










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgrociencia
On-line version ISSN 2521-9766Print version ISSN 1405-3195
Agrociencia vol.52 n.4 Texcoco May./Jun. 2018
Crop Science
Grain yield and its components, and root density in rice under irrigation and rainfed conditions
1 Campus Montecillo. Colegio de Postgraduados. 56230. Montecillo, Estado de México.
2 Campo Experimental Zacatepec, INIFAP, Zacatepec, Morelos, México.
3 Facultad de Ciencias Agrícolas, Universidad Autónoma del Estado de México, El Cerillo, Piedras Blancas, Toluca, Estado de México, México 50090. (clc@colpos.mx)
In Mexico, rice (Oryza sativa L.) is cultivated under irrigation and rainfed conditions with moisture restrictions and frequent periods of drought during the growing season. Grain yield (GY) is reduced severely when drought coincides with flowering and the grain-filling period. The objective of this study was to evaluate the GY and its components, and the root density (RD) in eight F6 lines of rice and one control (the national variety El Silverio), under irrigation (I) and rainfed (R) conditions in Zacatepec, Morelos, Mexico. The lines come from the nurseries of the Latin American Fund for Irrigation Rice (FLAR, for its acronym in Spanish; Santa Rosa, Cali, Colombia). The experiment was sown on June 10, 2013, with a seeding rate of 100 kg ha-1 and a fertilization rate of 180-40-40 of N-P-K. GY and the number of grains m-2 (G M-2) were the characteristics most affected by the water deficit in R. The RD in I (74 %) and R (72 %) was higher in the stratum of 0-30 cm; the RD in R (0.45, 0.26, 0.18 and 0.10 cm cm-3) was higher than in I (0.26, 0.16, 0.10 and 0.05 cm cm-3) in every stratum of the soil. Lines 2 and 7 were the most productive in I and R, and showed high aerial biomass (BM), harvest index (HI) and G M-2. Line 2; in addition, showed higher RD than the other lines in the strata of 15-30, 30-50 and 50-70 cm in I and R. The control showed lower GY and its components, and RD than the other genotypes. GY was positively related to its components in average of I and R. The most productive lines could be used commercially and as parents in a rice breeding program for irrigation and rainfed areas.
Key words: aerial biomass; root density; water deficit; harvest index; genotype x environment interaction; grain yield
En México, el arroz (Oryza sativa L.,) se cultiva en riego y secano con restricciones de humedad y periodos frecuentes de sequía durante el ciclo. El rendimiento de grano (RG) se reduce severamente cuando la sequía coincide con la floración y el periodo de llenado del grano. El objetivo de este estudio fue evaluar el RG y sus componentes, y la densidad de raíces (DR) en ocho líneas F6 de arroz y un testigo (la variedad nacional El Silverio), en condiciones de riego (R) y secano (S) en Zacatepec, Morelos, México. Las líneas provienen de los viveros del Fondo Latinoamericano para Arroz de Riego (FLAR; Santa Rosa, Cali, Colombia). La siembra se hizo el 10 de junio de 2013, con una densidad de siembra de 100 kg ha-1 y dosis de fertilización de 180-40-40 de N-P-K. El RG y el número de granos m-2 (G M-2) fueron las características más afectadas por el déficit hídrico en S. La DR en R (74 %) y S (72 %) fue mayor en el estrato de 0-30 cm; la DR en S (0.45, 0.26, 0.18 y 0.10 cm cm-3) fue mayor que en R (0.26, 0.16, 0.10 y 0.05 cm cm-3) en todos los estratos del suelo. Las líneas 2 y 7 fueron las más productivas en R y S, y mostraron alta biomasa aérea (BM), índice de cosecha (IC) y G M-2. La línea 2, además, mostró mayor DR que las otras líneas en los estratos de 15-30, 30-50 y 50-70 cm en R y S. El testigo mostró menor RG y sus componentes, y densidad de raíces que los otros genotipos. El RG se relacionó positivamente con sus componentes en promedio de R y S. Las líneas más productivas podrían utilizarse comercialmente y como progenitores en los programas de mejoramiento del rendimiento de grano de arroz para áreas de riego y secano.
Palabras clave: biomasa aérea; densidad de raíces; déficit hídrico; índice de cosecha; interacción genotipo x ambiente; rendimiento de grano
Introduction
In Mexico, rice is cultivated in an approximate area of 42,310 ha with an average domestic grain yield of 6.3 Mg ha-1 (SAGARPA, 2017). Of this surface, 75 % is cultivated under conditions of irrigation or permanent flooding during the cycle, with a mean yield of 6.2 Mg ha-1 and the remaining 25 % is cultivated under rainfed conditions with average yields of 4.4 Mg ha-1 (SAGARPA, 2017). Three agroclimate zones for rice cultivation are distinguished in the country: humid tropics, sub-humid tropics, and dry tropics. Each zone has different agronomic and production problems, among which the following stand out: restriction in water supply for irrigation; rain irregularity with frequent intermittent periods of intra-summer, terminal drought, or both; flooding provoked by hurricanes; stress due to high temperatures; biotic factors such as weeds, pests and diseases; deficiencies or nutritional toxicity; and high production costs (García et al., 2011; Barrios-Gómez et al., 2016). The state of Morelos is one of the zones in the dry tropics that stands out because of high grain yields under irrigation, with an average of 10.2 Mg ha-1 (SAGARPA, 2017), which is due, presumably, to the use of varieties with higher grain density and higher number of panicles per unit of area with direct sowing in the ‘high’ (1000 to 1400 masl) and ‘low’ (800 to 1000 masl) zones (Salcedo and Barrios, 2013).
Rice requires a high volume of water throughout its biological cycle and it is the only commercial cereal that is cultivated in soils saturated with water (in some systems, the plants remain submerged) during a part or all of the biological cycle; therefore, the adequate supply of water is the most important factor for crop production (Chatel et al., 2010). However, there are cultivation areas where grain production depends, to a great measure, on the rain during the cycle; in these conditions, the plants can experience periods of water stress, in addition to the stress caused by other biotic and abiotic factors that considerably reduce the yield and quality of the grain (Miranda-Domínguez et al., 2016). The water deficit, in and of itself, is the abiotic factor with the highest impact on the rainfed rice yield by affecting morphological (number of panicles and spikelets per panicle; plant height; aerial biomass; number of tillers, roots and leaves), physiological (photosynthesis, transpiration, stomatal conductance, efficiency in water use, relative water content, chlorophyll content, activity of the photosystem II, stability of the membrane, isotopic discrimination of carbon, and content of abscisic acid), biochemical (accumulation of proline, sugars, polyamines and antioxidants), and genomic (expression of genes that codify for transcription factors and defense proteins) characteristics of the plants (Shah, et al., 2011; Pandey and Shukla, 2015; Khondia et al., 2015).
The root perceives the soil water deficit, its key function is the absorption of water and soil nutrients, and it is a significant component of the adaptation of the plant in environments with moisture limitations. Obtaining new varieties with improved traits from root growth for water acquisition in soils in the process of drying could have a significant impact on the national and global food security (Lynch et al., 2014). Under these conditions, the grain yield could be improved by identifying in the plant the genetic and physiological factors that determine a higher biomass accumulation and partition of dry matter to the grain, i.e. the proportion of dry matter accumulated in the grain with regards to the total biomass or harvest index (Passioura and Angus, 2010). The high biomass accumulation in the plant and high harvest index could reduce the negative effect of the water deficit in the soil during flowering and grain filling. Therefore, the objective of this study was to evaluate the response of a group of rice genotypes, based on the grain yield and its components, and the root density, in field conditions under irrigation and rainfed during the phenologic stages of flowering and grain filling.
Materials and Methods
Experimental site
An experiment was established in the Experiment Agricultural Station of the National Institute of Forest, Agricultural and Livestock Research (Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, INIFAP) in Zacatepec, Morelos, Mexico (18º 39' N, 99º 12' W and 900 masl) during the 2013 spring-summer cycle. The locality has a warm sub-humid climate with summer rains (Awo (w)), mean annual precipitation of 839 mm, and mean annual temperature of 24.8 °C; the highest proportion of rain happens from May to October, with an ‘intra-summer’ or ‘midsummer’ drought period in July (García, 2004). The soil has clay texture with higher values of field capacity (FC) 54.8 %, of permanent wilting point (PWP) 32.2 %, pH (7.6), electric conductivity (EC = 0.59 dS m-1) and organic matter (OM = 2.2 %), nitrogen (N = 0.11 %), phosphorus (P = 14.3 ppm) and potassium (K = 0.29 ppm) contents in the 0-20 cm stratum, compared to those observed in the 20-40 cm stratum (FC = 53 %, PWP = 31.2 %, pH = 7.5, EC=0.45 dS m-1, OM = 1 %, N = 0.07 %, P = 5.1 ppm and K = 0.15 ppm); the apparent density (1.9 g cm-3) was higher in the 20-40 cm stratum than in the 0-20 cm stratum (1.85 g cm-3) (Laboratorio de Edafología, Colegio de Postgraduados. Montecillo, México).
Genetic material
The germplasm evaluated included eight F6 lines of rice with characteristics of high yield and resistance to Tagosodes oryzicolus and Pyricularia oryzae; as well as, with higher grain quality defined by its content of amylose, appearance of the processed grain, high recovery of full grains, and tolerance to harvest delay (Chatel et al., 2010). The lines come from the nurseries of the Latin American Fund for Irrigation Rice (Fondo Latinoamericano para Arroz de Riego, FLAR; Santa Rosa, Cali, Colombia; Table 1). The genetic material was preselected based on the grain yield in humid tropics, sub-humid and dry environments in the 2012 spring-summer cycle (S-S) in Palizada, Campeche; Zacatepec, Morelos; Cárdenas, Tabasco; and Tecomán, Colima, Mexico. The control was the commercial variety El Silverio from Oaxaca, Mexico (Table 1).
Table 1 Genetic material used in the field experiment.
| No. de genotipo |
Genealogía | Origen | Rendimiento de grano† (t ha-1) |
Días a antesis† |
Resistencia genética† |
| 1 | FL04621-2P-1-3P-3P-M | P-V 2005 | 6.8 | 85 | T. oryzicolus y P. oryzae |
| 2 | FL05392-3P-12-2P-2P-M | P-V 2006 | 13.2 | 90 | " |
| 3 | FL06747-4P-10-5P-3P-M | P-V 2007 | 10.5 | 89 | " |
| 4 | FL07201-6P-5-3P-3P-M | P-V 2008 | 9.4 | 84 | " |
| 5 | FL08224-3P-2-1P-2P-M | P-V 2009 | 10.9 | 90 | " |
| 6 | FL08378-3P-5-2P-2P-M | P-V 2009 | 11.4 | 87 | " |
| 7 | FL08224-3P-2-1P-3P-M | P-V 2009 | 15.9 | 92 | " |
| 8 | FL10129-12P-4-2P-3P-M | P-V 2011 | 12.1 | 88 | " |
| 9 | El Silverio (Testigo) | P-V 2011 | 12.0 | 91 | P. oryzae |
†Data provided by the M.Sc. Leonardo Hernández Aragón, National Leader of the Rice Program, INIFAP, Zacatepec, Morelos.
Treatments and experimental design
The experiment had two treatments of soil moisture: 1) flood irrigation (with the application of water by gravity each week since sowing until physiological maturity and maintaining the soil permanently flooded), and 2) rainfed, where irrigation was suspended 26 d after sowing (das) until physiological maturity. The treatments were randomized in an experimental design of complete random blocks with three replicates in each condition of soil moisture and the experimental unit was six furrows of 5 m long separated by 0.2 m. The sowing was made directly on dry soil by runs on June 10, 2013, with a seeding rate of 100 kg ha-1 and fertilization of 180-40-40, using urea, triple calcium superphosphate and potassium chloride, respectively. A third part of the N and all of the P and K were applied to the sowing; the second third of the N was applied at the beginning of tillering (25 das); the last third was at the booting stage (70 das).
The weeds were controlled with two applications of herbicides; the first at pre-emergence, 3 d after the sowing irrigation (Ronstar®, 2 L ha-1); the second at 21 das (Propanil® + 2,4-D dichlorophenoxyacetic® at a dose of 6.0 and 1.5 L ha-1, respectively). The control of Rupela albinella, Spodoptera frugiperda Smith and Oebalus insularis was performed when applying Cypermethrin® at a rate of 0.25 L ha-1, at 66 and 90 das.
Meteorological data
The data of maximum and minimum air temperature, as well as the accumulated precipitation every 10 d, were obtained from a meteorological station located 50 m from the experiment, inside the INIFAP Experiment Field Station in Zacatepec, Morelos.
Soil water content
The available moisture content (HA, %) of the soil was determined only in the rainfed treatment every 10 d in the strata of 0-20 and 20-40 cm since the emergence of the seedlings until physiological maturity, according to the gravimetric method [% (HA) = (weight of humid soil - weight of dry soil / weight of dry soil) 100].
Measured variables
The aerial biomass (BM, g m-2), grain yield (GY, g m-2), harvest index (HI=GY/BM), number of panicles m-2 (P M-2), number of grains per panicles (GP), number of grains m-2 (G M-2) and thousand grains weight (TGW, g) were evaluated at harvest.
The root density (RD, cm cm-3) was calculated from soil samples from all the experimental plots. Four soil samples were extracted from each plot with a Veihmeyer-type borer with a nozzle of 2.1 cm diameter in the 0-15, 15-30, 30-50 and 50-70 cm strata; two on the line of plants and two between lines of plants. When the soil samples had completely dried, root fragments were separated with tweezers from each sample and placed in plastic vials with 50 mL of distilled water. Then, root density (cm root cm-3 soil) was estimated according to the method outlined by Tennant (1975) and López-Castañeda and Richards (1994).
Statistical analysis
The data were analyzed through ANOVA for all the variables with SAS software version 9.1 for Windows (SAS, 2009), individually for each treatment of soil moisture and combined, to determine the significance of the genotype x environment interaction. Means comparison was carried out with Tukey test (DMSH, p ≤ 0.05). Additionally, a multivariate analysis of principal components of grain yield versus yield components and root density was carried out, for the data obtained in irrigation and rainfed conditions (STATGRAPHICS, 2010).
Results and Discussion
Precipitation, temperature and soil moisture
The amount of rainfall accumulated during the experiment reached 645 mm, with an irregular distribution. The rain was reduced considerably from 50 to 60 das (end of the vegetative phase), 80 to 90 das (emergence of the panicle) and 110 to 140 das (anthesis and grain filling) (Figure 1a). The rain recorded was lower than the minimum moisture requirement (> 800 mm) established in the most apt rice-producing zones in Morelos, with potential yields of 9.8 to 17.1 t ha-1 (Osuna, 2000). The water stress from drought during the vegetative growth, mostly in the initial phase, the flowering and terminal periods, can interrupt the floral initiation causing sterility of the spikelets and deficiencies in the grain filling, which results in lower grain weight and reduction in rice yield (Kamoshita et al, 2008; Kumar et al., 2014).
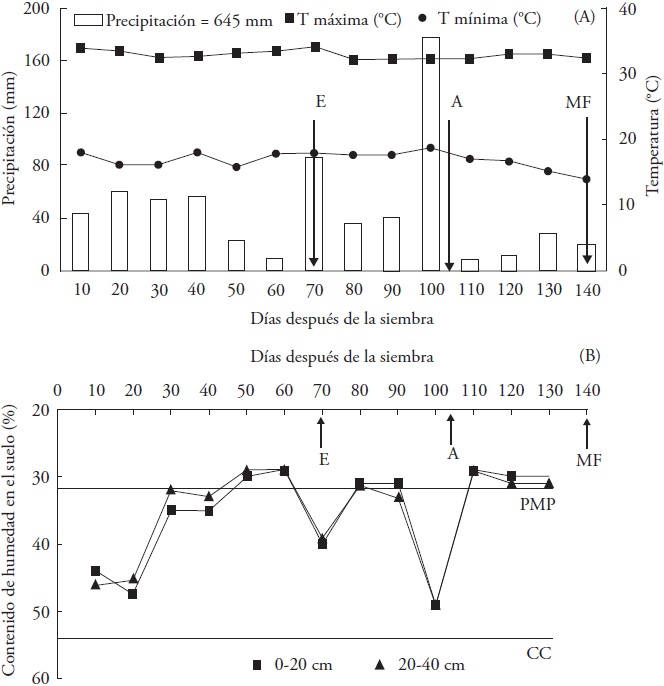
Figure 1 Average of maximum and minimum air temperatures, and accumulated precipitation (A) and soil moisture content in the 0-20 and 20-40 cm strata in the rainfed treatment (R), determined every 10 d during the experiment in the 2013 spring-summer cycle. Zacatepec, Morelos. PWP = Permanent wilting percentage; FC = Field capacity. Average number of days to booting (B), anthesis (A) and physiological maturity (PM).
The maximum and minimum air temperatures showed low variation with averages of 17 and 33 °C, respectively (Figure 1A); these values are between the minimum (13.5 °C) and maximum (35.4 °C) temperatures favorable for processes of seed germination, foliar initiation, root growth, and aerial organs of the plant and initiation of the panicle (Sánchez et al., 2014). The temperature values were also favorable for flowering, fertility of the spikelets, and grain yield (Jagadish et al., 2015). Temperatures higher than 35 °C cause sterility in the spikelets by affecting the emergence of the panicle (Cho and Oki, 2012).
In the experimental plot with irrigation treatment, the soil remained flooded during the biological cycle of the plants (data not shown). In contrast, the rainfed treatment exhibited availability of soil moisture that varied with the amount and distribution of rainfall, drought periods were found with soil water contents under PWP, the first at the end of the vegetative phase (50-60 das), then during the emergence of the panicle (80 to 90 das), and lastly, during anthesis and grain filling (110-140 das) (Figure 1B). The highest water deficit was observed in the 0-20 cm stratum, where more than 50 % of the root mass is concentrated (López-Castañeda and Richards, 1994).
Grain yield, yield components and root density under irrigation conditions
Under irrigation conditions, the variability between genotypes in terms of GY and its components was broad. GY, BM, HI, P M-2, G M-2, TGW and GP varied from 800 to 1606, from 1015 to 1425, from 43 to 53, from 125 to 257, from 15422 to 45538, from 28 to 37 and from 123 to 187, respectively; the widest variation was observed in grain yield and number of grains m-2 (Table 2). Lines 2 and 7 produced the highest GY accompanied by the highest values of BM, HI, G M-2, P M-2, TGW and GP (Table 2). The grain yield and its components in the control were lower than those found for the inbred lines. The high grain yield of lines 2 and 7 confirmed the outstanding behavior shown by the preselection cycles (Table 1). The grain yield of lines 2 and 7 was 44 % higher than the yield obtained with the two best experimental rice lines in average from three environments (Nayarit, Michoacán and San Luis Potosí) in Mexico (Barrios et al., 2016). The differences in grain yield could be due to the higher genetic potential for yield and resistance to diseases in the germplasm used here, than the one studied by Barrios et al. (2016); as well as the high ability of tillering, elongated large grain, and stability of the yield in all the environments (Salcedo and Barrios, 2013).
Table 2 Grain yield and yield components, and root density in different strata of the soil under irrigation conditions. 2013 spring-summer cycle. Zacatepec, Morelos, Mexico.
| No. de genotipo |
RG¶ (g m-2) |
BM¶ (g m-2) |
IC¶ (%) |
P M-2¶ | G M-2¶ | PMG¶ (g) |
GP¶ | Densidad de raíces (cm cm-3) | |||
| 0-15 cm | 15-30 cm | 30-50 cm | 50-70 cm | ||||||||
| 2 | 1606a | 1425ab | 51a | 257a | 45538a | 37a | 177b | 0.28a | 0.17bc | 0.09b | 0.09a |
| 7 | 1590a | 1425ab | 53a | 227b | 42458a | 33b | 187a | 0.22c | 0.19ab | 0.10ab | 0.05b |
| 3 | 1364b | 1165b | 52a | 143c | 18902de | 31bc | 132de | 0.29a | 0.16bc | 0.12a | 0.04b |
| 6 | 1330b | 1250bc | 51a | 143c | 22061cd | 32b | 155c | 0.29a | 0.15bc | 0.09ab | 0.05b |
| 5 | 1300b | 1254bc | 51a | 142c | 22682c | 31bc | 160c | 0.23bc | 0.22a | 0.08b | 0.04b |
| 4 | 1207c | 1172cd | 51a | 130c | 16871ef | 29cd | 130def | 0.27ab | 0.14c | 0.09ab | 0.06b |
| 1 | 972d | 1015d | 49b | 125c | 15422f | 28d | 123f | 0.28a | 0.18ab | 0.09ab | 0.05b |
| 8 | 890e | 1119cd | 45b | 230b | 31864b | 28d | 139d | 0.25abc | 0.16bc | 0.10ab | 0.04b |
| 9 (Testigo) | 800f | 1192cd | 43b | 131c | 16503ef | 28d | 126ef | 0.28a | 0.15bc | 0.11a | 0.05b |
| Media general | 1229 | 1247 | 50 | 170 | 25811 | 31 | 148 | 0.26 | 0.16 | 0.10 | 0.05 |
Means with different letter in a line are statistically different (Tukey, p ≤ 0.05). ¶See Materials and Methods.
The range of genetic variation for root density was 0.22 to 0.29, 0.15 to 0.22. 0.08 to 0.12 and 0.4 to 0.9 cm cm-3, in the strata: 0-15, 15-30, 30-50 and 50-70 cm, respectively; lines 2, 3, 6, 4, 1 and 9 (control); 7, 5 and 1; 7, 3, 6, 1, 8 and 9 (control); and line 2 showed the highest root density in the strata of 0-15, 15-30, 30-50 and 50-70 cm, respectively. The control line only had high root density in the strata of 0-15 and 30-50 cm (Table 3). In addition, it was determined that line 2 had higher root density in the strata from 0 down to 70 cm, which could be advantageous for a higher capture of water and soil nutrients. Ahmadi et al. (2014) indicated that the root system has a crucial function in the extraction of water and nutrients through the complex formed with the biogeochemical properties of the soil and in maintaining these functions under a broad range of stress scenarios, to ensure the survival of the plant and its reproduction. This function is even more important due to the growing instability of the weather and the limitations in the use of fertilizers and irrigation in the cultivation systems.
Table 3 Grain yield and yield components, and root density in different strata of the soil under rainfed conditions. 2013 spring-summer cycle. Zacatepec, Morelos, Mexico.
| No. de genotipo |
RG¶ (g m-2) |
BM¶ (g m-2) |
IC¶ (%) |
P M-2¶ | G M-2¶ | PMG¶ (g) |
GP¶ | Densidad de raíces (cm cm-3) | |||
| 0-15 cm | 15-30 cm | 30-50 cm | 50-70 cm | ||||||||
| 2 | 939a | 1433a | 40ab | 129a | 15045a | 26a | 117a | 0.35d | 0.33a | 0.21a | 0.13a |
| 7 | 980a | 1369a | 42a | 128a | 15131a | 26a | 118a | 0.53abc | 0.26bc | 0.18bc | 0.08d |
| 3 | 698cb | 1165b | 37ab | 121ab | 12359b | 21b | 102bc | 0.37bcd | 0.22c | 0.17c | 0.09bcd |
| 6 | 743b | 1162b | 39ab | 110c | 11515bc | 23b | 104abc | 0.38bcd | 0.25c | 0.18bc | 0.07d |
| 5 | 627cd | 1097bc | 37b | 113bc | 12557b | 21b | 111ab | 0.36cd | 0.26c | 0.20ab | 0.08d |
| 4 | 621cd | 991cd | 38ab | 116bc | 12500b | 21b | 108ab | 0.54ab | 0.24c | 0.18bc | 0.11abc |
| 1 | 385e | 924d | 30c | 92d | 8290d | 21b | 90c | 0.53abc | 0.30ab | 0.17c | 0.09bcd |
| 8 | 554d | 1030bc | 35b | 100d | 9900cd | 22b | 99bc | 0.57a | 0.22c | 0.18bc | 0.09cd |
| 9 (Testigo) | 203f | 937d | 18d | 95d | 11101bc | 21b | 118a | 0.47abcd | 0.23c | 0.17c | 0.13a |
| Media general | 638 | 1123 | 35 | 112 | 12044 | 22 | 107 | 0.45 | 0.26 | 0.18 | 0.10 |
Means with different letter in a line are statistically different (Tukey, p≤0.05). ¶See Materials and Methods.
Grain yield, yield components and root density under rainfed conditions
Under rainfed conditions, the grain yield was severely affected by the water deficit, and the average was 638 g m-2 vs. 1229 g m-2 under irrigation. The variability range in GY, BM, HI, P M-2, G M-2, TGW and GP was 203 to 980, 924 to 1433, 30 to 42, 92 to 129, 8290 to 15131, 21 to 26 and 90 to 118, respectively. BM and HI showed a broader range of variation in drought (BM=924 to 1433 and HI=30 to 42), than under irrigation (BM=1015 to 1425 and HI=43 to 53); the GY and G M-2 were the components with the highest genetic variation. Lines 2 and 7 produced the highest GY, BM, HI and G M-2; additionally, line 2 showed a high number of P M-2 and TGW, and line 7 showed a high number of GP (Table 3). The average HI of lines 3, 6, 5 and 4 was similar to that of lines 2 and 7. GY and all its components in the control were lower than the experimental lines. The higher yield of lines 2 and 7 both under irrigation and in drought confirmed the outstanding behavior shown by them in the preselection cycles (Tables 1 and 2). Our results are similar to those of Xangsayasane et al. (2014), who studied behavior of 227 experimental rice lines in three treatments: terminal drought, intermittent drought and irrigation, and found yield reduction of 34 and 59 % in the terminal drought treatments and intermittent drought with regards to irrigation. The reduction in GY was associated with a decrease in the percentage of filled grains, HI and P M-2.
The root density varied from 0.35 to 0.57, 0.22 to 0.33, 0.17 to 0.21 and 0.07 to 0.13 cm cm-3 in the strata of 0-15, 15-30, 30-50 and 50-70 cm, respectively; the lines 4 and 8; 2 and 1; 2 and 5; and 2 and control had a higher root density in the strata of 0-15, 15-30, 30-50 and 50-70 cm, respectively. Control showed high root density only in the stratum of 50-70 cm (Table 3). The roots have an important function in maintaining the water balance of the plant. Under conditions of moisture deficit, the water imbalance comes about when the rate of evaporation of the plant is higher than the rate of absorption of the roots (Boyer and Silk, 2004); under these conditions, higher dry weight of the root, root length, root percentage, number of nodal roots, specific root weight, root/shoot ratio, and capacity of osmotic adjustment are associated, individually or jointly, with the resistance of rice to drought (Fageria and Moreira, 2011; Feng et al., 2012; Gowda et al., 2011; Henry et al., 2016).
Comparison of irrigation vs. rainfed
The water deficiencies of the soil affected differentially the germplasm based on the GY and its components, and the RD in comparison with conditions of irrigation. The water deficit under rainfed conditions reduced the yield and its components and promoted a higher development of the root system (Table 4). The reduction due to water stress was more severe in GY (48 %) and G M-2 (53 %) than in the other yield components, while P M-2, HI, TGW and GP decreased 34, 30, 29 and 28 %, respectively. The BM was the least affected characteristic by drought (10 %) (Table 4). In 20 lines of rice selected for their high BM and subjected to flooding and rainfed conditions in the field, most of the genotypes had good behavior under conditions of flooding and only some behaved well under conditions of water stress. The germplasm susceptible to water stress showed a significant reduction in plant height, number of tillers per plant, and total aerial biomass at physiological maturity, although it was more delayed at flowering (Kondhia et al., 2015).
Table 4 Grain yield and yield components, and root density in different strata of the soil for all genotypes in irrigation and drought conditions. 2013 spring-summer cycle. Zacatepec, Morelos, Mexico.
| Tratamiento de humedad |
RG¶ | BM¶ | IC¶ | P M-2¶ | G M-2¶ | PMG¶ | GP¶ | Densidad de raíces (cm cm-3) | |||
| 0-15 cm |
15-30 cm |
30-50 cm |
50-70 cm |
||||||||
| Riego | 1229a | 1247a | 50a | 170a | 25811a | 31ª | 148a | 0.26b | 0.16b | 0.10b | 0.05b |
| Sequía | 638b | 1123b | 35b | 112b | 12044b | 22b | 107b | 0.45a | 0.26a | 0.18a | 0.10a |
Means with different letter in a row are statistically different (Tukey, p≤0.05). ¶See Materials and Methods.
In contrast with observations in grain yield and its components, the water deficit increased root density as compared to irrigation in every strata of the soil studied. The highest concentration of roots in average took place in the stratum of 0-30 cm both under irrigation (74 %) and drought (72 %) (Table 4). Kondo et al. (2003) studied the development of roots in the flowering stage in eleven genotypes of rice in the Philippines; they found that the effect of the genotype explained the higher proportion of variation in the number of nodal roots, specific root weight and root/shoot ratio, while the environmental effect was relatively large on the depth of root penetration and total dry weight of roots. In rice, the growth of the root system is influenced by the level of nutrition and moisture in the soil. In the province of Jiangsu, China, the Wuyunging 24 variety, subjected to three rates of nitrogen (100, 200 and 300 kg ha-1) and three soil moisture regimes (alternate irrigation with moderate drought, alternate irrigation with severe drought and continuous flooding), exhibited higher root dry weight in the treatments of continuous flooding and alternate irrigation with moderate drought with 200 and 300 kg N ha-1, as compared to alternate irrigation treatment with severe drought and 100 kg N ha-1 in the emergence of the panicle. On the contrary, the root/shoot ratio in the alternate irrigation treatment with severe drought and 100 kg N ha-1 was higher than in the other treatments, indicating that root growth in relation with the aerial parts is less inhibited in conditions of irrigation alternating with severe drought (Wang et al., 2016).
Interaction of varieties x levels of soil moisture
The interaction of varieties x levels of soil moisture was analyzed contrasting the GY and its components observed under irrigation with those under rainfed conditions. The GY and G M-2 showed the highest differences in irrigation - rainfed in average of all the genotypes, followed by the BM, P M-2, GP, HI and TGW (Table 5). Lines 2, 7, 3, 6, 5, 4, 1 and the control; the control; line 2; lines 2 and 7; line 2; line 7; and line 5, showed a greater difference irrigation - rainfed for GY; BM and HI; P M-2; G M-2; TGW; and GP, respectively (Table 5). When the water deficiency occurs during the vegetative phase of rice, its effect on the subsequent development and in the GY is minimal (30 %), although when it occurs during the development of the panicle, its effect on the GY is significant (70 %), delaying the anthesis (22-28 d) and reducing the number of spikelets per panicle (60 %) and the number of grains m-2 (100 %) or when it occurs during grain filling, its effect is also significant in the GY (50 %), delaying the physiological maturity (10-20 d) and decreasing the number of grains (40 %) and the individual weight of the grain (20 %) (Boonjung and Fukai, 1996). In low lands of southeastern Asia, intermittent and terminal drought in the rainy season reduced 13 and 35 % rice grain yield, while in the dry season yield decreased 34 % under intermittent drought and 59 % with terminal drought. These results allowed concluding that the selection of genotypes tolerant to drought could be achieved by alternating the evaluation of the germplasm under conditions of intermittent drought in the dry season, and with terminal drought in the rainy season (Xangsayasane et al., 2014).
Table 5 Irrigation - rainfed differences for grain yield and yield components, and root density in different strata of the soil. 2013 spring-summer cycle. Zacatepec, Morelos, Mexico.
| No. de genotipo |
RG¶ | BM¶ | IC¶ | P M-2¶ | G M-2¶ | PMG¶ | GP¶ | Densidad de raíces (cm cm-3) | |||
| 0-15 cm | 15-30 cm | 30-50 cm | 50-70 cm | ||||||||
| 2 | 667a | 109b | 11cd | 128a | 30493ª | 11.3a | 61ab | 0.07c | 0.16a | 0.13a | 0.05bc |
| 7 | 610a | 56b | 11cd | 99b | 27327ª | 6.7cd | 69a | 0.31a | 0.07cd | 0.08c | 0.03bc |
| 3 | 666a | 92b | 15bc | 22cd | 6544e | 10.0abc | 30ef | 0.08c | 0.05d | 0.06c | 0.05bc |
| 6 | 586a | 88b | 13cd | 32c | 10547c | 9.3abcd | 50bc | 0.09c | 0.10bc | 0.08bc | 0.03c |
| 5 | 673a | 156ab | 14cd | 29cd | 10126cd | 10.6ab | 49bcd | 0.13bc | 0.03d | 0.11ab | 0.04bc |
| 4 | 587a | 181 ab | 12cd | 14d | 4371e | 8.0abcd | 22fg | 0.27ab | 0.09bcd | 0.09abc | 0.05ab |
| 1 | 587a | 91b | 19ab | 33c | 7132de | 7.3bcd | 33def | 0.25abc | 0.12ab | 0.07c | 0.05bc |
| 8 | 336b | 89b | 10d | 130a | 21964b | 6.0d | 39cde | 0.32a | 0.06cd | 0.08bc | 0.06ab |
| 9 (Testigo) | 596a | 255a | 22a | 36c | 5402e | 7.7bcd | 8.3 g | 0.19abc | 0.07cd | 0.06c | 0.08a |
| Media general | 589 | 124 | 14 | 58 | 13767 | 8.5 | 40 | 0.18 | 0.085 | 0.083 | 0.048 |
Means with different letter in a row are statistically different (Tukey, p ≤ 0.05). ¶See Materials and Methods.
In average, for all the genotypes, the root density showed greater differences irrigation - rainfed in the stratum of 0-15 cm, as compared to the others (Table 5). Line 8, line 2 and the control, showed the highest root density in the stratum of 0-15, strata of 15-30 and 30-50 and 50-70 cm in comparison to the remaining germplasm, respectively (Table 5). The highest biomass accumulation in the roots can contribute to the higher grain yield and efficiency in using water in the agricultural rainfed systems. The larger size of the root system of the plant depends on higher root densities, total absorption surface area, and active root absorption surface area (Chu et al., 2014). A group of lines derived from the crosses between O. sativa (IR64) and O. glaberrima (RAM90 and RAM54) showed higher grain yield under irrigation and drought, as compared to the recurrent parent IR64 (susceptible to drought) and lower canopy temperature under drought; in addition, the root density was negatively associated to canopy temperature and soil moisture content (Kijoji et al., 2014). Given that the relationship between root density and grain yield in the field was also consistent in some lines, the complex relation between the grain yield, the root growth and the physiological mechanisms that control the growth of the crop and the productivity under field conditions is confirmed.
Grain yield vs. yield components and root density
The principal components analysis (PC) of grain yield versus yield components and root density, obtained for irrigation and rainfed conditions showed that the two first PC explained 83.6 % of the joint variation of the data. The first PC had positive correlation with TGW (R = 0.96) and GY (Y = 0.93), while the second PC showed positive correlation with BM (R = 0.64). The BM had a positive relationship with G M-2, P M-2 and GP, GY and TGW, and negative with HI; the HI showed negative relation with RD, particularly at 0-15 cm of soil depth (Figure 2). The positive association between the GY and its components, BM and G M-2 can be useful for the selection in contrasting environments in terms of the availability of moisture. The most outstanding rice lines in three environments of Mexico, in addition to high yield, showed high percentages of recuperation of full grains and good quality for industrial use of rice (Barrios et al., 2016). In barley cultivated under irrigation, drought and rainfed conditions in Mexico, the grain yield had a positive and significant relationship with the aerial biomass and the number of grains m-2. Thus, the aerial biomass, in its turn, was associated positively and significantly with the number of grains m-2, so that the genotypes with highest grain yield showed higher aerial biomass and higher number of grains m-2 (López-Castañeda, 2011).
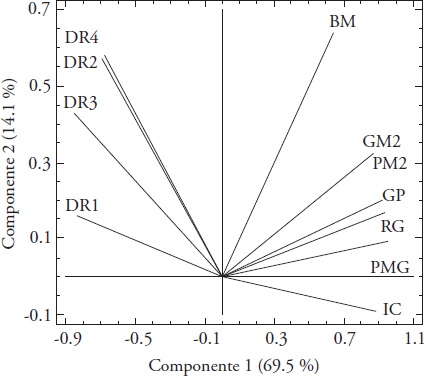
Figure 2 Biplot of the grain yield versus yield components and root density for nine genotypes of rice in maturity; BM, aerial biomass; GM2, number of grains m-2; PM2, number of panicles m-2; GP, number of grains per panicle; GY, grain yield; TGW, thousand grains weight; HI, harvest index; DR1, root density in the stratum of 0-15 cm; DR2, root density in the stratum of 15-30 cm; DR3, root density in the stratum of 30-50 cm; DR4, root density in the stratum of 50-70 cm soil depth.
Conclusions
The soil water deficit during flowering and grain filling decreased grain yield and its components in the rice germplasm cultivated under rainfed conditions in Zacatepec, Morelos, Mexico, while this deficiency of soil moisture increased the root density in all the strata of the soil studied. Lines 2 and 7 showed the highest grain yields under irrigation and rainfed conditions, as well as values higher than the national average in irrigation and rainfed conditions. The aerial biomass, harvest index, number of panicles m-2, number of grains m-2 and number of grains per panicle, as well as the thousand grains weight were significantly associated with the grain yield in both soil moisture conditions. The most productive lines could be used as parents in a rice breeding program and as commercial varieties in the rice production systems, both under irrigation and rainfed conditions.
Literatura Citada
Ahmadi, N., A. Audebert, M. J. Bennett, A. Bishop, A. Costa de Oliveira, B. Curtois, A. Diedhiou, A. Diévart, P. Gantet, A. Ghesquièrè, E. Guiderdoni, A. Henry, Y. Inukai, L. Kochian, L. Laplace, M. Lucas, D. T. Luu, B. Manneh, X. Mo, R. Muthurajan, C. Périn, A. Price, S. Robin, H. Sentenac, B. Sine, Y. Uga, A. A. Véry, M. Wissuwa, P. Wu and J. Xu. 2014. The roots of future rice harvests. Rice 7: 1-9. [ Links ]
Barrios G., E. J., V. H. Rodríguez M., L. Hernández A., L. Tavitas F., A. Hernández P., L. M. Tapia V., y J. M. Pinzón G. 2016. Evaluación de líneas de arroz de grano delgado para riego en México. Interciencia 41: 476-481. [ Links ]
Boonjung, H., and S. Fukai. 1996. Effects of soil water deficit at different growth stages on rice growth and yield under upland conditions. 2. Phenology, biomass and yield. Field Crops Res. 48: 47-55. [ Links ]
Boyer, J. S., and W. K. Silk. 2004. Hydraulics of plant growth. Functional Plant Biol. 31:761-773. [ Links ]
Chatel, M., E. P. Guimarães, Y. Ospina, F. Rodríguez, y V. H. Lozano. 2010. Mejoramiento de poblaciones de arroz de secano empleando selección recurrente y desarrollo de variedades. In: Degiovanni, V. B., C. P. Martínez R., y F. O. Motta (eds). Producción Eco-eficiente del Arroz en América Latina. Centro Internacional de Agricultura Tropical (CIAT), Publicación CIAT No. 370. Cali, Colombia. pp: 207-222. [ Links ]
Cho, J., and T. Oki. 2012. Application of temperature, water stress, CO2 in rice growth models. Rice 5: 1-8. [ Links ]
Chu, G., T. Chen, Z. Wang, J. Yang, and J. Zhang. 2014. Morphological and physiological traits of roots and their relationships with water productivity in water-saving and drought-resistant rice. Field Crops Res. 165: 36-48. [ Links ]
Fageria, N. K., and A. Moreira. 2011. The role of mineral nutrition on root crop growth of crop plants. Adv. Agron. 110: 251-331. [ Links ]
Feng, F., X. Xu, X. Du, H. Tong, L. Luo, and H. Mei. 2012. Assessment of drought resistance among wild rice accessions using a protocol based on single-tiller propagation and PVC-tube cultivation. Aust. J. Crop Sci. 6: 1204-1211. [ Links ]
García A., J. L., L. Hernández A., y L. Tavitas F. 2011. El Silverio: Nueva variedad de arroz para el trópico mexicano. Rev. Mex. Ciencias Agr. 2: 607-612. [ Links ]
García, E. 2004. Modificaciones al Sistema de Clasificación Climática de Köppen. Quinta edición. Instituto de Geografía, UNAM. México, D.F. 90 p. [ Links ]
Gowda, V. R. P., A. Henry, A. Yamauchi, H. E. Shashidhar, and R. Serraj. 2011. Root biology and genetic improvement for drought avoidance in rice. Field Crops Res. 122: 1-13. [ Links ]
Henry, A., R. Wehler, A. Grondin, R. Franke, and M. Quintana. 2016. Environmental and physiological effects on grouping of drought-tolerant and susceptible rice varieties related to rice (Oryza sativa) root hydraulics under drought. Ann. Bot. 118: 711-724. [ Links ]
Jagadish, S.V. K., M.V.R. Murty, and W.P. Quick. 2015. Rice responses to rising temperatures - challenges, perspectives and future directions. Plant, Cell Env. 38: 1686-1698. [ Links ]
Kamoshita, A., R. C. Babu, N. M. Boopathi, and S. Fukai. 2008. Phenotypic and genotypic analysis of drought-resistance traits for development of rice cultivars adapted to rainfed environments. Field Crops Res. 109 :1-23. [ Links ]
Kijoji, A. A., S. Nchimbi-Msolla, Z. L. Kanyeka, R. Serraj, and A. Henry. 2014. Linking root traits and grain yield for rainfed rice in sub-Saharan Africa: Response of Oryza sativa x Oryza glaberrima introgression lines under drought. Field Crops Res. 165: 25-35. [ Links ]
Kondhia, A., R. E. Tabien, and A. Ibrahim. 2015. Evaluation and selection of high biomass rice (Oryza sativa L.) for drought tolerance. Am. J. Plant. Sci. 6: 1962-1972. [ Links ]
Kondo, M., P. P. Pablico, D. V. Aragones, R. Agbisit, J. Abe, S. Morita, and B. Courtois. 2003. Genotypic and environmental variations in root morphology in rice genotypes under upland field conditions. Plant and Soil 255: 189-200. [ Links ]
Kumar, A., S. Dixit, T. Ram, R. B. Yadaw, K. K. Mishra, and N. P. Mandal. 2014. Breeding high-yielding drought-tolerant rice: genetic variations and conventional and molecular approaches. J. Exp. Bot. 65: 6265-6278. [ Links ]
López-Castañeda, C., and R. A. Richards. 1994. Variation in temperate cereals in rainfed environments. I. Grain yield, biomass and agronomic characteristics. Field Crop Res. 37: 51-62. [ Links ]
López-Castañeda, C. 2011. Variación en rendimiento de grano, biomasa y número de granos en cebada bajo tres condiciones de humedad del suelo. Trop. Subtrop. Agroecosystems 14: 907-918. [ Links ]
Lynch, J. P., J. G. Chimungu, and K. M. Brown. 2014. Root anatomical phenes associated with water acquisition from drying soil: targets for crop improvement. J. Exp. Bot. 65: 6155-6166. [ Links ]
Miranda-Domínguez, L. E., C. López-Castañeda, I. Benítez-Riquelme y J. A. Mejía-Contreras. 2016. Desarrollo radical y rendimiento en diferentes variadeades de trigo, cebada y triticale bajo condiciones limitantes de humedad en el suelo. Terra Latinoam. 34: 393-407. [ Links ]
Osuna C, F. J., L. Hernández A., J. Salcedo A., L. Tavitas F., y L. J. Gutiérrez D. 2000. Manual para la Producción de Arroz en la Región Central de México. Libro Técnico No. 1. Campo Experimental Zacatepec, Centro de Investigación Regional del Centro, INIFAP, SAGAR. Zacatepec, Morelos, México. 92 p. [ Links ]
Pandey, V., and A. Shukla. 2015. Acclimation and tolerance strategies of rice under drought stress. Rice Sci. 22: 147-161. [ Links ]
Passioura, J. B., and J. F. Angus. 2010. Improving productivity of crops in water-limited environments. Adv. Agron. 106: 37-75. [ Links ]
SAGARPA. 2017. Servicio de Información y Estadística Agroalimentaria y Pesquera. Anuario Estadístico de la Producción Agrícola. http://www.sagarpa.gob.mx (Consulta: marzo 2018). [ Links ]
Salcedo A., J., y E. J. Barrios G. 2013. Morelos A-08: cultivar de arroz de grano aromático para el estado de Morelos. Rev. Mex. Ciencias Agr. 4: 963-968. [ Links ]
Sánchez, B., A. Rasmussen, and J. R. Porter. 2014. Temperatures and the growth and development of maize and rice: a review. Global Change Biol. 20: 408-417. [ Links ]
SAS. 2009. The SAS System Program release 9.1 for Windows. SAS Institute, Inc., Cary, North Carolina, U.S.A. Software of statistical analysis. [ Links ]
Shah, F., J. Huang, K. Cui, L. Nie, T. Shah, C. Chen, and K. Wang. 2011. Impact of high-temperature stress on rice plant and its traits related to tolerance. J. Agric. Sci. Cambridge 149: 545-556. [ Links ]
STATGRAPHICS. 2010. Centurion XVI. Versión 16.1.11. Statpoint Technologies Inc. Software of statistical analysis and graphics (www.STATGRAPHICS.com). [ Links ]
Tennant, D. M. 1975. A test of a modified line intersect method of estimating root length. J. Ecol. 63: 995-1001. [ Links ]
Wang, Z., W. Zhang, S. S. Beebout, H. Zhang, L. Liu, J. Yang, and J. Zhang. 2016. Grain yield, water and nitrogen use efficiencies of rice as influenced by irrigation regimes and their interaction with nitrogen rates. Field Crops Res. 193: 54-69. [ Links ]
Xangsayasane, P., B. Jongdee, G. Pantuwan, S. Fukai, J. H. Mitchell, P. Inthapanya, and D. Jothiyangkoon. 2014. Genotypic performance under intermittent and terminal drought screening in rainfed lowland rice. Field Crops Res. 156: 281-292. [ Links ]
Received: October 2016; Accepted: March 2018
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
