











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En México, el arroz se cultiva en un área aproximada de 42,310 ha con un rendimiento de grano promedio nacional de 6.3 Mg ha-1 (SAGARPA, 2017). El 75 % de esta superficie se cultiva en condiciones de riego o inundación permanente durante el ciclo, con un rendimiento medio de 6.2 Mg ha-1 y el 25 % restante se cultiva bajo condiciones de secano con rendimientos promedio de 4.4 Mg ha-1 (SAGARPA, 2017). En el país se distinguen tres zonas agroclimáticas para el cultivo del arroz: trópico húmedo, trópico subhúmedo y trópico seco. Cada zona tiene diferentes problemáticas agronómicas y de producción, entre las que destacan la restricción en el abastecimiento de agua para riego; irregularidad pluvial con frecuentes periodos de sequía intermitente intra-estival o terminal o ambos; inundaciones provocadas por huracanes; estrés por altas temperaturas; factores bióticos como malezas, plagas y enfermedades; deficiencias o toxicidad nutricional y altos costos de producción (García et al., 2011; Barrios-Gómez et al., 2016). El estado de Morelos es una de las zonas de trópico seco que destaca por altos rendimientos de grano bajo riego, con un promedio de 10.2 Mg ha-1 (SAGARPA, 2017) lo que se debe, presumiblemente, a la utilización de variedades con mayor densidad del grano y mayor número de panículas por unidad de área en siembra directa en las zonas ‘alta’ (1,000 a 1,400 msnm) y ‘baja’ (800 a 1000 msnm) (Salcedo y Barrios, 2013).
El arroz requiere un alto volumen de agua a lo largo de su ciclo biológico y es el único cereal comercial que se cultiva en suelos saturados de agua (en algunos sistemas, las plantas se mantienen sumergidas) durante una parte o la totalidad del ciclo biológico; por esta razón, el suministro adecuado de agua es el factor más importante en la producción (Chatel et al., 2010). Pero hay áreas de cultivo donde la producción del grano depende, en gran medida, de la lluvia durante el ciclo; en estas condiciones, las plantas pueden experimentar periodos de estrés hídrico, además del estrés causado por otros factores bióticos y abióticos que reducen considerablemente el rendimiento y la calidad del grano (Miranda-Domínguez et al., 2016). El déficit hídrico, por sí mismo, es el factor abiótico con mayor impacto en el rendimiento del arroz de secano al afectar características morfológicas (número de panículas y de espiguillas por panícula; altura de planta; biomasa aérea; número de macollos, raíces y hojas), fisiológicas (fotosíntesis, transpiración, conductancia estomática, eficiencia en el uso del agua, contenido relativo de agua, contenido de clorofila, actividad del fotosistema II, estabilidad de la membrana, discriminación isotópica de carbono y contenido de ácido abscísico), bioquímicas (acumulación de prolina, azúcares, poliaminas y antioxidantes) y genómicas (expresión de genes que codifican para factores de transcripción y proteínas de defensa) de las plantas (Shah, et al., 2011; Pandey y Shukla, 2015; Khondia et al., 2015).
La raíz percibe el déficit hídrico edáfico, su función clave es la absorción de agua y nutrientes del suelo, y es un componente significativo de la adaptación de la planta en ambientes con limitaciones de humedad. La obtención de nuevas variedades con caracteres mejorados del crecimiento de la raíz para la adquisición de agua en suelos en proceso de secamiento, podría tener un impacto significativo en la seguridad alimentaria nacional y global (Lynch et al., 2014). En estas condiciones, el rendimiento de grano podría mejorarse identificando en la planta los factores genéticos y fisiológicos que determinan mayor acumulación de biomasa y asignación de materia seca al grano, i.e. la proporción de materia seca acumulada en el grano con respecto a la biomasa total o índice de cosecha (Passioura y Angus, 2010). La acumulación alta de biomasa en la planta y alto índice de cosecha podrían reducir el efecto negativo del déficit hídrico del suelo durante la floración y el llenado del grano. Por lo tanto, el objetivo de este estudio fue evaluar la respuesta, con base en el rendimiento de grano y sus componentes, y la densidad de raíces en condiciones de campo bajo riego y secano durante las etapas fenológicas de floración y de llenado del grano en germoplasma de arroz.
Materiales y Métodos
Sitio experimental
Un experimento se estableció en el Campo Agrícola Experimental del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) en Zacatepec, Morelos, México (18º 39' N, 99º 12' O y 900 msnm) durante el ciclo primavera-verano de 2013. La localidad tiene un clima cálido subhúmedo con lluvias en verano (Awo (w)), una precipitación media anual de 839 mm, y temperatura media anual de 24.8 °C; la mayor proporción de la lluvia ocurre de mayo a octubre, con un periodo de sequía ‘intra-estival’ o ‘canícula’ en julio (García, 2004). El suelo tiene textura arcillosa y con mayores valores de capacidad de campo (CC) 54.8 %, del punto de marchitez permanente (PMP) 32.2 %, pH (7.6), conductividad eléctrica (CE=0.59 dS m-1) y contenidos de materia orgánica (MO=2.2 %), nitrógeno (N=0.11 %), fósforo (P=14.3 ppm) y potasio (K=0.29 ppm) en el estrato de 0-20 cm comparados con los observados en el estrato de 20-40 cm (CC=53 %, PMP=31.2 %, pH=7.5, CE=0.45 dS m-1, MO=1 %, N=0.07 %, P=5.1 ppm y K=0.15 ppm); la densidad aparente (1.9 g cm-3) fue mayor en el estrato de 20-40 cm que en el estrato de 0-20 cm (1.85 g cm-3) (Laboratorio de Edafología, Colegio de Postgraduados. Montecillo, México).
Material genético
El germoplasma evaluado incluyó ocho líneas F6 de arroz con características de alto rendimiento y resistencia a Tagosodes oryzicolus y Pyricularia oryzae; así como con calidad de grano superior definida por su contenido de amilosa, apariencia del grano procesado, alta recuperación de granos enteros y tolerancia al retraso en cosecha (Chatel et al., 2010). Las líneas proceden de los viveros del Fondo Latinoamericano para Arroz de Riego (FLAR; Santa Rosa, Cali, Colombia; Cuadro 1). El material genético se preseleccionó por rendimiento de grano en ambientes de trópico húmedo, subhúmedo y seco en el ciclo primavera-verano (P-V) de 2012 en Palizada, Campeche; Zacatepec, Morelos; Cárdenas, Tabasco y Tecomán, Colima, México. El testigo fue la variedad comercial El Silverio de Oaxaca, México (Cuadro 1).
Cuadro 1 Material genético utilizado en el experimento de campo.
| No. de genotipo |
Genealogía | Origen | Rendimiento de grano† (t ha-1) |
Días a antesis† |
Resistencia genética† |
| 1 | FL04621-2P-1-3P-3P-M | P-V 2005 | 6.8 | 85 | T. oryzicolus y P. oryzae |
| 2 | FL05392-3P-12-2P-2P-M | P-V 2006 | 13.2 | 90 | " |
| 3 | FL06747-4P-10-5P-3P-M | P-V 2007 | 10.5 | 89 | " |
| 4 | FL07201-6P-5-3P-3P-M | P-V 2008 | 9.4 | 84 | " |
| 5 | FL08224-3P-2-1P-2P-M | P-V 2009 | 10.9 | 90 | " |
| 6 | FL08378-3P-5-2P-2P-M | P-V 2009 | 11.4 | 87 | " |
| 7 | FL08224-3P-2-1P-3P-M | P-V 2009 | 15.9 | 92 | " |
| 8 | FL10129-12P-4-2P-3P-M | P-V 2011 | 12.1 | 88 | " |
| 9 | El Silverio (Testigo) | P-V 2011 | 12.0 | 91 | P. oryzae |
†Datos proporcionados por el M.C. Leonardo Hernández Aragón, Líder Nacional del Programa de Arroz, INIFAP, Zacatepec, Morelos.
Tratamientos y diseño experimental
El experimento consistió en dos tratamientos de humedad edáfica: 1) riego por inundación (con la aplicación de agua por gravedad cada semana desde la siembra hasta la madurez fisiológica y mantener el suelo inundado permanentemente) y 2) secano, donde el riego se suspendió a los 26 d después de la siembra (dds) hasta la madurez fisiológica. Los tratamientos se aleatorizaron en un diseño experimental de bloques completos al azar con tres repeticiones en cada condición de humedad y la unidad experimental fue seis surcos de 5 m de largo separados a 0.2 m. La siembra se hizo en forma directa en suelo seco a chorrillo el 10 de junio de 2013, con una densidad de siembra de 100 kg ha-1 y dosis de fertilización de 180-40-40, empleando urea, superfosfato de calcio triple y cloruro de potasio, respectivamente. Un tercio del N y todo el P y K se aplicaron a la siembra; el segundo tercio del N se aplicó al iniciar el amacollamiento (25 dds); el último tercio se aplicó durante el ‘embuche’ (70 dds).
Las malezas se controlaron con dos aplicaciones de herbicida; la primera en preemergencia, 3 d después del riego de siembra (Ronstar®, 2 L ha-1); la segunda a los 21 dds (Propanil® + 2,4-D diclorofenoxiacético® a una dosis de 6.0 y 1.5 L ha-1, respectivamente). El control de Rupela albinella, Spodoptera frugiperda Smith y Oebalus insularis, se realizó al aplicar Cipermetrina® a razón de 0.25 L ha-1, a los 66 y 90 dds.
Datos meteorológicos
Los datos de temperatura máxima y mínima del aire, así como la precipitación acumulada cada 10 d, se obtuvieron de una estación meteorológica localizada a 50 m de distancia del experimento, dentro del Campo Experimental del INIFAP en Zacatepec, Morelos.
Contenido hídrico del suelo
El contenido de humedad aprovechable (HA, %) del suelo se determinó sólo en el tratamiento de secano cada 10 d en los estratos 0-20 y 20-40 cm desde la emergencia de las plántulas hasta la madurez fisiológica, de acuerdo con el método gravimétrico [% (HA) = (peso de suelo húmedo - peso de suelo seco/peso de suelo seco) 100].
Variables medidas
La biomasa aérea (BM, g m-2), el rendimiento de grano (RG, g m-2), el índice de cosecha (IC=RG/BM), el número de panículas m-2 (P M-2), el número de granos por panícula (GP), el número de granos m-2 (G M-2) y el peso de mil granos (PMG, g) se evaluaron a la cosecha.
La densidad de raíces (DR, cm cm-3) se calculó a partir de muestras de suelo de todas las parcelas del experimento. Cuatro muestras de suelo se extrajeron de cada parcela con una barrena del tipo Veihmeyer con boquilla de 2.1 cm de diámetro en los estratos de 0-15, 15-30, 30-50 y 50-70 cm; dos sobre la línea de plantas y dos entre líneas de plantas. En las muestras de suelo completamente secas se separaron los fragmentos de raíz con unas pinzas de depilar en cada muestra y se colocaron en viales de plástico con 50 mL de agua destilada. La densidad de raíces (cm raíz cm-3 suelo) se estimó de acuerdo con el método descrito por Tennant (1975) y López-Castañeda y Richards (1994).
Análisis estadístico
Los datos se analizaron mediante el ANDEVA para todas las variables medidas con el programa SAS versión 9.1 para Windows (SAS, 2009) en forma individual para cada tratamiento de humedad edáfica y en forma combinada, para determinar la significancia de la interacción genotipo x ambiente. La comparación de medias se realizó con la prueba de Tukey (DMSH, p ≤ 0.05). Adicionalmente, se realizó un análisis multivariado de componentes principales del rendimiento de grano versus los componentes del rendimiento y la densidad de raíces, para los datos obtenidos en riego y secano (STATGRAPHICS, 2010).
Resultados y Discusión
Precipitación, temperatura y humedad edáfica
La lluvia acumulada en el curso del experimento ascendió a 645 mm, distribuidos irregularmente durante el mismo. La lluvia se redujo considerablemente desde 50 a 60 dds (fin de la fase vegetativa), 80 a 90 dds (emergencia de la panícula) y 110 a 140 dds (antesis y llenado de grano) (Figura 1a). La lluvia registrada fue inferior al requerimiento mínimo de humedad (> 800 mm) establecido en las zonas productoras de arroz más aptas de Morelos, con rendimientos potenciales de 9.8 a 17.1 t ha-1 (Osuna, 2000). El estrés por sequía durante el crecimiento vegetativo, en especial en la fase inicial, la floración y períodos terminales, puede interrumpir la iniciación floral causando esterilidad de la espiguilla y deficiencias en el llenado del grano, lo que resulta en menor peso del grano y reducción en el rendimiento del arroz (Kamoshita et al, 2008; Kumar et al., 2014).
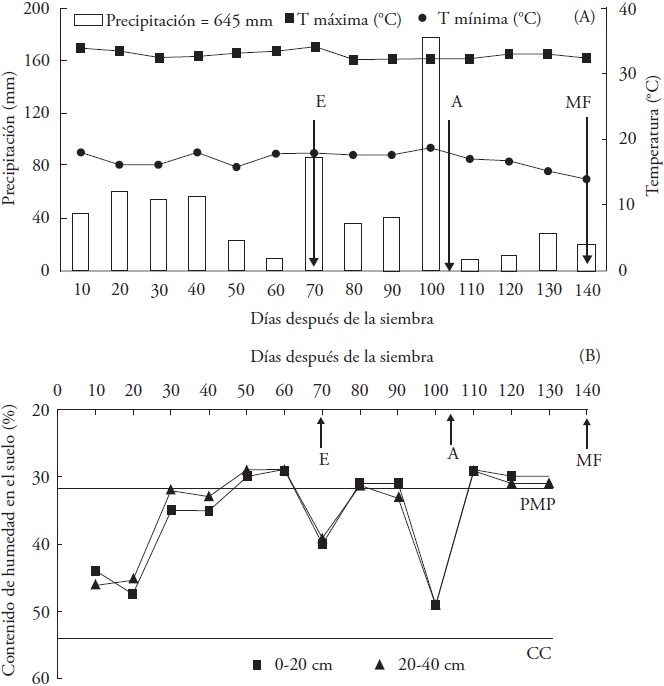
Figura 1 Promedio de temperatura máxima y mínima del aire, y precipitación acumulada (A) y contenido de humedad del suelo en los estratos de 0-20 y 20-40 cm en el tratamiento de secano (B), determinados cada 10 d durante el experimento en el ciclo primavera-verano 2013. Zacatepec, Morelos. PMP = Porcentaje de marchitamiento permanente; CC = Capacidad de campo. Promedio del número de días a embuche (E), antesis (A) y madurez fisiológica.
Las temperaturas máxima y mínima del aire mostraron poca variación con promedios de 17 y 33 °C, respectivamente (Figura 1A); estos valores se ubican entre las temperaturas mínima (13.5 °C) y máxima (35.4 °C) favorables para los procesos de germinación de las semillas, iniciación foliar, crecimiento de la raíz y órganos aéreos de la planta e iniciación de la panícula (Sánchez et al., 2014). Los valores de temperaturas también fueron favorables para la floración, fertilidad de las espiguillas y rendimiento de grano (Jagadish et al., 2015). Las temperaturas mayores a 35 °C causan esterilidad en las espiguillas al afectar la emergencia de la panícula (Cho y Oki, 2012).
En la parcela experimental con tratamiento de riego el suelo permaneció inundado durante el ciclo biológico de las plantas (datos no mostrados). En contraste, el tratamiento de secano exhibió, disponibilidad de humedad edáfica que varió con la cantidad y distribución de la lluvia, se registraron periodos de sequía con contenidos hídricos del suelo inferiores al PMP, el primero al final de la fase vegetativa (50-60 dds), luego durante la emergencia de la panícula (80 a 90 dds) y, por último, durante la antesis y el llenado del grano (110-140 dds) (Figura 1B). El mayor déficit hídrico se observó en el estrato de 0-20 cm, donde se concentra más del 50 % de la masa de raíces (López-Castañeda y Richards, 1994).
Rendimiento de grano, sus componentes y densidad de raíces en condiciones de riego
En condiciones de riego la variabilidad entre genotipos en cuanto a RG y sus componentes fue amplia. El RG, la BM, el IC, P M-2, G M-2, PMG y GP variaron de 800 a 1606, de 1015 a 1425, de 43 a 53, de 125 a 257, de 15422 a 45538, de 28 a 37 y de 123 a 187, respectivamente; la mayor variación se observó en el rendimiento de grano y el número de granos m-2 (Cuadro 2). Las líneas 2 y 7 produjeron el mayor RG acompañado de los más altos valores BM, IC, G M-2, P M-2, PMG y GP (Cuadro 2). El rendimiento de grano y sus componentes en el testigo fueron inferiores a los registrados para las líneas experimentales. El alto rendimiento de grano de las líneas 2 y 7 confirmó el comportamiento sobresaliente mostrado los ciclos de preselección (Cuadro 1). El rendimiento de grano de las líneas 2 y 7 fue 44 % mayor al rendimiento obtenido con las dos mejores líneas experimentales de arroz en promedio de tres ambientes (Nayarit, Michoacán y San Luis Potosí) en México (Barrios et al., 2016). Las diferencias en rendimiento de grano podrían deberse al mayor potencial genético de rendimiento y resistencia a enfermedades en el germoplasma empleado aquí, que el estudiado por Barrios et al. (2016); así como la alta habilidad de amacollamiento, grano grande alargado y estabilidad del rendimiento en todos los ambientes (Salcedo y Barrios, 2013).
Cuadro 2 Rendimiento de grano y sus componentes, y densidad de raíces en distintos estratos del suelo en riego. Ciclo primavera-verano 2013. Zacatepec, Morelos, México.
| No. de genotipo |
RG¶ (g m-2) |
BM¶ (g m-2) |
IC¶ (%) |
P M-2¶ | G M-2¶ | PMG¶ (g) |
GP¶ | Densidad de raíces (cm cm-3) | |||
| 0-15 cm | 15-30 cm | 30-50 cm | 50-70 cm | ||||||||
| 2 | 1606a | 1425ab | 51a | 257a | 45538a | 37a | 177b | 0.28a | 0.17bc | 0.09b | 0.09a |
| 7 | 1590a | 1425ab | 53a | 227b | 42458a | 33b | 187a | 0.22c | 0.19ab | 0.10ab | 0.05b |
| 3 | 1364b | 1165b | 52a | 143c | 18902de | 31bc | 132de | 0.29a | 0.16bc | 0.12a | 0.04b |
| 6 | 1330b | 1250bc | 51a | 143c | 22061cd | 32b | 155c | 0.29a | 0.15bc | 0.09ab | 0.05b |
| 5 | 1300b | 1254bc | 51a | 142c | 22682c | 31bc | 160c | 0.23bc | 0.22a | 0.08b | 0.04b |
| 4 | 1207c | 1172cd | 51a | 130c | 16871ef | 29cd | 130def | 0.27ab | 0.14c | 0.09ab | 0.06b |
| 1 | 972d | 1015d | 49b | 125c | 15422f | 28d | 123f | 0.28a | 0.18ab | 0.09ab | 0.05b |
| 8 | 890e | 1119cd | 45b | 230b | 31864b | 28d | 139d | 0.25abc | 0.16bc | 0.10ab | 0.04b |
| 9 (Testigo) | 800f | 1192cd | 43b | 131c | 16503ef | 28d | 126ef | 0.28a | 0.15bc | 0.11a | 0.05b |
| Media general | 1229 | 1247 | 50 | 170 | 25811 | 31 | 148 | 0.26 | 0.16 | 0.10 | 0.05 |
Medias con distinta letra en una hilera son estadísticamente diferentes (Tukey, p≤0.05). ¶Ver Materiales y Métodos.
El rango de variación genética para densidad de raíces fue de 0.22 a 0.29, 0.15 a 0.22. 0.08 a 0.12 y 0.4 a 0.9 cm cm-3, en los estratos: 0-15, 15-30, 30-50 y 50-70 cm de profundidad, respectivamente; las líneas 2, 3, 6, 4, 1 y 9 (testigo); 7, 5 y 1; 7, 3, 6, 1, 8 y 9 (testigo); y la línea 2 mostraron la mayor densidad de raíces en los estratos de 0-15, 15-30, 30-50 y 50-70 cm, respectivamente. La línea testigo sólo tuvo alta densidad de raíces en los estratos de 0-15 y 30-50 cm (Cuadro 3). Además, se determinó que la línea 2 tuvo mayor densidad de raíces en los estratos de 0 a 70 cm, lo que podría ser ventajoso para una mayor captura de agua y nutrientes del suelo. Ahmadi et al. (2014) indicaron que el sistema de raíces tiene una función crucial en la extracción de agua y nutrientes a través del complejo que forman con las propiedades biogeoquímicas del suelo y de mantener estas funciones bajo un amplio rango de escenarios de estrés, para asegurar la sobrevivencia de la planta y su reproducción; esta función es aún más importante debido a la creciente inestabilidad del clima y las limitaciones en el uso de fertilizantes y riego en los sistemas de cultivo.
Cuadro 3 Rendimiento de grano y sus componentes, y densidad de raíces en distintos estratos del suelo en condiciones de secano. Ciclo primavera-verano 2013. Zacatepec, Morelos, México.
| No. de genotipo |
RG¶ (g m-2) |
BM¶ (g m-2) |
IC¶ (%) |
P M-2¶ | G M-2¶ | PMG¶ (g) |
GP¶ | Densidad de raíces (cm cm-3) | |||
| 0-15 cm | 15-30 cm | 30-50 cm | 50-70 cm | ||||||||
| 2 | 939a | 1433a | 40ab | 129a | 15045a | 26a | 117a | 0.35d | 0.33a | 0.21a | 0.13a |
| 7 | 980a | 1369a | 42a | 128a | 15131a | 26a | 118a | 0.53abc | 0.26bc | 0.18bc | 0.08d |
| 3 | 698cb | 1165b | 37ab | 121ab | 12359b | 21b | 102bc | 0.37bcd | 0.22c | 0.17c | 0.09bcd |
| 6 | 743b | 1162b | 39ab | 110c | 11515bc | 23b | 104abc | 0.38bcd | 0.25c | 0.18bc | 0.07d |
| 5 | 627cd | 1097bc | 37b | 113bc | 12557b | 21b | 111ab | 0.36cd | 0.26c | 0.20ab | 0.08d |
| 4 | 621cd | 991cd | 38ab | 116bc | 12500b | 21b | 108ab | 0.54ab | 0.24c | 0.18bc | 0.11abc |
| 1 | 385e | 924d | 30c | 92d | 8290d | 21b | 90c | 0.53abc | 0.30ab | 0.17c | 0.09bcd |
| 8 | 554d | 1030bc | 35b | 100d | 9900cd | 22b | 99bc | 0.57a | 0.22c | 0.18bc | 0.09cd |
| 9 (Testigo) | 203f | 937d | 18d | 95d | 11101bc | 21b | 118a | 0.47abcd | 0.23c | 0.17c | 0.13a |
| Media general | 638 | 1123 | 35 | 112 | 12044 | 22 | 107 | 0.45 | 0.26 | 0.18 | 0.10 |
Medias con distinta letra en una hilera son estadísticamente diferentes (Tukey, p≤0.05). ¶Ver Materiales y Métodos.
Rendimiento de grano, sus componentes y densidad de raíces en condiciones de secano
En condiciones de secano el rendimiento de grano fue afectado severamente por el déficit hídrico, obteniéndose en promedio 638 g m-2 vs. 1229 g m-2 en riego. El rango de variabilidad en el RG, BM, IC, P M-2, G M-2, PMG y GP fue de 203 a 980, de 924 a 1433, de 30 a 42, de 92 a 129, de 8290 a 15131, de 21 a 26 y de 90 a 118, respectivamente. La BM y el IC mostraron un rango de variación más amplio en sequía (BM=924 a 1433 e IC=30 a 42) que en riego (BM=1015 a 1425 e IC=43 a 53); el RG y G M-2 fueron los componentes con mayor variación genética. Las líneas 2 y 7 produjeron el mayor RG, BM, IC y G M-2; adicionalmente, la línea 2 mostró alto número de P M-2 y PMG, y la línea 7 mostró alto número de GP (Cuadro 3). El IC promedio de las líneas 3, 6, 5 y 4 fue similar al de las líneas 2 y 7. El RG y todos sus componentes en el testigo fueron inferiores a las líneas experimentales. El rendimiento superior de las líneas 2 y 7 tanto en riego como en sequía, confirmó el comportamiento sobresaliente mostrado por ellas en los ciclos de preselección (Cuadros 1 y 2). Xangsayasane et al. (2014) obtuvieron resultados similares al estudiar el comportamiento de 227 líneas experimentales de arroz en tres tratamientos: sequía terminal, sequía intermitente y riego, y reducción del rendimiento de 34 y 59 % en los tratamientos de sequía terminal y sequía intermitente con respecto a riego. La reducción en el RG se asoció con una disminución en el porcentaje de granos llenos, IC y P M-2.
La densidad de raíces varió de 0.35 a 0.57, 0.22 a 0.33, 0.17 a 0.21 y 0.07 a 0.13 cm cm-3 en los estratos de 0-15, 15-30, 30-50 y 50-70 cm, respectivamente; las líneas 4 y 8; 2 y 1; 2 y 5, y 2 y testigo tuvieron mayor densidad de raíces en los estratos de 0-15, 15-30, 30-50 y 50-70 cm, respectivamente. El testigo mostró alta densidad de raíces sólo en el estrato de 50-70 cm (Cuadro 3). Las raíces tienen una función importante en el mantenimiento del equilibrio hídrico de la planta. En condiciones de déficit de humedad, el desequilibrio hídrico sobreviene cuando la tasa de evaporación de la planta es mayor que la tasa de absorción de las raíces (Boyer y Silk, 2004); en tales condiciones, mayores peso seco de raíz, longitud de raíz, porcentaje de raíz, número de raíces nodales, peso específico de raíces, cociente raíz/parte aérea y capacidad de ajuste osmótico se asocian, individual o conjuntamente, con la resistencia del arroz a sequía (Fageria y Moreira, 2011; Feng et al., 2012; Gowda et al., 2011; Henry et al., 2016).
Comparación riego vs. secano
Las deficiencias hídricas del suelo afectaron diferencialmente al germoplasma con base en el RG y sus componentes y la DR en comparación con la condición de riego. El déficit hídrico en secano redujo el rendimiento y sus componentes y promovió el mayor desarrollo del sistema radical (Cuadro 4). La reducción debida al estrés hídrico fue más severa en el RG (48 %) y G M-2 (53 %) que en los otros componentes del rendimiento, mientras que P M-2, IC, PMG y GP disminuyeron 34, 30, 29 y 28 %, respectivamente. La BM fue la característica menos afectada por la sequía (10 %) (Cuadro 4). En 20 líneas de arroz seleccionadas por alta BM y sometidas a condiciones de inundación y secano en campo, la mayoría de los genotipos tuvieron buen comportamiento en condiciones de inundación y sólo algunos se comportaron bien en condiciones de estrés hídrico. El germoplasma susceptible al estrés hídrico mostró una reducción significativa en altura de planta, número de macollos por planta y biomasa aérea total en la madurez fisiológica, aunque fue más tardío a floración (Kondhia et al., 2015).
Cuadro 4 Rendimiento de grano y sus componentes, y densidad de raíces en distintos estratos del suelo de todos los genotipos en riego y sequía. Ciclo primavera-verano 2013. Zacatepec, Morelos, México.
| Tratamiento de humedad |
RG¶ | BM¶ | IC¶ | P M-2¶ | G M-2¶ | PMG¶ | GP¶ | Densidad de raíces (cm cm-3) | |||
| 0-15 cm |
15-30 cm |
30-50 cm |
50-70 cm |
||||||||
| Riego | 1229a | 1247a | 50a | 170a | 25811a | 31ª | 148a | 0.26b | 0.16b | 0.10b | 0.05b |
| Sequía | 638b | 1123b | 35b | 112b | 12044b | 22b | 107b | 0.45a | 0.26a | 0.18a | 0.10a |
Medias con distinta letra en una hilera son estadísticamente diferentes (Tukey, p≤0.05). ¶Ver Materiales y Métodos.
En contraste con lo observado en el rendimiento y sus componentes, el déficit hídrico aumentó la densidad de raíces respecto a riego en todos los estratos del suelo estudiados. La mayor concentración de raíces en promedio ocurrió en el estrato de 0-30 cm tanto en riego (74 %) como en sequía (72 %) (Cuadro 4). Kondo et al. (2003) estudiaron el desarrollo de las raíces en la etapa de floración en once genotipos de arroz en Filipinas y el efecto del genotipo explicó la mayor proporción de la variación en el número de raíces nodales, peso específico de la raíz y cociente raíz/parte aérea, mientras que el efecto ambiental fue relativamente grande sobre la profundidad de la raíz y el peso seco total de raíces. En el arroz, el crecimiento del sistema radical es influenciado por el nivel de nutrición y humedad en el suelo. En la Provincia de Jiangsu, China, la variedad Wuyunging 24, sometida a tres dosis de nitrógeno (100, 200 y 300 kg ha-1) y tres regímenes de humedad en el suelo (riego alternado con sequía moderada, riego alternado con sequía severa e inundación continua), exhibió peso seco de raíces en los tratamientos de inundación continua y riego alternado con sequía moderada con 200 y 300 kg N ha-1 mayor que en riego alternado con sequía severa y 100 kg N ha-1 en la etapa de emergencia de la panícula. Por el contrario, el cociente raíz/parte aérea en el tratamiento de riego alternado con sequía severa y 100 kg N ha-1 fue mayor que en los otros tratamientos, indicando que el crecimiento de la raíz en relación con la parte aérea es menos inhibido en condiciones de riego alternado con sequía severa (Wang et al., 2016).
Interacción variedades x niveles de humedad del suelo
La interacción variedades x niveles de humedad edáfica se analizó contrastando el RG y sus componentes observados bajo riego con aquellos bajo condiciones de secano. El RG y G M-2 mostraron las mayores diferencias riego - secano en promedio de todos los genotipos, seguidos por la BM, P M-2, GP, IC y PMG (Cuadro 5). Las líneas 2, 7, 3, 6, 5, 4, 1 y el testigo; el testigo; la línea 2; las líneas 2 y 7; la línea 2; la línea 7; y la línea 5, mostraron mayor diferencia riego - secano para el RG; BM e IC; P M-2; G M-2; PMG; y GP, respectivamente (Cuadro 5). Cuando la deficiencia hídrica ocurre durante la fase vegetativa del arroz, su efecto en el desarrollo subsecuente y en el RG es mínimo (30 %), pero cuando ocurre durante el desarrollo de la panícula, su efecto en el RG es significativo (70 %), retrasando la antesis (22-28 d) y reduciendo el número de espiguillas por panícula (60 %) y el número de granos m-2 (100 %) o cuando ocurre durante el llenado del grano, su efecto también es significativo en el RG (50 %), retrasando la madurez fisiológica (10-20 d) y disminuyendo el número de granos (40 %) y el peso individual del grano (20 %) (Boonjung y Fukai, 1996). En las tierras bajas del sureste de Asia se determinó que la sequía intermitente y terminal en la época lluviosa redujeron 13 y 35 % el rendimiento de grano de arroz, mientras que en la época seca el rendimiento disminuyó 34 % bajo sequía intermitente y 59 % con sequía terminal. Estos resultados permitieron concluir que la selección de genotipos tolerantes a sequía puede lograrse alternando la evaluación del germoplasma en condiciones de sequía intermitente en la época seca y con sequía terminal en la época lluviosa (Xangsayasane et al., 2014).
Cuadro 5 Diferencias riego - secano para el rendimiento y sus componentes, y densidad de raíces en distintos estratos del suelo. Ciclo primavera-verano 2013. Zacatepec, Morelos, México.
| No. de genotipo |
RG¶ | BM¶ | IC¶ | P M-2¶ | G M-2¶ | PMG¶ | GP¶ | Densidad de raíces (cm cm-3) | |||
| 0-15 cm | 15-30 cm | 30-50 cm | 50-70 cm | ||||||||
| 2 | 667a | 109b | 11cd | 128a | 30493ª | 11.3a | 61ab | 0.07c | 0.16a | 0.13a | 0.05bc |
| 7 | 610a | 56b | 11cd | 99b | 27327ª | 6.7cd | 69a | 0.31a | 0.07cd | 0.08c | 0.03bc |
| 3 | 666a | 92b | 15bc | 22cd | 6544e | 10.0abc | 30ef | 0.08c | 0.05d | 0.06c | 0.05bc |
| 6 | 586a | 88b | 13cd | 32c | 10547c | 9.3abcd | 50bc | 0.09c | 0.10bc | 0.08bc | 0.03c |
| 5 | 673a | 156ab | 14cd | 29cd | 10126cd | 10.6ab | 49bcd | 0.13bc | 0.03d | 0.11ab | 0.04bc |
| 4 | 587a | 181 ab | 12cd | 14d | 4371e | 8.0abcd | 22fg | 0.27ab | 0.09bcd | 0.09abc | 0.05ab |
| 1 | 587a | 91b | 19ab | 33c | 7132de | 7.3bcd | 33def | 0.25abc | 0.12ab | 0.07c | 0.05bc |
| 8 | 336b | 89b | 10d | 130a | 21964b | 6.0d | 39cde | 0.32a | 0.06cd | 0.08bc | 0.06ab |
| 9 (Testigo) | 596a | 255a | 22a | 36c | 5402e | 7.7bcd | 8.3 g | 0.19abc | 0.07cd | 0.06c | 0.08a |
| Media general | 589 | 124 | 14 | 58 | 13767 | 8.5 | 40 | 0.18 | 0.085 | 0.083 | 0.048 |
Medias con distinta letra en una hilera son estadísticamente diferentes (Tukey, p≤0.05). ¶Ver Materiales y Métodos.
En promedio, para todos los genotipos, la densidad de raíces mostró mayores diferencias riego - secano en el estrato de 0-15 cm que en el resto (Cuadro 5). La línea 8, la línea 2 y el testigo, mostraron la mayor densidad de raíces en el estrato de 0-15, estratos 15-30 y 30-50 y 50-70 cm en comparación con el germoplasma restante, respectivamente (Cuadro 5). La mayor acumulación de biomasa en las raíces puede contribuir al mayor rendimiento de grano y eficiencia en el uso del agua en los sistemas agrícolas de secano. El mayor tamaño del sistema de raíces de la planta depende de mayores densidades de raíces, área superficial total de absorción y área activa de la superficie de absorción de las raíces (Chu et al., 2014). Un grupo de líneas derivadas de las cruzas entre O. sativa (IR64) y O. glaberrima (RAM90 y RAM54) mostraron mayor rendimiento de grano en riego y sequía comparados con el progenitor recurrente IR64 (susceptible a sequía) y menor temperatura del dosel vegetal bajo sequía; además, la densidad de raíces se asoció negativamente con la temperatura del dosel vegetal y el contenido de humedad en el suelo (Kijoji et al., 2014). Dado que también la relación entre la densidad de raíces y el rendimiento de grano en campo sólo fue consistente en algunas líneas, se confirmó la relación compleja entre el rendimiento de grano, el crecimiento de las raíces y los mecanismos fisiológicos que controlan el crecimiento del cultivo y la productividad en condiciones de campo.
Rendimiento de grano vs. componentes del rendimiento y densidad de raíces
El análisis de componentes principales (CP) del rendimiento de grano versus los componentes del rendimiento y la densidad de raíces, obtenidos para riego y secano mostró que los dos primeros CP explicaron el 83.6 % de la variación conjunta de los datos. El primer CP tuvo correlación positiva con el PMG (R=0.96) y el RG (R=0.93), mientras que el segundo CP mostró correlación positiva con la BM (R=0.64). La BM tuvo relación positiva con G M-2, P M-2 y GP, RG y PMG, y negativa con el IC; el IC mostró relación negativa con la DR, sobre todo a los 0-15 cm de profundidad del suelo (Figura 2). La asociación positiva entre el RG y sus componentes, BM y G M-2 puede ser útil para la selección en ambientes contrastantes en cuanto a la disponibilidad de humedad. Las líneas de arroz más sobresalientes en tres ambientes de México, además de alto rendimiento, mostraron altos porcentajes de recuperación de granos enteros y buena calidad para uso industrial en arroz (Barrios et al., 2016). En cebada cultivada en condiciones de riego, sequía y secano en México se determinó que el rendimiento de grano tuvo relación positiva y significativa con la biomasa aérea y el número de granos m-2. Así, la biomasa aérea a su vez, se asoció positiva y significativamente con el número de granos m-2, de tal forma que los genotipos con mayor rendimiento de grano mostraron mayor biomasa aérea y mayor número de granos m-2 (López-Castañeda, 2011).
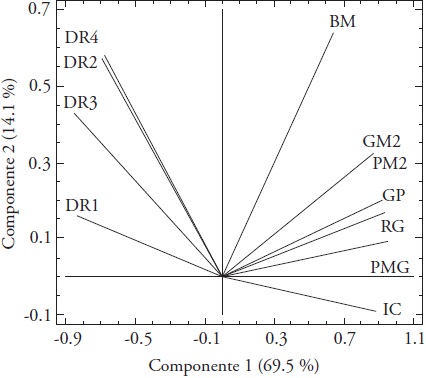
Figura 2 Biplot del rendimiento de grano versus componentes del rendimiento y densidad de raíces para nueve genotipos de arroz en la madurez; BM, biomasa aérea; GM2, número de granos m-2; PM2, número de panículas m-2; GP, número de granos por panícula; RG, rendimiento de grano; PMG, peso de mil granos; IC, índice de cosecha; DR1, densidad de raíces en el estrato de 0-15 cm; DR2, densidad de raíces en el estrato de 15-30 cm; DR3, densidad de raíces en el estrato de 30-50 cm; DR4, densidad de raíces en el estrato de 50-70 cm de profundidad del suelo.
Conclusiones
El déficit hídrico del suelo durante la floración y el llenado del grano disminuyó el rendimiento de grano y sus componentes en germoplasma de arroz cultivado en condiciones de secano en Zacatepec, Morelos, México, mientras que la deficiencia de humedad aumentó la densidad de raíces en todos los estratos del suelo estudiados. Las líneas 2 y 7 mostraron los mayores rendimientos de grano en riego y secano, así como valores superiores al promedio nacional en riego y secano. La biomasa aérea, el índice de cosecha, el número de panículas m-2, número de granos m-2 y número de granos por panícula, así como el peso de mil granos se asociaron significativamente con el rendimiento de grano en ambas condiciones de humedad. Las líneas más productivas podrían utilizarse como progenitores en los programas de mejoramiento y como variedades comerciales en los sistemas de producción de arroz, tanto en condiciones de riego como secano.

