











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Wheat (Triticum spp.) is the most cultivated cereal for human consumption worldwide and despite its growing production, grain yield losses caused mainly by rust diseases can reach 30 % to 60 % and up to total loss in extreme situations (Roelfs et al., 1992; Curtis, 2002). In order to deal with the current world challenges for food provision, it was highlighted the need of more efficient and integrated cereal farming systems which may include strategies to decrease the development of pest populations without threatening the environment and human health (FAO, 2014). An earlier and pronounced induction of intrinsic defence mechanisms in resistant plant varieties was observed upon pathogen attack (Moloi and van der Westhuizen, 2006). This has encouraged the utilization of biotic and abiotic stressors to enhance defence responses in vulnerable plants, which may contribute to avoid or minimize crop losses. Several physiological and biochemical responses indicate that some elicitors improve the systemic acquired resistance of plants through the stimulation of synthesis of defence related metabolites and enzymes (Gozzo, 2003). In wheat, aphid infestation raised the levels of H2O2 and NADPH oxidase activity (Moloi and van der Westhuizen, 2006). An early systemic induction of expression levels of peroxidase and pathogenesis-related (PR)-genes was observed after fungal infection in wheat spikes (Pritsch et al., 2001), while infiltration of detached young wheat leaves with Fusarium mycotoxin triggered accumulation of ROS and defence related transcripts and proteins (Desmond et al., 2008). Lignification in wounded and intact wheat leaves (Barber et al., 1989) as well as induction of the activity of the defence-related enzymes phenylalanine ammonia-lyase and peroxidase (Vander et al., 1998) were promoted by chitosan; a β-(1-4) linked (acetylated or deacetylated) D-glucosamine polysaccharide. The elicitor properties of chitosan were studied for decades in different plants, and are dependent of physicochemical characteristics such as the degree of polymerization and acetylation. The chemical nature and length of chitosan oligosaccharides can differentially influence the transcription, accumulation and activity of PR proteins in rice (Lin et al., 2005) and peach (Ma et al., 2013), and increase the activity of enzymes and the synthesis of defensive metabolites in wheat (Vander et al., 1998) and tobacco (Falcón-Rodríguez et al., 2009). According to Li et al. (2016), fungal-derived oligochitosans are able to exert a direct fungitoxic activity against phytopathogens and simultaneously elicit the activity of defensive enzymes and accumulation of phenolic compounds in the medicinal plant Z. bungeanum. However, the influence of chitosan derivatives on the transcription patterns of defence-related genes in wheat has not been studied so far. Therefore, the hypothesis of our study was that the different structural features of chitosan derivatives might influence the transcriptional patterns related to protective responses in wheat. The aim of our study was to evaluate the induction of local and systemic accumulation of defence-related transcripts in wheat seedlings by chitosans with different physicochemical characteristics.
Materials and Methods
Preparation and characterization of chitosan and chitosan derivatives
Commercially available chitosan (Sigma-Aldrich, USA) was dissolved overnight in 1 % acetic acid, pH 5.5. The chitosan average molecular weight (M) was determined by capillary viscosimetry (Cabrera and Van Cutsem, 2005) using an Ubbelohde viscometer and the Mark-Houwink’s equation [η]=KMα , where [η]=intrinsic viscosity, K=0.076 and α=0.76. The degree of acetylation (DA) was determined by infrared spectroscopy using the equation DA %=31.92 A1320/A1420-12.2 according to Brugnerotto et al. (2001). For the production of chitooligomers a 10 g L-1 chitosan solution was hydrolysed with papain (Sigma-Aldrich, USA) at 35 °C for 24 h as suggested by Vishu et al. (2004). The chitooligosaccharides (COS) were characterized by matrix assisted laser desorption/ionization-time of flight-mass spectrometry (MALDI-TOF-MS) at the Complex Carbohydrate Research Center, University of Georgia. Briefly, a 5 mg mL-1 COS solution in deionized water was sonicated for 1 h to dissolve the soluble material, filtered through 0.22-μm Durapore membrane filters (Millipore, Ireland) and mixed with a 2,5-dihydroxybenzoic acid matrix (Cabrera and Van Cutsem, 2005). Mass spectra were recorded on an Applied Biosystems 5800 MALDI-TOF in the positive ion mode.
Plant material and treatments
Wheat (Triticum aestivum cv. Tacupeto F2001) seedlings were grown in a controlled environment chamber, with a 16 h photoperiod and 28/24 °C day/night temperature. Two experiments were conducted according to the type of chitosan derivative evaluated (polymeric chitosan or COS). For each experiment, 18 pots with 25-day-old seedlings were randomly assigned to six different groups, where chitosan or COS at 1 g L-1 and 0.1 g L-1 were either sprayed to leaves or applied to the substrate of growing plants. Such concentrations were chosen according to Falcón-Rodríguez et al. (2009; 2011). Control groups were treated with dd-water. Samples of leaves from four to five seedlings were collected from each pot, pooled together and considered a replication. Subsequently, leaves were frozen and ground with liquid nitrogen at 0, 6 and 12 h after treatments for further expression analysis of the defence-related genes chitinase (CHI), β-1-3 glucanase (GLU) and phenylalanine ammonia-lyase (PAL).
RNA extraction and reverse transcription-polymerase chain reaction (RT-PCR)
Total RNA was extracted with the Concert Plant Reagent (Invitrogen, USA) according to the manufacturer’s instructions and genomic DNA was removed with the DNA-free kit (Ambion, USA). One microgram of total RNA was reverse transcribed using the SuperScript II Reverse Transcriptase (Invitrogen, USA) and oligodT priming as recommended by the supplier. Semiquantitative PCR was performed using the GoTaq® Flexi DNA Polymerase kit (Promega, USA) with serially diluted cDNA template in 28 cycles of 95 °C for 1 min, 58 °C for 1 min and 72 °C for 1 min. Control reactions to normalize RT-PCR amplification were run with primers for glyceraldehyde 3-phosphate dehydrogenase (GAPDH). The gene-specific primer pairs used for the PCR were designed from sequences in the GenBank database (Table 1). Amplification products were analysed by 1 % agarose gel electrophoresis stained with Gel-Red (Biotium, USA) and visualized under UV light with the Kodak Gel Logic 100 Imaging System. Band densities were quantified using the software 1D Image Analysis (Kodak, USA). The results were expressed as a ratio between the total band densitometric intensity of each analysed gene and the reference gene GAPDH.
Table 1 Primers used in semi-quantitative reverse transcription-polymerase chain reaction (RT-PCR) analysis of gene expression.
| Gene | Primer sequence | GenBank accession number |
| Chitinase (CHI) | Sense 5’- GTTCTACACGTACGACGCCTTCTT-3’ Antisense 5’- CCCGCCGTTGATGACGTTGGTGAT-3’ |
AB029935.1 |
| β-1,3-Glucanase (GLU) | Sense 5’-ATCGACCTCATCATGGACGTGG-3’ Antisense 5’-GTCGAAGAGGTTTGTGTAGGTGAG-3’ |
AB244639.2 |
| Phenylalanine ammonia-lyase (PAL) | Sense 5’-CCAATGTTCTGTCCGTCCTTGCT-3 ’ Antisense 5’-AACTGCGCGAACATCAGCTTGC-3’ |
AY005474.1 |
|
Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) |
Sense 5’-GATTTGGTCTCCACTGACTTTGTCG-3’ Antisense 5’-CAACTAACATGAACCAGGCGTCG-3’ |
AY290728.1 |
Results and Discussion
Characterization of chitosan and chitosan derivatives
Chitosan solution presented a deacetylation degree of 84.5 %, which was calculated by considering the intensity ratio of infra-red absorption bands of N-acetyl glucosamine and glucosamine. In addition, viscometry results showed that chitosan in solution had a molecular weight of 50.8 kDa, considered low for a polymeric chitosan. Hydrolysed chitosan derivatives were characterized and mass spectrometry analysis revealed that oligomeric species found in the COS mixture had three or four monomeric units (chitotrioses and a chitotetraose) with different degrees of acetylation (Figure 1). Four main ion species were detected as sodium or potassium adducts at m/z values of 580.6, 607, 663.6 and 783.1, where m/z 580.6 corresponds to a monoacetylated chitotriose, whereas fully acetylated COS were detected at m/z 663.6. Molecular weight and acetylation degree directly influence the biological activity of chitosan and its derivatives (Kauss et al., 1989; Falcón-Rodríguez et al., 2009). Highly active COS have a degree of polymerization from four to six units with acetyl groups (Barber et al., 1989), whereas fully deacetylated COS lack capacity to elicit defensive reactions in plants.
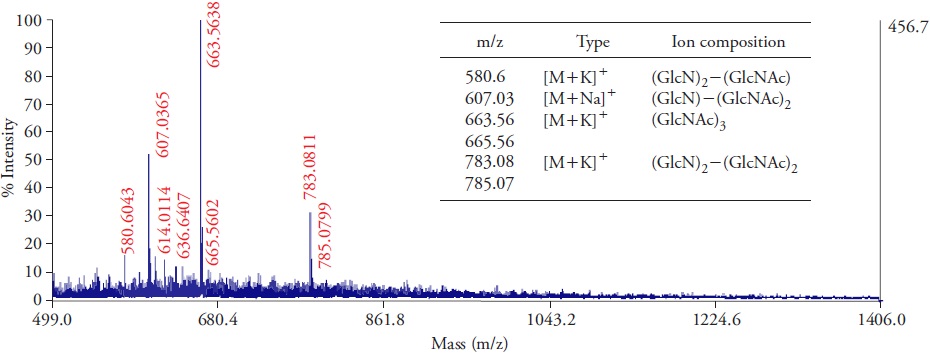
Chitosan exhibits inductive properties even when degree of acetylation varies from 1 % to 60 %, but with different intensity (Vander et al., 1998). Also, this characteristic is associated to selective modulation of enzymes PAL and peroxidase in tobacco plants (Falcón-Rodríguez et al., 2011). Interestingly, a linear deacetylated COS with a degree of polymerization of five reduced the incidence of Fusarium infection in Z. bungeanum and increased activity of enzymes related to protective mechanisms (Li et al., 2016). This may suggest certain specificity between the structural features in chitosan derivatives with respect to the studied plant species.
Expression of the genes chitinase (CHI), β-1-3 glucanase (GLU) and phenylalanine ammonia-lyase (PAL)
Chitosan and COS treatments induced both local and systemic responses in leaves of wheat seedlings, but with different speed and intensity depending on type of elicitor, concentration and gene evaluated (Figures 2 and 3). Expression of CHI and GLU was increased to different extents in seedlings treated with both types of elicitor, whereas PAL expression was differentially regulated: PAL transcript levels were increased by chitosan (Figure 2E, F) but decreased by COS (Figure 3E, F), with respect to controls.
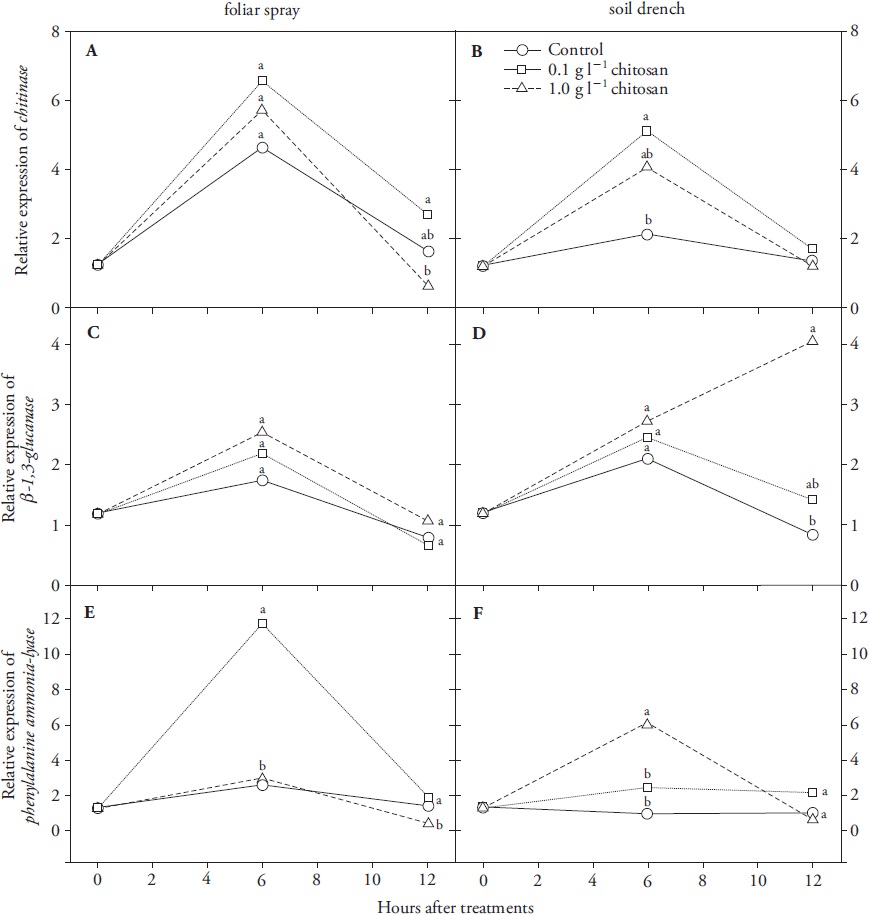
Mean values of three replications are presented. Means with different letters were significantly different (p≤0.05).
Figure 2 Changes in relative gene expression levels of defence-related markers chitinase (CHI) (A, B), β-1,3-glucanase (GLU) (C, D) and phenylalanine ammonia-lyase (PAL) (E, F) in leaves of chitosan-treated wheat seedlings by foliar spray or soil drench.
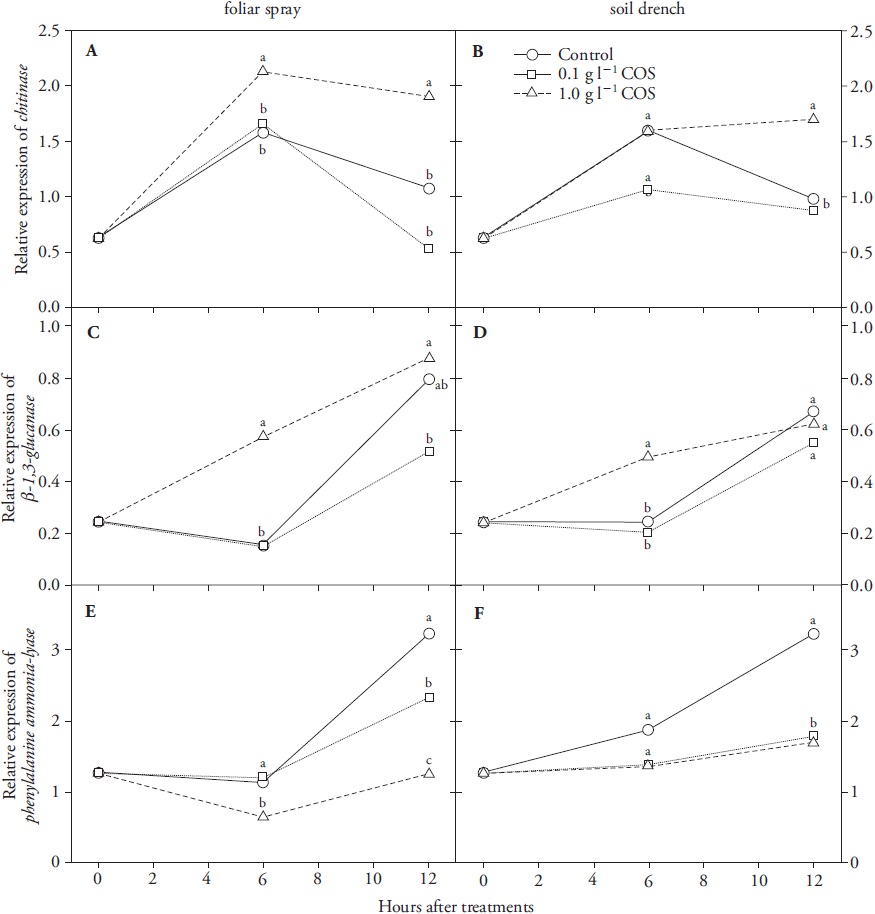
Mean values of three replications are presented. Means with different letters were statistically different (p≤0.05).
Figure 3 Changes in the relative gene expression levels of defence-related markers chitinase (CHI) (A, B), β-1,3-glucanase (GLU) (C, D) and phenylalanine ammonia-lyase (PAL) (E, F) in leaves of wheat seedlings treated with chitooligosaccharides (COS) by foliar spray or soil drench.
Chitosan treatments elicited distal expression of the three evaluated genes; CHI and PAL transcripts peaked at hour 6 and went back to normal after 12 h (Figure 2B, F). GLU transcripts increased 12 h after 1 g L-1 application (Figure 2D). Differences in expression among analyzed genes may rely on a different sensitivity to the stimulus, where a certain concentration may be sufficient for triggering a particular response. In our trial, the influence of elicitor concentration for transient PAL induction, within the first 6 h post-treatments, was evident (Figure 2 E and F): lower chitosan concentration (0.1 g L-1) increased PAL transcripts in directly sprayed leaves. Distal induction of PAL was only favoured by more concentrated solution (1 g L-1) applied to roots. This observation may show that perception of polymeric chitosan by radicular tissue was probably hampered by the soil matrix, thus influencing the availability of active molecules that trigger the accumulation of PAL transcripts. Increment of PAL activity can indirectly improve defensive performance of plant tissues by reinforcing physical barriers with lignin (Barros et al., 2015) and by enabling production of phytoalexins (Hasan et al., 2013).
A non-linear dose-response effect on accumulation of phenylpropanoid compounds and on increment of PAL, GLU and CHI enzymatic activities was reported for cultured parsley cells treated with fungal elicitors (Kombrink and Hahlbrock, 1986). Hence, chitosan dose-response curves would be necessary to test this effect on wheat seedlings. Foliar application of chitosan derivatives increased enzymatic activity of PAL and GLU in tobacco seedlings, and promoted systemic protection against the oomycete pathogen Phytophthora nicotianae (Falcón-Rodriguez et al., 2011). However, when chitosan was directly applied to wheat leaves no significant induction of CHI and GLU transcript levels was observed (Figure 2A, C), which would suggest different specificity among plant species. Lin et al. (2005) found that the polymeric chitosan was less effective than the smaller COS at raising H2O2 levels and at inducing expression of PR genes CHI and GLU in rice cells. Increment in H2O2 levels is one of the first cell signalling mechanisms for activation of defensive responses in plants.
One g L-1 COS was a more effective concentration than 0.1 g L-1 to enhance the expression levels of CHI and GLU in both foliar and root treated wheat seedlings (Figure 3). In foliar treated plants CHI transcript levels were higher than controls within the first 6 h post-treatment and this effect was maintained by hour 12 (Figure 3A). Subsequent experiments showed that the induction of CHI transcript levels in foliar treated wheat plants was evident earlier than hour 6 (data not shown). The expression of GLU was transiently enhanced after 6 h in leaves of both foliar and root treated seedlings with 1 g L-1 COS (Figure 3C, D), but controls reached levels of treated plants by hour 12. Our results are similar to those reported by Ma et al. (2013), who showed that GLU expression was induced in peach fruit at 12 and 24 h after dip treatment with 5 g L-1 chitosan or COS. In contrast, COS significantly reduced both local and distal PAL expression (Figure 3E, F). This may likely indicate a regulatory effect, where COS might differentially induce or repress some defence-related pathways for a limited period of time. Such dynamic behaviour in expression profiles between PAL and the PR genes CHI and GLU could optimize cell energy waste due to de novo protein synthesis from induced transcripts.
An enhanced GLU enzymatic activity was observed in tobacco seedlings treated with COS by Falcón-Rodriguez et al. (2011). The increment in the expression and activity of enzymes GLU and CHI was associated to an enhanced plant tolerance to fungal infections (Zhu et al., 1994). These enzymes hydrolyse components of cell wall of diverse phytopathogens and insect cuticle; therefore, they are key effectors of the systemic acquired resistance in plants, which is mainly triggered by phytohormone salicylic acid (Durner et al., 1997).
The relationship between the increments of salicylic acid content and the activity of GLU in chitosan-treated peanut leaves was reported by Sathiyabama and Balasubra-manian (1998). One of the known pathways for salicylic acid biosynthesis in plants is led by PAL (Chen et al., 2009). However, the decrease of PAL transcript levels observed in the COS-treated wheat seedlings, did not avoid the increment of the PR transcripts CHI and GLU, which may have occurred by a COS-induced salicylic acid accumulation predominantly through an alternative pathway controlled by the enzyme isochorismate synthase (Catinot et al., 2008). Therefore, it would be very interesting to study the effect of chitosan derivatives on the expression of isochorismate synthase, but also to determine if the induction of these defence-related transcripts correlates with an in vivo decreased susceptibility of chitosan- and COS-treated wheat seedlings to fungal infections.
The differences between the expression patterns elicited by chitosan and COS indicate that molecular species in the COS mixture may have a particular action mechanism different from that of polymeric chitosan, which is a more complex molecule with a different acetylation pattern. Cell recognition patterns for non-acetylated species seems to lack specific receptors, hence eliciting properties of chitosan appears to be mediated by the interaction between positively charges and negatively charged membrane phospholipids (Silipo et al., 2010). Some potential receptors that bind with high affinity to more acetylated species were characterized from plasma membrane of model plants (Kaku et al., 2006; Miya et al., 2007), suggesting that N-acetylated COS may trigger specific responses, which may in part explain the different expression patterns between COS- or chitosan-treated wheat seedlings. Our results agree with Vander et al. (1998), who found that partially N-acetylated chitosans elicited enzymatic activities of resistant markers in wheat leaves, whereas oligomeric species failed to induce lignin deposition. Biosynthesis of lignin precursors is carried out in the phenylpropanoid pathway, which is regulated by PAL. Therefore, it would be interesting to see whether or not a combined treatment of polymeric chitosan and COS could enhance the induction of defence responses in a greater extent.
Conclusion
The capacity to differentially modulate the local and systemic expression of defence-related genes in wheat by chitosan derivatives was shown by our research. Type and concentration of elicitor appear to be determinant factors for the response: COS with a polymerization degree from three to four units exert a stronger effect at higher doses. But, polymeric chitosan concentrations apparently do not follow a direct linear relationship with induction of defence-related transcripts accumulation. These results may help understand the activation of protective processes elicited by chitosan derivatives in wheat plantlets and its possible relation to induction of systemic resistance.

