between years. For 2014, 218 and 180 d went by for the winter and summer shoots, respectively. The 1st or summer flowering presented anthesis in September, and the 2nd or winter flowering, in February (Figure 3). In 2015, the winter shoots completed flower development in 225 d, producing the 1st flowering between the first week of September and the first of October. In the case of summer shoots, the flower development required 193 d, with anthesis on the first week of March 2016 (2nd flowering).
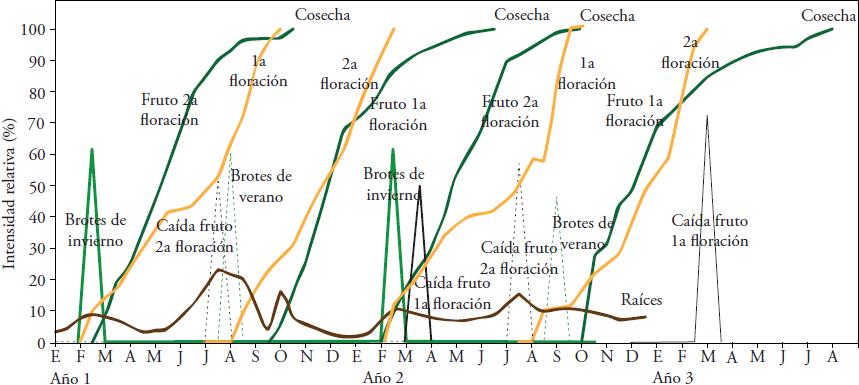
Figure 3 Main phenological stages of ‘Méndez’ avocado in the southern region of the state of Jalisco. 1st flowering: summer flowering; 2nd flowering: winter flowering.
In the two years of study, the summer shoots required, in average, 35 d less than the winter ones to complete the flower development. This difference was lower than the 120 and 180 d mentioned for both flushes of ‘Hass’ in Nayarit and Michoacán, respectively (Cossio-Vargas et al., 2008; Rocha-Arroyo et al., 2011a). The lesser time required for the summer ‘Méndez’ shoots to complete their flower development could be because when they emerged (August-September) the environmental temperature had already begun its descent (21 °C in average), stimulating floral development. The effect of the temperature decrease in developing ‘Hass’ shoots was mentioned by Rocha-Arroyo et al. (2010) and Salazar-García et al. (2013).
Fruit drop
The fruit dropped in the so-called “second drop”, “physiological drop” or “June drop” (northern hemisphere) presented diameters between 5 and 6 cm. For fruits from the first flowering period (summer), the drop was 61.8 % and it happened in March. The fruit drop from the second flowering period (winter) was in July (55.4 %). The magnitude of the physiological drop recorded in our study is similar to the one described for ‘Hass’ in other production regions: 45 % in Nayarit, Mexico (Cossio-Vargas et al., 2008) and 54 % in South Africa (Toerien, 1979). The age of the dropped fruit from the summer and winter flowering periods was 22 and 26 weeks after anthesis, respectively. The fruit drop from the second flowering was shortly before the summer vegetative flush and for those from the first flowering, it was after the winter vegetative flush (Figure 3).
Cossio-Vargas et al. (2008) and Lovatt (1990) mention the coincidence, and Wolstenholme and Whiley (1989) the lack of coincidence between the fruit drop and the presence of vegetative growth, but there is no conclusive information about the causes that originate the second fruit drop (Dixon et al., 2006). A hypothesis mentions the hormonal imbalance as responsible for this fruit drop, and highlights abscisic acid (ABA) as primary cause (Adato and Gazit, 1977). In order to decrease the intensity of this drop in ‘Hass’ avocado, leaf spraying with 2,4-D or aminoethoxyvinylglycine were evaluated because two months before the beginning of the fruit fall, and in the second year of study, it decreased 18 and 17.7 %, respectively; in addition, it increased the production of fruit of large calibers (170 to ≥ 266 g fruit-1) in 32 and 23.6 %, respectively, compared to the control (Salazar-García et al., 2006b). Garner and Lovatt (2016) confirmed that ABA is involved in the second fruit drop in ‘Hass’ avocado in California.
Fruit growth
The ‘Méndez’ fruit that result from the summer and winter flowering periods required 10 and eight months, respectively, to reach physiological maturity (22.7 % d.m.) (Figure 3). For fruit from the winter flowering, there are similar results for ‘Hass’ (21.5 % d.m.) in Nayarit and Michoacán (Cossio-Vargas et al., 2008; Rocha-Arroyo et al., 2011a). The fruit from the summer flowering reached physiological maturity in July, with an average diameter of 6.5 cm; and those from the winter flowering between September and October, with an average diameter of 6.4 cm. The fruits derived from the summer flowering required two more months to reach physiological maturity, which can be attributed to the fact that when they began their growth (September-October), the air (21.8 °C) and soil (20.8 °C) temperatures had begun their descent, which decreased the rhythm of tree metabolism.
Root production
Root production did not stop during the year and there were two periods of higher intensity (Figure 3). The most important flush was in the rainy months of the summer (June-August), prior to the summer vegetative flush. The second flush of root growth was less intense, from February to March, and coincided with the winter flowering and its corresponding vegetative flush. The highest production of roots in the summer coincided with what is described for ‘Hass’ cultivated without irrigation in Nayarit (Cossio-Vargas et al., 2008), and with and without irrigation for the same cultivar in Michoacán (Bárcenas-Ortega et al., 2007). In Israel, Kalmar and Lahav (1976) found in ‘Ettinger’ and ‘Fuerte’ a positive association between the amount of water applied and the root production, but in Michoacán, Bárcenas-Ortega et al. (2007) documented the opposite in ‘Hass’.
According to Bernstein et al. (2004), in avocado, root growth can be more restricted by the salinity than by the presence of vegetative shoots. In our study, in both orchards the CE of the soil varied from 0.26 to 0.33 dS m-1, which is why it was not a limiting factor for the development of roots and did not affect the fruit drop. In addition, the lowest production of roots happened when the fruit was at its maximum growth (Figure 3).
Rainfall and air and soil temperatures
In both orchards, the annual rainfall was 728 mm, concentrated between June and September (Figure 1). In the Colorín 1 orchard, the maximum and minimum average temperatures of the warmest (April and May) and coldest (January and February) months were 32, 31 and 6.4 and 6.0 °C, respectively (Figure 1A). In the Ocote Cuate 2 orchard the temperatures of the warmest (May and June) and coldest (March) months were 30.8 and 30.6 and 15.0, respectively (Figure 1C).
The highest soil temperature (23.8 °C) was in May and the lowest (14.5 °C) in March in Colorín 1 (Figure 1B). For Ocote Cuate 2 the maximum soil temperature was in May (25.2 °C) and the minimum in January (15.9 °C) and February (16.0 °C) (Figure 1D). In both orchards, the maximum soil temperatures remained between 21 and 27 °C, mentioned by Yusof et al. (1969) as adequate for the growth of avocado roots. Likewise, the minimum temperature was higher than 13 °C, pointed out by Whiley et al. (1990) as the lower limit from which avocado root growth stops.
Conclusions
The ‘Méndez’ avocado presented two vegetative growth flushes: winter and summer. The summer shoots produced winter flowering, and the winter shoots summer flowering. Floral development was completed in 186 and 221 d for summer and winter shoots, respectively. The maximum root production was in rainy season and the lowest during the maximum growth of the fruit. The fruit drop was slightly higher for the 1st flowering (summer). Fruits from the summer flowering period required two more months than those from winter to achieve physiological maturity. The permanent presence of unoccupied vegetative shoots (without fruit) together with the decrease in temperature during spring are key for the summer flowering whose fruit is of high commercial value.
Literatura Citada
Adato, I., and S. Gazit. 1977. Role of ethylene in avocado fruit development and ripening: I. Fruit drop. J. Exp. Bot. 28:636-643. [ Links ]
Aguilera-Montañez, J. L., y S. Salazar-García. 1991. The avocado industry in Michoacán México. South African Avocado Growers' Assn. Yrbk. 14: 94-97. [ Links ]
Arpaia, M. L., G. W. Witney, P. W. Robinson, and M. V. Mickelbart. 1994-95. ‘Hass’ avocado phenology in California: preliminary results. Subtrop. Fruit News 3: 1-2. [ Links ]
Bárcenas-Ortega, A.E., J. Pantoja-Ambríz, S. Aguirre-Paleo, A. T. Chávez-Bárcenas, S. Salazar-García, y J. L. González-Durán. 2007. Influencia del riego sobre la dinámica de crecimiento de raíces en árboles de aguacate ‘Hass’. Proc. Interamer. Soc. Trop. Hort. 51: 23-29. [ Links ]
Bernstein, N., A. Meiri, and M. Zilberstaine. 2004. Root growth of avocado is more sensitive to salinity than shoot growth. J. Amer. Soc. Hort. Sci. 129(2):188-192. [ Links ]
Bingham, F. T. 1982. Boron. In: Page, A. L., R. H. Miller, and D. R. Keeney (eds). Methods of Soil Analysis, part 2. Second edition. Amer. Soc. Agron. and Soil Sci. Soc. Amer. Madison, WI, USA. pp: 431-446. [ Links ]
Bray, R. H., and L. T. Kurtz. 1945. Determination of total, organic and available phosphorus in soil. Soil Sci. 59: 39-45. [ Links ]
Cossio-Vargas, L. E., S. Salazar-García, I. J. L. González-Durán, y R. Medina-Torres. 2008. Fenología del aguacate ‘Hass’ en el clima Semicálido de Nayarit. Rev. Chapingo Ser. Hort. 14(3): 319-324. [ Links ]
Dahnke, W. C. 1990. Testing soils for available nitrogen. In: Westerman, R. L. (ed.). Soil Testing and Plant Analysis. Amer. Soc. Agron. and Soil Sci. Soc. Amer. Madison, WI, USA. pp: 120-140. [ Links ]
Dixon, J., C. B. Lamond, D. B. Smith, and T.A. Elmlsy. 2006. Patterns of fruit growth and fruit drop of ‘Hass’ avocado trees in the Western Bay of Plenty, New Zealand. New Zealand Avocado Growers’ Assn. Ann. Res. Report 6:47-54. [ Links ]
Doll, E. C., and R. E. Lucas. 1973. Testing soil for potassium, calcium and magnesium. In: Walsh, L. M., and J. D. Beaton (eds). Soil testing and Plant Analysis. Soil Sci. Soc. Amer. Madison, WI, USA. pp: 133-152. [ Links ]
García-Amaro, E. 1998. Comisión Nacional para el estudio de la Biodiversidad (CONABIO). ‘Cartas de clima de la República Mexicana’ (Modificaciones al sistema de clasificación de Köppen). Escala 1:1000 000 México. Shapefile. http://www.igeograf.unam.mx/sigg/utilidades/docs/pdfs/publicaciones/geo_siglo21/serie_lib/modific_al_sis.pdf (Consulta: enero 2016). [ Links ]
Gardiazabal-Irazábal, F., y G. Rosenberg. 1991. Cultivo del Palto. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 201 p. [ Links ]
Garner, L. C., and C. J. Lovatt. 2016. Physiological factors affecting flower and fruit abscission of ‘Hass’ avocado. Scientia Hort. 199: 32-40. [ Links ]
Kalmar, D., and E. Lahav. 1976. Water requirements of avocado in the Western Galilee. a) The effect of different irrigation treatments on water consumptions, salt content in the soil and root distribution. Alon Hanotea. 30:629-643. [ Links ]
Lindsay, W. L., and W. A. Norvell. 1978. Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Amer. J. 42: 421-428. [ Links ]
Lovatt, C. J. 1990. Factors affecting fruit set/early fruit drop in avocado. California Avocado Soc. Yrbk 74: 193-199. [ Links ]
McLean, E. O. 1982. Soil pH and lime requirement. In: Page, A. L., R. H. Miller, and D. R. Keeney (eds.). Methods of Soil Analysis, Part 2. Second edition. Amer. Soc. Agron. and Soil Sci. Soc. Amer. Madison, WI, USA. pp: 199-223. [ Links ]
Mena-Volker, F. 2004. Fenología del palto, su uso como base del manejo productivo. 2º Seminario Internacional de Paltos. 29 Sep.-1 Oct. Quillota, Chile. Sociedad Gardiazábal y Magdahl Ltda. http://www.avocadosource.com . (Consulta Abril 2013). [ Links ]
Nelson, D. W., and L. E. Sommers. 1982. Total carbon, organic carbon and organic matter. In: Page, A. L., R. H. Miller, and D. R. Keeney (eds). Methods of Soil Analysis, Part 2. Second edition. Amer. Soc. Agron. and Soil Sci. Soc. Amer. Madison, WI, USA. pp: 539-594. [ Links ]
Robinson, P. W., M. V. Mickelbart, X. Liu, C. Adams, G. Witney, and M. L. Arpaia. 2002. Development of a phenological model of avocado tree growth in California. In: Drew, R. (ed.). Proc. Intl. Symp. Trop. and Subtropical Fruits. Acta Hort. 575: 859-864. [ Links ]
Rocha-Arroyo, J. L., S. Salazar-García, A. E. Bárcenas-Ortega, I. J. L. González-Durán, y L. E. Cossio-Vargas. 2011a. Fenología del aguacate ‘Hass’ en Michoacán. Rev. Mex. Cienc. Agríc. 2: 303-316. [ Links ]
Rocha-Arroyo, J. L., S. Salazar-García, A. E. Bárcenas-Ortega, I. J. L. González-Durán, y R. Medina-Torres. 2011b. Crecimientos vegetativo y reproductivo del aguacate ‘Hass’ en diversos climas de Michoacán, México. In: Proc. VII World Avocado Congr. Sept. 5-9, 2011. Cairns, Australia. pp: 463-445. [ Links ]
Rocha-Arroyo, J. L., S. Salazar-García, y A. E. Bárcenas-Ortega. 2010. Determinación irreversible a la floración del aguacate ‘Hass’ en Michoacán. Rev. Mex. Cienc. Agríc. 1: 469-478. [ Links ]
Salazar García, S. and J. I. Cortés Flores, 1986. Root distribution of mature avocado trees growing in soils of different texture. Calif. Avocado Soc. Yrbk. 70: 165-174. [ Links ]
Salazar-García, S. 2002. Nutrición del Aguacate, Principios y Aplicaciones. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias e Instituto de la Potasa y el Fósforo. Querétaro, México. 165 p. [ Links ]
Salazar-García, S., and C. J. Lovatt. 1998. GA3 application alters flowering phenology of the ‘Hass’ avocado. J. Amer. Soc. Hort. Sci. 123: 791-797. [ Links ]
Salazar-García, S., and I. Lazcano-Ferrat. 1999. Diagnóstico nutrimental del aguacate ‘Hass’ bajo condiciones de temporal. Rev. Chapingo Ser. Hort. 5: 173-184. [ Links ]
Salazar-García, S., E. M. Lord, and C.J. Lovatt. 1998. Inflorescence and flower development of the ‘Hass’ avocado (Persea americana Mill.) during “on” and “off” crop years. J. Amer. Soc. Hort. Sci. 123: 537-544. [ Links ]
Salazar-García, S., E.M. Lord, and C.J. Lovatt. 1999. Inflorescence development of the ‘Hass’ avocado: commitment to flowering. J. Amer. Soc. Hort. Sci. 124: 478-482. [ Links ]
Salazar-García, S., L. C. Garner, and C.J. Lovatt. 2013. Reproductive biology. In: Schaffer, B., B. N. Wolstenholme, and A.W. Whiley (eds). The Avocado, Botany, Production and Uses. 2nd Edition CABI, Oxfordshire, UK. pp: 118-167. [ Links ]
Salazar-García, S., L. E. Cossio-Vargas, C. J. Lovatt, I. J. L. González-Durán, and M. H. Pérez Barraza. 2006a. Crop load affects vegetative growth flushes and shoot age influences irreversible commitment to flowering of ‘Hass’ avocado. HortScience 41: 1541-1546. [ Links ]
Salazar-García, S., L. E. Cossio-Vargas, I. J. L. González Durán., and C. J. Lovatt. 2006b. Effect of foliar-applied plant bioregulators on “June fruit drop”, yield and fruit size of ‘Hass’ avocado. Acta Hort. 727: 197-202. [ Links ]
Salazar-García, S., M. E. Ibarra-Estrada, y R. Medina-Torres. 2016. Tejidos alternativos al foliar para evaluar la respuesta a la fertilización con Zn o B en aguacate ‘Hass’. Rev. Fitotec. Mex. 39: 247-252. [ Links ]
SMN. Sistema Meteorológico Nacional. 2016. http://smn.cna.gob.mx/tools/RESOURCES/Normales8110/NORMAL14030.TXT/ (Consulta: agosto 2016). [ Links ]
Statistical Analysis System (SAS Institute). (2011). User’s Guide: Mathematical Programming. Version 9.3. Cary, N.C. USA. [ Links ]
Steel, R. G. D., and J. H. Torrie. 1984. Principles and Procedures of Statistics: A Biometrical Aproach. 2th. Ed. McGraw Hill Publishers. pp: 234-235. [ Links ]
Thorp, T. G., P. Anderson, and M. Camilleri. 1995. Avocado tree growth cycles - a quantitative model. In: III Proc. World Avocado Congr. Oct. 22-27, 1995. Tel Aviv, Israel. pp: 76-79. [ Links ]
Toerien, J. C. 1979. Seasonal incidence of ring-neck on avocados and possible causes. South African Avocado Growers’ Assn. 3:49-51. [ Links ]
Whiley, A. W., and B. N. Wolstenholme. 1990. Carbohydrate management in avocado trees for increased production. South African Avocado Growers’ Assn. Yrbk. 13: 25-27. [ Links ]
Whiley, A. W., B. N. Wolstenholme, J. B. Saranah, and P. A. Anderson. 1990. Effect of root temperature on growth of two avocado rootstocks cultivars. Acta Hort. 275: 153-160. [ Links ]
Whiley, A. W., J. B. Saranah, B. W. Cull, and K. G. Pegg. 1988. Manage avocado tree growth cycles for productivity gains. Qld. Agric. J. 114: 29-36. [ Links ]
Wolstenholme, B. N., and A. W. Whiley. 1989. Carbohydrate and phenological cycling as management tools for avocado orchards. South African Avocado Growers’ Assn. Yrbk. 12: 33-37. [ Links ]
Yusof, I. M., D. W. Buchanan, and J. F. Gerber. 1969. The response of avocado and mango to soil temperature. J. Amer. Soc. Hort. Sci. 94: 619-621. [ Links ]
Received: April 2017; Accepted: August 2017
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
