











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La biomasa forestal permite caracterizar la capacidad acumulativa de materia orgánica en los ecosistemas en el tiempo (Eamus et al., 2000) y cuantificar los nutrimentos almacenados en los tejidos de la planta o del tipo de vegetación (Fonseca et al., 2009). Estas estimaciones se usan en estudios de eficiencia nutricional y evaluación de las funciones ambientales y servicios ecosistémicos de los bosques naturales (Ferrere et al., 2014) o plantaciones forestales.
La biomasa acumulada en un bosque o plantación es un indicador del crecimiento vegetal y del C fijado (Návar, 2009). Esta información es necesaria para valorar la contribución de las cubiertas vegetales en la reducción de gases con efecto invernadero (Fonseca et al., 2009) y en la planeación regional del manejo forestal sustentable (Kauffman et al., 2009; Cutini et al., 2013), ambos enfocados a reducir los impactos del cambio climático en el planeta (Malhi y Grace, 2000; Snowdon et al., 2001) planteados en el Protocolo de Kyoto (Naciones Unidas, 1998).
La biomasa en los ecosistemas forestales se divide en biomasa área (fuste, ramas y follaje) y biomasa subterránea (raíces), esta última es la más costosa y complicada de estudiar (Gárete y Blanco, 2013). La biomasa aérea de un árbol se define como la suma de la cantidad total de materia orgánica de hojas, ramas, tronco y corteza (Garzuglia y Saket, 2003) y puede calcularse con métodos directos e indirectos (Vásquez y Arellano, 2012). De ellos el primero es el más empleado (Díaz-Franco, 2007). Cuando la estimación corresponde a un bosque, la muestra debe considerar variables dasométricas, edad, calidad de sitio, composición de especies, y condiciones climáticas, edáficas y topográficas (Avendaño-Hernández et al., 2009), o el tipo de clones establecidos en las plantaciones forestales comerciales (PFC), para mejorar la estimación por cada condición especifica.
Los modelos alométricos son herramientas para estimar biomasa y C capturado en los bosques (Návar, 2010). Esas funciones usan la correlación entre variables que son difícil de medir, como volumen (V), C, C equivalente (CO 2e ), biomasa verde y biomasa seca de cada componente, y con variables fáciles de medir, como el diámetro normal (dn) y la altura total (AT) de los árboles (Solano et al., 2014). En combinación con datos de inventarios forestales y algunas funciones de crecimiento y rendimiento, las ecuaciones alométricas permiten cuantificar el potencial de crecimiento vegetal y fijación de gases con efecto invernadero, es decir el CO2 capturado durante la fotosíntesis; además, representan una alternativa de valoración de las áreas forestales para el pago de servicios ambientales por captura y secuestro de CO2 (Ruíz-Aquino et al., 2014).
El objetivo del estudio fue estimar la biomasa aérea total y por componente estructural a partir de modelos alométricos y factores de expansión de biomasa (FEB’s); además, evaluar la aplicabilidad en inventarios forestales para clones de Eucalyptus urophylla S. T. Blake de PFC de Tabasco, México. La hipótesis fue que a través de cualquiera de los métodos utilizados es posible estimar con precisión la biomasa aérea por árbol y por unidad de superficie.
Materiales y Métodos
Área de estudio
Las plantaciones evaluadas de E. urophylla, con labores culturales, sin intervención silvícola y densidad de 1100 plantas por ha están en el municipio de Huimanguillo, Tabasco, México (17° 55’ N, 94° 06’ O y altitud promedio de 30 m). En la región existen estaciones con lluvia y secas, con clima cálido húmedo (Am), temperatura media anual de 26 °C y precipitación de 2500 mm. Los suelos son de tipo Feozem con relieve de tipo lomerío (INEGI, 2005).
Selección y medición de la muestra
Para recolectar la información se realizó un muestreo completamente al azar en siete plantaciones dispersas de E. urophylla, en el cual se seleccionaron y destruyeron 93 árboles clónales, de uno a siete años de edad, elegidos por la condición morfológica superior dentro de las plantaciones y tratando de cubrir la variabilidad mayor de edades y crecimiento. En todos los árboles se midió el diámetro normal (dn) antes de derribarlos, y luego se midió su altura total (AT), diámetro y altura de tocón (dt y Atc), los diámetros basales (db) por rama y a la altura de inserción en el fuste (Adb). Todas las mediciones se registraron en metros.
Separación por componentes
Fuste con corteza, ramas y follaje se separaron y pesaron para obtener el peso “verde” de las muestras en campo (Pv, en kg) con la metodología propuesta por Avendaño-Hernández et al. (2009) y Gómez-Díaz et al. (2011). El fuste se seccionó en trozas de un metro de longitud, iniciando en la altura del tocón hasta la punta, cada sección se pesó en una báscula electrónica con plataforma (capacidad de 100 kg y precisión de 0.001 kg). También se obtuvieron tres rodajas de 5 cm de grosor correspondientes al tocón, parte media y punta del fuste de cada árbol. Las ramas y el follaje se separon y pesaron en una báscula electrónica (capacidad de 5 kg y precisión de 0.001 kg). Una muestra de ramas y follaje, de aproximadamente 0.5 kg, se seleccionó para determinar la relación entre el peso verde y peso seco. Este último se determinó en laboratorio. La metodología fue documentada por Ruíz-Aquino et al. (2014) y Soriano-Luna et al. (2015).
Estimación de volumen con corteza y biomasa aérea
El volumen de cada sección o troza (V troza ) se obtuvo con la fórmula de Newton y el volumen de punta (V punta ) con la fórmula del cono. El volumen total del árbol (Vt) se calculó con el método de trozas traslapadas propuesto por Bailey (1995). Las muestras de ramas y hojas se deshidrataron a 72 °C y las rodajas a 105 °C, hasta alcanzar un peso constante, en una estufa para secado. Los pesos se registraron con una precisión de 0.001 kg. Con el peso húmedo y seco se determinó el contenido de humedad por componente y la relación entre ellos. La biomasa área total se definió como la suma del peso seco de los tres componentes (Domínguez-Cabrera et al., 2009; Lim et al., 2013); además, se determinó el promedio de proporción de biomasa por componente con respecto a la biomasa total.
Análisis exploratorio de la muestra
Un análisis exploratorio de las variables independientes (dn y AT) y las dependientes (B f , B r , B h , B t y V) mostró la variabilidad amplia de las dimensiones de los individuos (Cuadro 1).
Cuadro 1 Valores de estadísticos básicos de las variables de los árboles muestreados en plantaciones forestales comerciales de Eucalyptus urophylla, con uno a siete años de edad en Huimanguillo, Tabasco, México.

Dn: diámetro normal, AT: altura total, Bf: biomasa de fuste, Br: biomasa de ramas, Bh: biomasa de follaje, Bt: es la biomasa total, V: volumen, SD: desviación estándar de la media.
Ajuste de modelos de biomasa y análisis estadístico
Previo al ajuste se verificó la normalidad de los datos con la prueba de Shapiro-Wilk (SW) (α=5 %). Aquí se planteó que si H 0 igual o menor que α, se aceptaría el supuesto de normalidad de la información y se rechazaría H 1 , y si H 1 era mayor que el nivel de significancia de α se concluiría falta de normalidad de los datos, con lo que se aceptaría H 1 . Además, para corregir la heterocedasticidad, se incluyó una función de ponderación basada en la variable combinada (dn 2 AT), como peso en los residuales (α2= σx i k ) (Neter et al., 1996; Álvarez-González et al., 2007): donde σi 2 es la varianza residual de la variable independiente (x) y k es el valor del exponente de optimización, que consiste en usar los errores al cuadrado, del modelo ajustado sin peso ê i 2 como variable dependiente, en el modelo potencial de varianza del error (Harvey, 1976).
Modelos de tipo lineal y no lineal se ensayaron para modelar la biomasa total y por componente estructural (Loetsch et al., 1973; Segura y Andrade, 2008; Méndez-González et al., 2011; Gómez-García et al., 2013). Estos modelos incluyeron como variables independientes a dn y AT. Además, se probó un modelo general que considera dt, Atc, la altura y diámetro del fuste limpio (Afl y dfl) y la sumatoria de los diámetros basales de todas las ramas vivas en el árbol (Σdb) (Cuadro 2).
Cuadro 2 Modelos ajustados para estimar la biomasa total y por componente estructural.

y: biomasa total o por componente estructural, bi: parámetros a estimar, e: término del error del modelo.
El ajuste estadístico de las ecuaciones 1 a 4 se realizó con el procedimiento MODEL, de SAS 9.2, mediante mínimos cuadrados ordinarios (SAS Institute Inc., 2008). La elección del modelo más eficaz se basó en los valores menores de la raíz del cuadrado medio del error (RCME) y valor mayor del coeficiente de determinación ajustado por el número de parámetros (R 2 a).
Los cuatro modelos seleccionados por componente estructural se ajustaron simultáneamente con la técnica de ecuaciones aparentemente no relacionadas (SUR o seemingly unreleated regressions). Esto lo consideró Álvarez-Gonzáles et al. (2007) al incluir como variables endógenas o dependientes a las y y exógenas a las variables medidas directamente en el bosque. De esta forma en el ajuste se involucraron los errores conjuntos y se obtuvieron estimadores consistentes en los componentes de biomasa (Hernández et al., 2013), además de los estadísticos de ajuste RCME, R 2 a, significancia de los parámetros y error estándar aproximado (EEa) (Álvarez-González et al., 2005).
El modelo 5 incluyó las variables dt, Atc, Alf y dfl, además de la sumatoria de los diámetros basales de todas las ramas vivas en el árbol (Σdb) y se ajustó con los métodos de selección backward (eliminación hacia atrás) y forward (selección hacia adelante) como lo recomienda Volke (2008). Así, en el modelo se incluyen solo las variables que explican con precisión mayor la variable respuesta. El modelo más adecuado se seleccionó con los criterios de bondad de ajuste ya descrito, el valor menor del estadístico Cp de Mallows, el valor mayor en R 2 y la significancia parcial de la prueba de F.
La precisión en las estimaciones de biomasa total y por componente estructural de las mejores ecuaciones se evaluó a través del sesgo y la diferencia agregada en porcentaje, de acuerdo con Barrero et al. (2015), y de las fórmulas propuestas por Fonseca et al. (2009), Lencinas y Mohr-Bell (2007) y Vargas-Larreta et al. (2010).
Factor de expansión de biomasa (FEB)
El FEB se estimó de tres maneras. La primera fue a través del cociente de la biomasa aérea total y la biomasa del fuste (Dauber et al., 2002). Esto permitió identificar la distribución y cambiar las proporciones de la biomasa área de los componentes estructurales del árbol o de la especie con el incremento de las dimensiones o la edad de los árboles. FEB se estimó con las fórmulas 6 y 7 (Domínguez-Cabrera et al., 2009; Rodríguez-Ortiz et al., 2012). Los resultados se sumaron y promediaron para definir FEB en las plantaciones por edad (E) y categoría diamétrica (CD).
 (6)
(6)
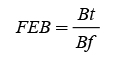 (7)
(7)
donde B f , B h y B r es la biomasa fuste, biomasa follaje y biomasa de ramas.
La segunda metodología para determinar FEB consideró la proporción directa del volumen (V, m3) con respecto a la biomasa de fuste y a la biomasa total (kg) de cada árbol. FEB permite identificar las existencias volumétricas en las plantaciones en relación a la biomasa. El factor se estimó con la estructura estudiada por Ruíz-Aquino et al. (2014), para biomasa de fuste (expresión 8) y esa proporción se calculó con la biomasa total mediante la ecuación 9.
 (8)
(8)
 (9)
(9)
En el tercer método para estimar FEB se definió la relación lineal de los componentes de biomasa con el volumen, donde el valor que multiplica al volumen es la biomasa promedio (kg) por m3 que puede obtenerse. Esta relación se representa con las expresiones 10 y 11, para biomasa total y de fuste, respectivamente.
 (10)
(10)
 (11)
(11)
donde α 0 y α 1 son los parámetros a estimar y representan FEB de biomasa total y fuste, respectivamente.
Comparación de los métodos de estimación de FEB
Los promedios estimados de B t y B f por árbol y población se sometieron a ANDEVA de una vía (Martínez-González et al., 2006) para determinar la existencia de diferencias estadísticas significativas. Las hipótesis fueron: H 0 =u 1 =u 2= u 3 . Las medias de las estimaciones son iguales, aceptando la hipótesis de igualdad. H 1 = al menos una de las medias es distinta y se rechaza la hipótesis de igualdad.
Al contrastar la hipótesis de igualdad de medias (u k ) por árbol y población, la regla de decisión que se aplicó fue con base en la significancia de F en la prueba de Duncan (Pd) agrupando la muestra por método con significancia del 5 %.
Inventario de biomasa aérea
Para observar la aplicabilidad de los resultados en un inventario forestal y los FEB, tres ecuaciones de volumen total generadas en la región de Huimanguillo, Tabasco, en 2015 (12), 2012 (13) y 2007 (14) se compararon y se emplean en las PFC clónales de E. urophylla para la estimación de volumen.
 (12)
(12)
 (13)
(13)
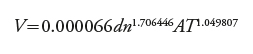 (14)
(14)
Para corroborar la aplicabilidad de las ecuaciones generadas en un inventario forestal, un análisis con datos de inventario se realizó con un diseño de muestreo simple aleatorio, los estimadores del inventario se obtuvieron en 28 sitios rectangulares, de 500 m2, medidos en 2014, en siete plantaciones de E. urophylla de uno a siete años de edad. La comparación de los resultados se realizó mediante análisis de varianza entre las estimaciones con la prueba de t-Cramer con confiabilidad del 95 %, para verificar si los cálculos eran iguales entre sí.
Resultados y Discusión
Distribución de biomasa aérea
La biomasa por componente estructural de las plantaciones promedió 43.0 %, 44.8 % y 37.8 % del peso en la cosecha de fuste, ramas y follaje. La distribución de biomasa por componente indicó que su proporción en el fuste y su volumen aumentaron con el incremento de dn y AT. Lo opuesto ocurrió con la proporción de biomasa de ramas y follaje (Ferrere et al., 2008; Soriano-Luna et al. 2015).
Los árboles clónales de E. urophylla establecidos en PFC concentraron 92.42 % de la biomasa total en el fuste, 5.54 % en las ramas y 2.03 % en el follaje (Cuadro 3). Estos resultados contrastaron con los de Cerruto et al. (2015) de plantaciones de E. grandis en Brasil, pues a los 5.5 años de edad la proporción fue 82 % para fuste y 18 % para ramas, hojas y corteza. Gomes et al. (2013) señalaron intervalos máximos de biomasa en el fuste de E. urophylla, de 3.6 años de edad, 76.2 a 82.5 % en Brasil; Geldres et al. (2006) determinaron en plantaciones de E. nitens, de 4 a 7 años de edad, 84.1, 9 y 6.8 % de fuste, ramas y hojas. Álvarez-González et al. (2005) observaron en plantaciones de E. globulus, de 13 a 24 años, proporciones de 85.9 % de fuste y corteza y 11 y 3.1 % de ramas y follaje. Estos resultados permiten plantear una hipótesis para investigaciones futuras, comparativamente las plantaciones tienen velocidad mayor de crecimiento y eficiencia fotosintética mayor.
Cuadro 3 Distribución porcentual de biomasa por categoría diamétrica (CD) de los componentes estructurales en árboles de Eucalyptus urophylla.
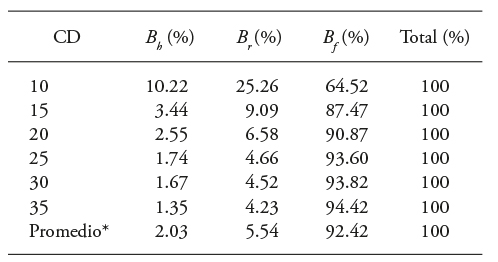
CD: categoría diamétrica (cm), B f : biomasa de fuste, B r : biomasa de ramas, B h : biomasa de follaje, B t : es la biomasa total, Promedio*: el promedio está representado para todos los diámetros.
Ajuste y análisis estadístico de modelos de biomasa
El primer ajuste indicó que los modelos son adecuados para estimar B f , B r, B h y B t ; aunque con problemas de heterocedasticidad (Figura 1).
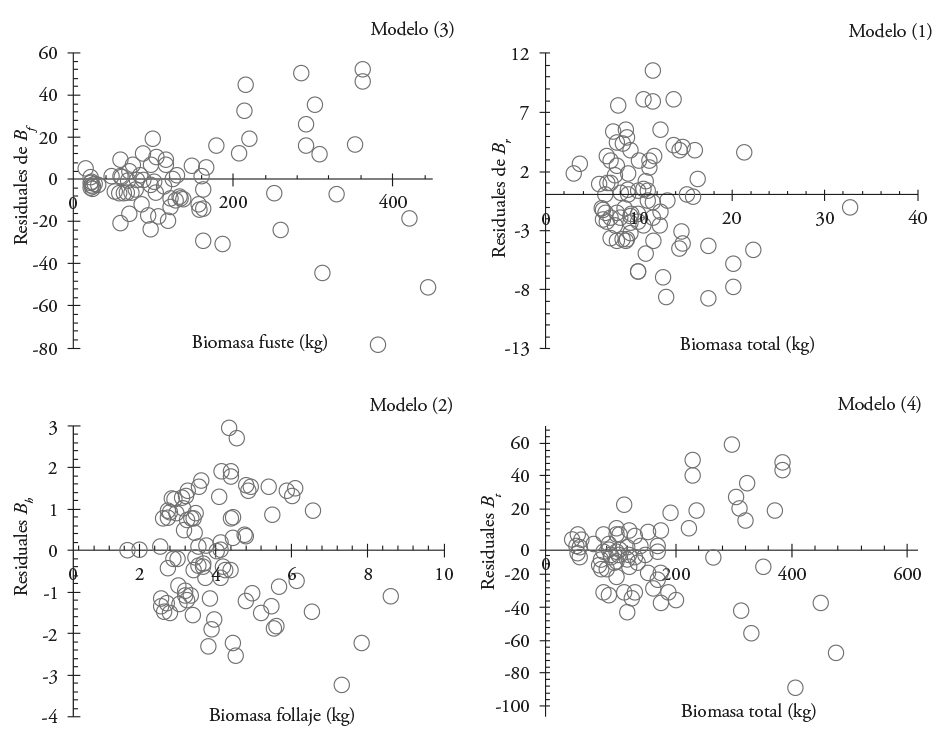
Figura 1 Distribución de residuales de modelos utilizados para estimar la biomasa de cada componente estructural y la total en Eucalyptus urophylla en Huimanguillo, Tabasco, México.
La corrección por heterocedasticidad y el ajuste de los modelos con la técnica de ecuaciones aparentemente no relacionadas (SUR) (Cuadro 4) garantizó la distribución homogénea de los residuales en la varianza de los errores constante (Figura 2). Con la corrección y ajuste nuevo de los modelos se construyó el primer sistema de ecuaciones de biomasa (SI).
Cuadro 4 Valores de ajuste y parámetros de los modelos de biomasa para árboles clonales de Eucalyptus urophylla en Huimanguillo, Tabasco, México.
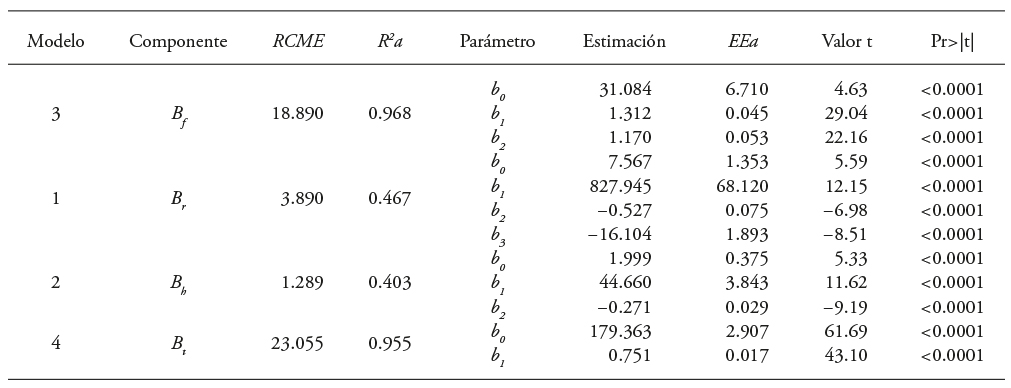
B f , B r , B h y B t : biomasa del fuste, ramas, hojas y total, respectivamente RCME: raíz del cuadrado medio del error. R 2 a: coeficiente de determinación ajustado. EEa: error estándar aproximado de los parámetros, Valor de t: valor de la distribución de t de Student, Pr>|t|: probabilidad asociada al valor de t de Student.
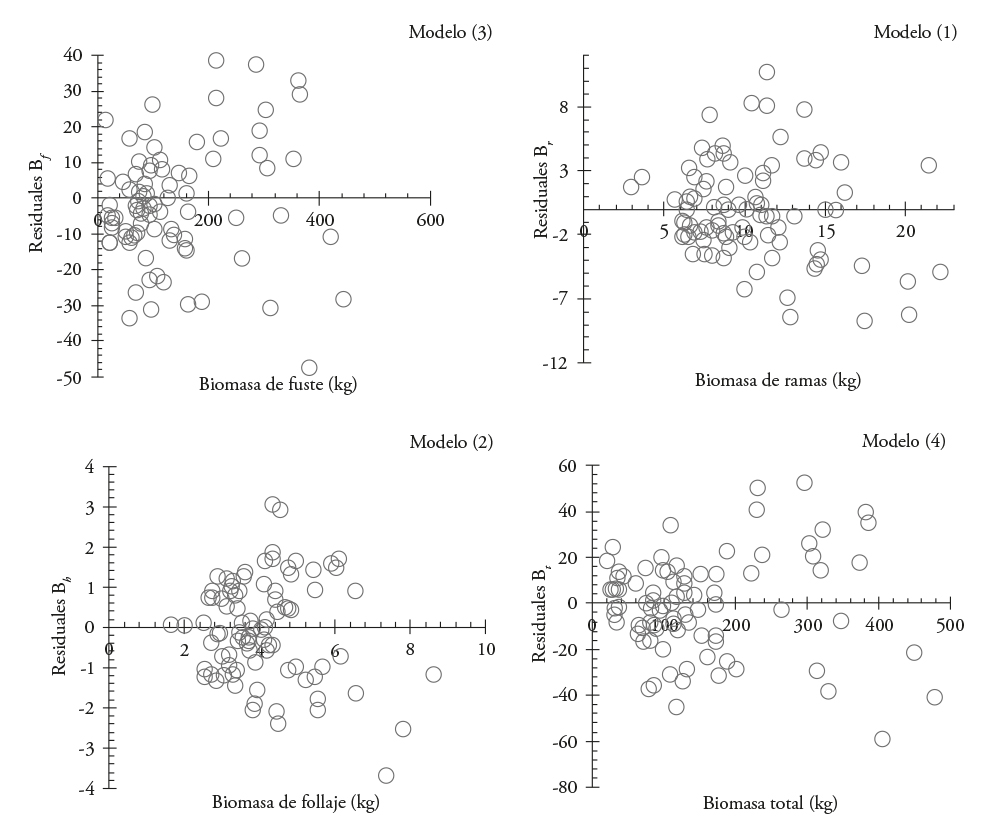
Figura 2 Distribución de residuales corregidos por heterocedasticidad de los modelos utilizados para estimar la biomasa por componente estructural y total en Eucalyptus urophylla en Huimanguillo, Tabasco, México.
Para los tres componentes estructurales y la biomasa total en la prueba de SW se obtuvieron valores entre 0.91 y 0.98. Estos valores señalan que no hubo violación del supuesto de normalidad de la regresión, en los cuatro casos. El resultado es similar a lo que describieron Balzarini et al. (2008) y aplicaron Gaillard et al. (2014).
El modelo 5, ajustado con los procedimientos de regresión y selección de variables “backward” y “forward”, permitió identificar los siguientes modelos más adecuados al incluir variables adicionales, que explican con precisión mayor la predicción de la biomasa de ramas y follaje:
 (15)
(15)
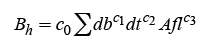 (16)
(16)
Estos modelos se ajustaron simultáneamente y corrigieron por heterocedasticidad con la función de potencia (Figura 3); lo anterior originó un segundo sistema de ecuaciones (SII) para la estimación de biomasa de fuste (3), biomasa total (4) y biomasa de ramas y follaje (15 y 16). Este sistema tuvo ajuste estadístico mayor (Cuadro 5); lo que lo hace estadísticamente más confiable. La prueba de SW demostró la normalidad de los datos con valores entre 0.91 y 0.97 para los cuatro modelos ajustados.
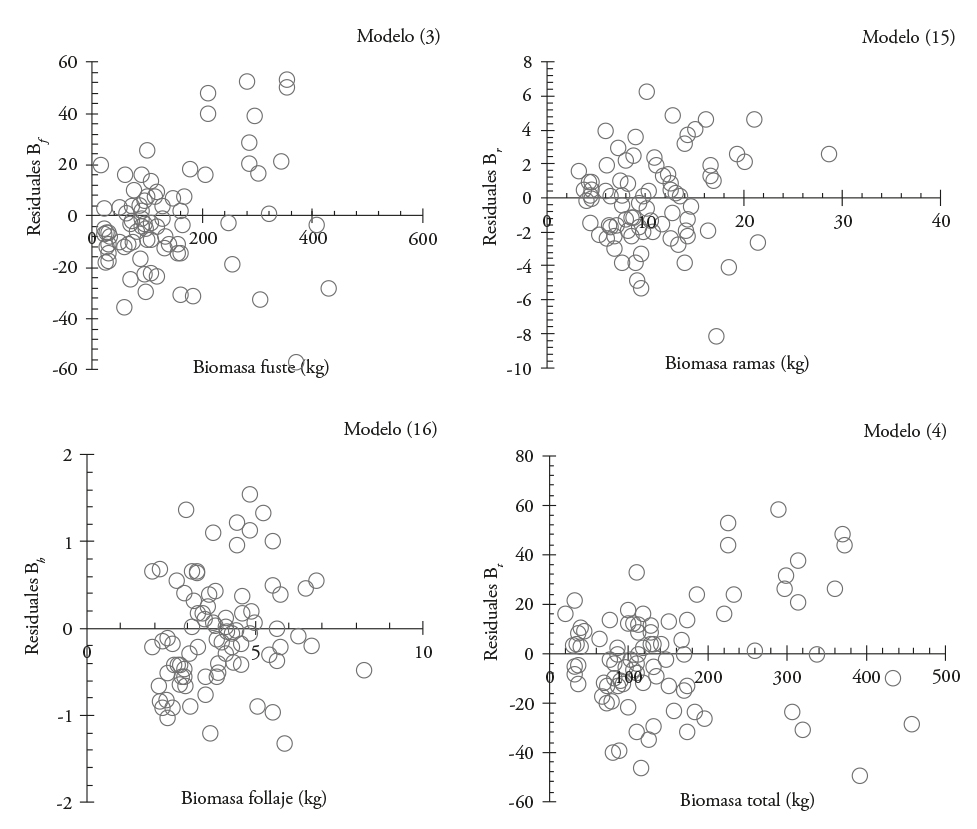
Figura 3 Distribución de residuales al corregir por heterocedasticidad los mejores modelos ajustados para estimar biomasa por componente estructural en Eucalyptus urophylla en Huimanguillo, Tabasco, México.
Cuadro 5 Parámetros y estadísticos de ajuste de los modelos al incluir las variables de altura de fuste limpio y sumatoria de los diámetros basales de las ramas por componente estructural de biomasa para Eucalyptus urophylla en Huimanguillo, Tabasco, México.
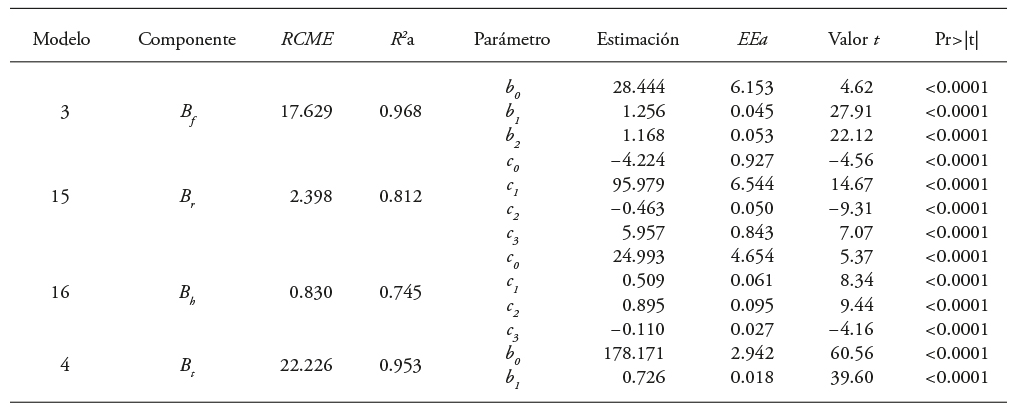
B f , B r , B h y B t : biomasa de fuste, ramas, hojas y total, respectivamente, RCME: Raíz del cuadrado medio del error, R 2 a: coeficiente de determinación ajustado, EEa: error estándar aproximado.
Según los resultados, aunque los valores de los parámetros de los modelos seleccionados son cercanos, SII sobreestimó ligeramente B f , B r y B t , y la estimación de la B h fue conservadora en la predicción por individuo y en la diferencia total por muestra. El primer sistema (SI) de acuerdo al valor de los estadísticos de precisión subestimó todas las variables e hizo conservadores los cálculos de biomasa aérea (Cuadro 6).
Cuadro 6 Estadísticos de evaluación de la capacidad predictiva de biomasa total y por componente de los sistemas de ecuaciones para Eucalyptus urophylla en Huimanguillo, Tabasco, México.

Debido a la precisión de los estadísticos de ajuste de los sistemas de ecuaciones (SI y SII) y porque se generaron para estimar biomasa aérea total y de sus componentes, sin utilizar métodos destructivos, pueden estimarse la biomasa total y de los componentes con la información de inventario forestal tradicional o con variables adicionales. Estas pueden obtenerse a través de instrumentos de medición electrónica, como el dendrómetro (Criterium RD 1000 ®).
Factores de expansión de biomasa (FEB)
El FEB promedio obtenido con las ecuaciones 6 y 7 mostró que 17 % de la biomasa aérea se distribuye en ramas y follaje, y 83 % está en el fuste (Cuadro 7). Esta distribución concordó con lo reportado por Téllez et al. (2008) para plantaciones de E. urophylla en Oaxaca, México, con 17.6 % de biomasa de ramas y follaje y 82.4 % en fuste. Los resultados de Álvarez-González et al. (2005) con E. globulus, en España, fueron 14.1 % de biomasa de hojas y ramas y 85.9 % en fuste y los de Ferrere et al. (2008), en Argentina, mostraron la distribución de biomasa en E. viminalis con 20 % en ramas y follaje y 80 % en fuste.
Cuadro 7 Factores de expansión de biomasa (FEB) por categoría diamétrica para datos de biomasa aérea.

Las estimaciones con los sistemas I y II subestimaron FEB en 1.51 % y 1.48 %. SII se aproximó mejor a los valores evaluados por categoría diamétrica, sin considerar la edad de la plantación.
Al considerar la edad en el análisis, SI, en promedio, tuvo aproximaciones mejores, pero subestimó FEB en 2.5 %. En este caso se propone que 19 % de biomasa correspondió a ramas y follaje y 81 % a fuste. Esta situación es semejante a lo referido en las proporciones de biomasa por componente estructural (Cuadro 8). En ambos sistemas FEB para E. urophylla decrecieron a medida que la biomasa total aérea aumentó (Rodríguez-Ortiz et al. 2012; Chávez-Pascual et al., 2013).

Con el modelo 8 de FEB, para B t , el promedio de biomasa total por cada m3 de volumen maderable fue 600.94 kg, En este caso el factor disminuye con el incremento de las dimensiones de los árboles. Para las categorías diamétrica de 10 cm y 15 cm existieron en promedio 795.05 kg m-3, para 20 y 25 cm fue 557.94 82 kg m-3 y para 30 cm y 35 cm fue 447.82 kg m-3. Los resultados por edad indicaron que el promedio para estas plantaciones, con uno a siete años, fue 652.22 kg m-3, y presentaron tendencia semejante de 7 a 4 años de edad, con 531.06 kg m-3, a los 3 años 613.34 kg m-3 y aumentó a 812.25 kg m-3 en las plantaciones de 1 año de edad. Lo anterior se debe a la gran cantidad de biomasa foliar que tienen las plantaciones jóvenes.
En el caso de FEB para B f los promedios fueron 496.45 kg m-3 por CD y 503.02 kg m-3. Los resultados analizados por CD mostraron 508.76 kg m-3, 500.03 kg m-3 y 480.45 kg m-3 para las plantaciones con dimensiones de 10 y 15 cm, 20 y 25 cm, y 30 y 35 cm, respectivamente. En ambos casos FEB disminuyeron con el crecimiento de los árboles y el aumento de la edad. Esto se debió a que la acumulación de biomasa en el fuste aumenta conforme el individuo disminuye la biomasa aérea foliar, lo que sucede a medida que las categorías diamétricas o edades aumentan.
Los modelos (10) y (11) para estimar B t y B f que consideraron la relación lineal de estas variables con el volumen del árbol (m3) mostraron FEB de 510.06 kg m-3 y 472.54 kg m-3 (Cuadro 9). Estos resultados fueron semejantes a los de los modelos 8 y 9.
Cuadro 9 Estadísticos de bondad de ajuste y parámetros de los modelos 10 y 11 para estimar Bt y Bf en árboles de Eucalyptus urophylla de PFC en Huimanguillo, Tabasco, México.
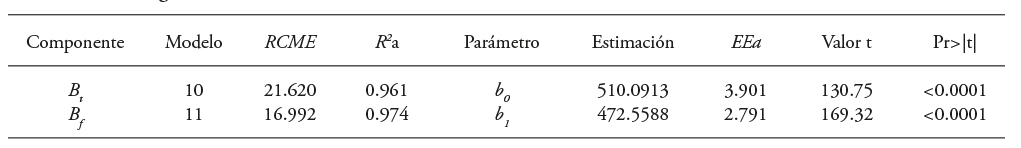
RCME: raíz del cuadrado medio del error, R 2 a: coeficiente de determinación ajustado por el número de parámetros, EEa: error estándar aproximado, b 0 y b 1 : parámetros estimados, Valor de t: valor de la distribución de t de Student, Pr>|t|: probabilidad asociada al valor de t de Student.
Comparación de métodos de estimación e inventario de biomasa aérea
Los resultados del análisis de varianza de las medias de los datos observados versus las tres estimaciones con SI y SII y los modelos de proporción directa, relacionados con el volumen (10 y 11) permitieron concluir que las variables B t y B f fueron iguales entre sí (Pr<F=0.963 para B t y Pr>F>0.997 para B f ). Por lo tanto, se acepta la hipótesis nula (H 0 ) y puede afirmarse que los tres métodos propuestos son estadísticamente iguales y confiables para estimar estas variables. Ellos difieren solo en la estructura matemática de los modelos.
Una comparación de resultados, tomando como referencia los modelos de biomasa de fuste y biomasa total de los modelos 3 y 4 del primer sistema de ecuaciones, demostró que el modelo de volumen 12 es el más adecuado para estimar la biomasa aérea con datos de un inventario, le siguió el modelo 13 y luego el 14 (Cuadro 10). El análisis de varianza entre las estimaciones, con la prueba de t-Cramer, indicó que son iguales entre sí, debido a que el valor de probabilidad en el ANDEVA fue alto (F=2.6887 y p=0.7978).
Cuadro 10 Estimadores con el muestreo simple aleatorio para PFC de Eucalyptus urophylla de entre uno y siete años de edad establecidas en Huimanguillo, Tabasco, México con los modelos 10 y 11.
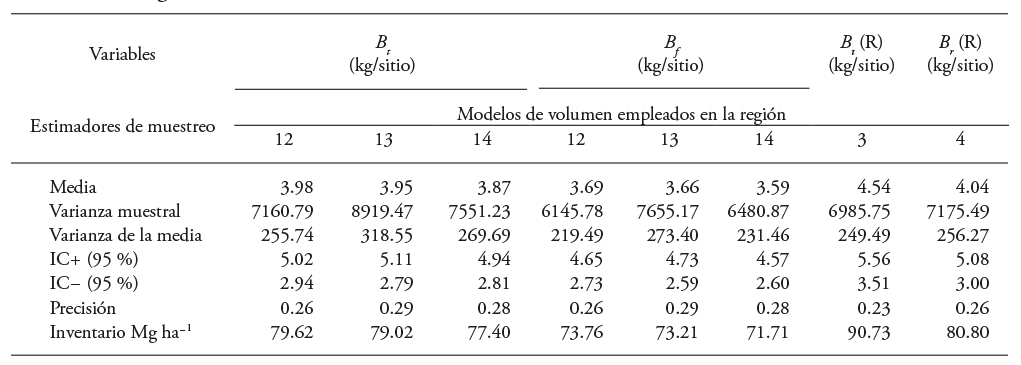
B f , B r , B h y B t : biomasa de fuste, ramas, hojas y total, respectivamente, R: valor de la biomasa de referencia, IC+: limite superior del intervalo de confianza al 95 %, IC-: limite inferior del intervalo de confianza.
Los datos de inventario, analizados con los estadísticos de muestreo simple al azar, indicaron en promedio 156.08 m3 ha-1 de volumen en PFC de E. urophylla, 14 cm en diámetro normal, 18 m de altura y densidad de 986 árboles ha-1. Además, al incorporar los resultados del Cuadro 10 en el modelo 4 de B t el volumen (m3 ha-1) se estimó en 80.80 Mg ha-1 de biomasa aérea. Esto concuerda con los 70.27 Mg ha-1 de plantaciones de 7 años de E. viminalis en Argentina (Ferrere et al., 2008). Pero, el valor es inferior a los rendimientos reportados en plantaciones de 5, 6 y 7 años de E. nitens, (rendimiento de 73.1, 111.8 y 159.5 Mg ha-1), con densidad de plantación de 1,500 árboles por hectárea, antes de aclarearse (Geldres et al., 2006).
La biomasa aérea por hectárea fue similar a los rendimientos (142 Mg ha-1) estimados en plantaciones de E. globulus de España, pues la cantidad reportada en este estudio es, en promedio, la mitad de lo mencionado en edad de 13 años por Álvarez-González et al. (2005). Además, estas estimaciones de acuerdo con los escenarios propuestos por Seppánen (2002 y 2003), para E. urophylla, E. grandis y el híbrido E. grandis x E. urophylla son bajas, porque a la edad de 7 años se espera tener rendimientos promedio de biomasa aérea de 147.7 Mg ha-1 en esta región, en el escenario menos favorable.
Conclusiones
La correlación alta entre el diámetro normal y la altura total de un árbol con la biomasa aérea permite obtener modelos alométricos para estimar la biomasa total y los componentes estructurales de Eucalyptus urophylla. La inclusión de variables adicionales, como altura de fuste limpio y sumatoria de los diámetros basales de todas las ramas vivas, en los modelos de biomasa de ramas y hojas, mejora estadísticamente los modelos y la precisión de las ecuaciones evaluadas.
La identificación de la relación directa entre el volumen y la biomasa aérea permite aplicar las ecuaciones de biomasa generadas y los sistemas propuestos con datos de un inventario forestal. Las PFC analizadas a la edad del turno definido en siete años tienen en producción promedio de biomasa aérea de 80.89 Mg ha-1.
Las ecuaciones de biomasa aérea y los factores de expansión obtenidos son confiables para utilizarlos en inventarios forestales. Así, son herramienta que facilitan la planeación y ejecución de las actividades de manejo forestal implementadas para E. urophylla, y son útiles para evaluar y cuantificar los beneficios adicionales de las plantaciones, como cuantificar el C capturado y acumulado en las plantaciones forestales. Esto es indispensable para negociar bonos de C en los mercados internacionales.

