











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La orquídea Laelia autumnalis es nativa de México, presenta inflorescencias con flores púrpura vistosas, de diversas tonalidades, formas, tamaños y aromas. Esta especie se usa como ornamental desde la época prehispánica por distintos pueblos indígenas, aunque nunca ha sido objeto de un programa de mejoramiento genético. Los ejemplares silvestres se comercializan ilegalmente en mercados locales y regionales de diversas comunidades de México a precios muy bajos (Beltrán-Rodríguez et al., 2012). La generación de variedades nuevas y mejoradas puede aumentar el valor y la aceptación de esta especie en el mercado nacional y constituir una ventaja económica a la hora de aprovechar un mercado de exportación.
El mejoramiento por mutagénesis es uno de los métodos más eficientes para obtener nuevos cultivares de plantas (Aros et al., 2012; Kazi, 2015), en particular ornamentales como las orquídeas (Kikuchi, 2000; Luan et al., 2012; Lee et al., 2015), anturio (Puchooa, 2005), y crisantemo (Yamaguchi et al., 2008; Kumar et al., 2012). Si se combina el uso de radiaciones ionizantes con la técnica de cultivo in vitro se reduce el tiempo y los costos en el desarrollo de un nuevo cultivar (Yunus et al., 2013); se puede obtener una producción continua de plantas a gran escala, así como mantener y multiplicar a los individuos mutantes (Barakat y El-Sammack, 2011; Ángeles-Espino et al., 2013). Además, la variación somaclonal que se puede presentar durante la micropropagación ofrece la posibilidad de incrementar la frecuencia de mutaciones génicas y puntuales, que son las más importantes en el mejoramiento genético (Estrada-Basaldua et al., 2011). El cultivo in vitro es indispensable para la germinación asimbiótica de semillas de orquídeas, proceso que en su hábitat requiere de la simbiosis con hongos micorrícicos (Verma et al., 2014).
El uso de radiaciones ionizantes en las plantas ha mejorado una gama amplia de caracteres como la arquitectura, el rendimiento, la floración, la tolerancia a estrés biótico y abiótico (Kon et al., 2007). La principal ventaja es su capacidad para cambiar uno o pocos caracteres en un cultivar excelente sin modificar el resto del genotipo (Patil et al., 2015). La eficiencia de estas radiaciones es alta porque cerca de 89 % de las variedades vegetales obtenidas por mutagénesis se desarrollaron con mutágenos físicos como los rayos X, rayos gamma y neutrones rápidos; dentro de estos agentes mutagénicos los más usados en especies cultivadas (granos, frutales, y ornamentales) son los rayos gamma, pues con esta técnica se ha desarrollado 60 % de las variedades vegetales mutantes (Kon et al., 2007).
Los mutágenos alteran el ADN y causan mutaciones puntuales, deleciones y aberraciones cromosómicas (Tanaka et al., 2010), y los individuos nuevos tienen cambios importantes en su estructura genómica (Emmanuel y Levy, 2002). La técnica de mutagénesis tiene eficiencia especial en plantas con periodos juveniles largos antes de la floración y la producción de semillas (Predieri, 2001), así como en plantas ornamentales nativas con variabilidad genética limitada (Lee et al., 2008).
Los mutágenos físicos, en particular las radiaciones gamma, son partículas pequeñas de radiación ionizante que tienen capacidad alta de energía penetrable en los tejidos biológicos (Sadhukhan et al., 2015); su longitud de onda es 10 nm (Kitano et al., 2015). Su efecto biológico se basa en la interacción con átomos o moléculas en la célula, en particular con el agua, para producir radicales libres y efectuar cambios en bases y rupturas en las uniones de hidrógeno entre cadenas complementarias de ADN (Kovács y Keresztes, 2002). Estos radicales pueden dañar o modificar componentes importantes de las células vegetales y cambian la morfología, anatomía, bioquímica y fisiología de las plantas, en función de la dosis de radiación (Wi et al., 2007). Estos efectos incluyen cambios en la estructura celular y el metabolismo de la planta (Kovács y Keresztes, 2002), como dilatación de las membranas de los tilacoides, alteración en la fotosíntesis, modulación del sistema antioxidante y acumulación de compuestos fenólicos (Wi et al., 2005). El núcleo celular es el principal organelo afectado por la radiación ionizante (Pavan et al., 2013).
El efecto de la radiación en los tejidos celulares se divide en tres fases, la física o inicial que dura sólo una fracción de segundo, la etapa química que dura unos pocos segundos y la etapa biológica en la cual la escala de tiempo varía de decenas de minutos a decenas de años (Pavan et al., 2013).
El intervalo en el cual se favorece la aparición de mutaciones útiles en los programas de mejoramiento genético, es la dosis letal media (DL50) o dosis reductiva media (GR50), que es la cantidad de radiación absorbida con la cual sobrevive 50 % de la población expuesta o se reduce el crecimiento en 50 %, por lo que es importante conocer este rango (Ángeles-Espino et al., 2013) antes de iniciar un programa de mejoramiento genético asistido por mutagénesis. La DL50 es única para cada especie, genotipo e incluso en diferentes tejidos de una planta. En callos generados de flores liguladas de Chrysanthemum morifolium cv. Delistar White con DL50 de 0.5 Gy, se obtuvieron modificaciones en la forma de la flor y el número de flores por inflorescencia (Barakat et al., 2010). En callos embriogénicos de Tricyrtis hirta la DL50 se determinó en 20 Gy y se encontraron variantes con enanismo, con hojas más verdes y delgadas y mayor número y tamaño de flores (Nakano et al., 2010). En semillas de Moluccella laevis la DL50 de 25 Gy aumentó el número de flores por vara (Minisi et al., 2013). Para explantes de hoja en Torenia fournieri la DL50 fue 63 Gy para plantas diploides y 72 Gy para las poliploides, en ambos casos se obtuvieron mutantes con hojas rojas (Chanchula et al., 2015).
Para orquídeas nativas de México no se conoce la radiosensibilidad de los tejidos a la radiación gamma, ni se ha determinado la dosis óptima para obtener variantes de importancia ornamental. Por lo anterior, el objetivo de este estudio fue determinar la DL50 y GR50 con rayos gamma (60Co) en protocormos de Laelia autumnalis cultivados in vitro. La hipótesis fue que existe una dosis óptima para generar variantes en L. autumnalis a partir de protocormos irradiados.
Materiales y Métodos
Esta investigación se efectuó en el Laboratorio de Cultivo in vitro de Tejidos Vegetales de la Facultad de Agrobiología de la Universidad Michoacana de San Nicolás de Hidalgo, ubicado en Uruapan, Michoacán, México (19° 23’ 41’’ N y 102° 03’ 31’’), desde mayo de 2014 a marzo de 2015. La irradiación de protocormos se hizo en el Departamento del Irradiador Gamma del Instituto Nacional de Investigaciones Nucleares (ININ), ubicado en Ocoyoacac, Estado de México.
Material vegetal
En este estudio se utilizó la semilla polvo de seis frutos obtenidos mediante autofecundación manual de plantas de L. autumnalis de la colección del Banco de Germoplasma del Sistema Nacional de Recursos Fitogenéticos.
Medio de cultivo
Un medio con las sales minerales de Murashige y Skoog (1962) (MS) fue usado, sin fitohormonas y adicionado con sacarosa (30 g L-1), mio-inositol (100 mg L-1), tiamina (0.4 mg L-1) y agar (6 g L-1); el pH se ajustó a 5.7±0.1 con NaOH 1 N. De este medio se sirvieron 20 mL en frascos de vidrio de 100 mL de capacidad y se esterilizaron en autoclave durante 15 min a 1.2 kg cm-2 de presión y 121 °C.
Establecimiento del cultivo aséptico
Para la siembra se tomaron 33 muestras de semillas (20 mg cada una) y se colocaron en jeringas de 10 mL, y se efectuó la desinfección con hipoclorito de sodio comercial 15 % v/v (6 % de ingrediente activo) por 15 min. En la campana de flujo laminar se retiró la solución desinfectante y se hicieron tres enjuagues con agua estéril. La semilla polvo de cada muestra se suspendió en 0.5 mL de agua estéril y se colocó en un frasco con medio de cultivo. La incubación se efectuó con fotoperiodo de 16/8 h luz/oscuridad y radiación fotosintéticamente activa de 45 μE m-2 s-1 proporcionada por lámparas de luz fluorescente blanca de 75 W.
Irradiación con rayos gamma 60Co
A los 45 d después de la siembra (dds), los protocormos obtenidos se trataron con diez dosis de irradiación (5, 10, 15, 20, 25, 30, 35, 40, 45 y 50 Gy), más un tratamiento testigo sin irradiar. El diseño experimental fue completamente al azar con 11 tratamientos y tres repeticiones, y la unidad experimental fue un frasco con los protocormos obtenidos de la germinación de 20 mg de semilla, cantidad que fluctuó entre 500 y 700 protocormos. Un día después de la irradiación (46 dds), los protocormos de cada frasco se trasfirieron a medio de cultivo fresco. A los 90 dds se transfirieron a cajas Petri para facilitar la evaluación. En todo este tiempo, el desarrollo de los protocormos se dividió en tres etapas fenológicas: protocormos fotosintéticos, protocormos con promeristemos y protocormos con hojas. Estas fases se observaron de manera simultánea en todas las unidades experimentales, porque el desarrollo de plántulas en L. autumnalis es un proceso asincrónico.
A los 135 dds, los protocormos se subcultivaron en contenedores de plástico de 250 mL con 50 mL de medio basal fresco y se incubaron en las condiciones ambientales ya descritas.
Variables evaluadas en protocormos
Porcentajes de supervivencia de protocormos, de protocormos fotosintéticos, de protocormos con promeristemos y con hojas. Cada unidad experimental (frasco) se dividió en cuadrantes para facilitar los conteos. En cada cuadrante se contó el número de protocormos vivos, muertos, fotosintéticos, con promeristemos y con hojas. Con la suma de protocormos en cada etapa fenológica se obtuvo el número total de protocormos por unidad experimental, que se usó como denominador para obtener los porcentajes de supervivencia, de protocormos fotosintéticos, de protocormos con promeristemos y con hojas de la siguiente forma: % de supervivencia por unidad experimental = (Número de protocormos vivos por frasco/Número total de protocormos contenidos en el frasco) X 100. En cada variable se utilizó el mismo procedimiento.
Materia fresca de protocormos. Todos los protocormos por unidad experimental se pesaron en una balanza analítica marca OHAUS® con legibilidad de 0.01 mg.
Estas variables se evaluaron 45 d después de la irradiación (90 dds).
Variables evaluadas en plántulas
Longitud de plántula, pseudobulbo y raíz. La distancia entre la base y el ápice de la plántula, pseudobulbo y raíz, se midió con vernier digital.
Anchura de hoja y pseudobulbo. Se midió en la parte media de cada órgano, con vernier digital.
Número de hojas y raíces. Se cuantificó directamente en cada plántula.
Materia fresca de plántulas (mg). Cada plántula se pesó en balanza analítica digital (OHAUS®) con legibilidad de 0.01 mg.
Biomasa de plántulas. Cada plántula se secó 72 h en una estufa marca FELISA, a 70 °C. Las plántulas secas se pesaron en balanza analítica digital (OHAUS®) con legibilidad de 0.01 mg.
El registro de estas variables se realizó en cinco plántulas por repetición, 15 en total por tratamiento, a los 214 d después de la irradiación (259 dds).
Análisis estadístico
Con los datos obtenidos a los 45 y 214 ddi se realizó ANDEVA, se calculó el coeficiente de correlación de Pearson y la prueba de Tukey para la comparación de medias entre tratamientos, con SAS versión 9.0 (Statistical Analysis System, 2002).
Para conocer la magnitud de la respuesta de cada variable a las dosis de radiación, se evaluó un modelo de regresión lineal. Sin embargo, sólo las variables supervivencia y emergencia de hojas se ajustaron a este modelo, por lo cual la determinación de la DL50 y GR50 se realizó sólo para estas variables.
Resultados y Discusión
Efecto de la radiación sobre el desarrollo de protocormos
El análisis de varianza reveló que la radiación influyó en la supervivencia de protocormos, la presencia de protocormos fotosintéticos, de protocormos con promeristemos, con hojas y la materia fresca de protocormos (Cuadro 1).
Cuadro 1 Análisis de varianza para el efecto de la radiación gamma 60Co en la supervivencia y desarrollo de protocormos de Laelia autumnalis
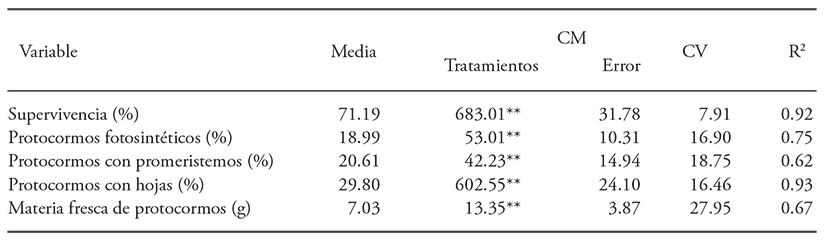
** p≤0.01; ns=no significativo.
La radiación con dosis bajas (5 a 20 Gy) no afectó la supervivencia de protocormos comparado al tratamiento testigo, aunque hubo reducción de protocormos vivos entre 17.6 y 46.4 % en los tejidos tratados con 25 a 50 Gy (Figura 1A). Dosis de 20 a 30 Gy estimularon en los protocormos la presencia de clorofila, que se notó como un cambio de color amarillo claro a verde (Figura 1B), niveles no diferentes a 45 Gy. La acumulación de materia fresca fue similar entre los protocormos del tratamiento testigo y los tratados con los niveles de radiación, aunque distinta entre los irradiados a 5 y 10 Gy respecto a 35 Gy, los primeros con 69 y 66 % más materia fresca acumulada (Figura 1C). Una respuesta similar se observó para la formación de promeristemos, estadísticamente similar en los tratamientos irradiados y el testigo, pero casi el doble en los protocormos irradiados a 20, 30 y 35 Gy, comparado con 50 Gy (Figura 1D).
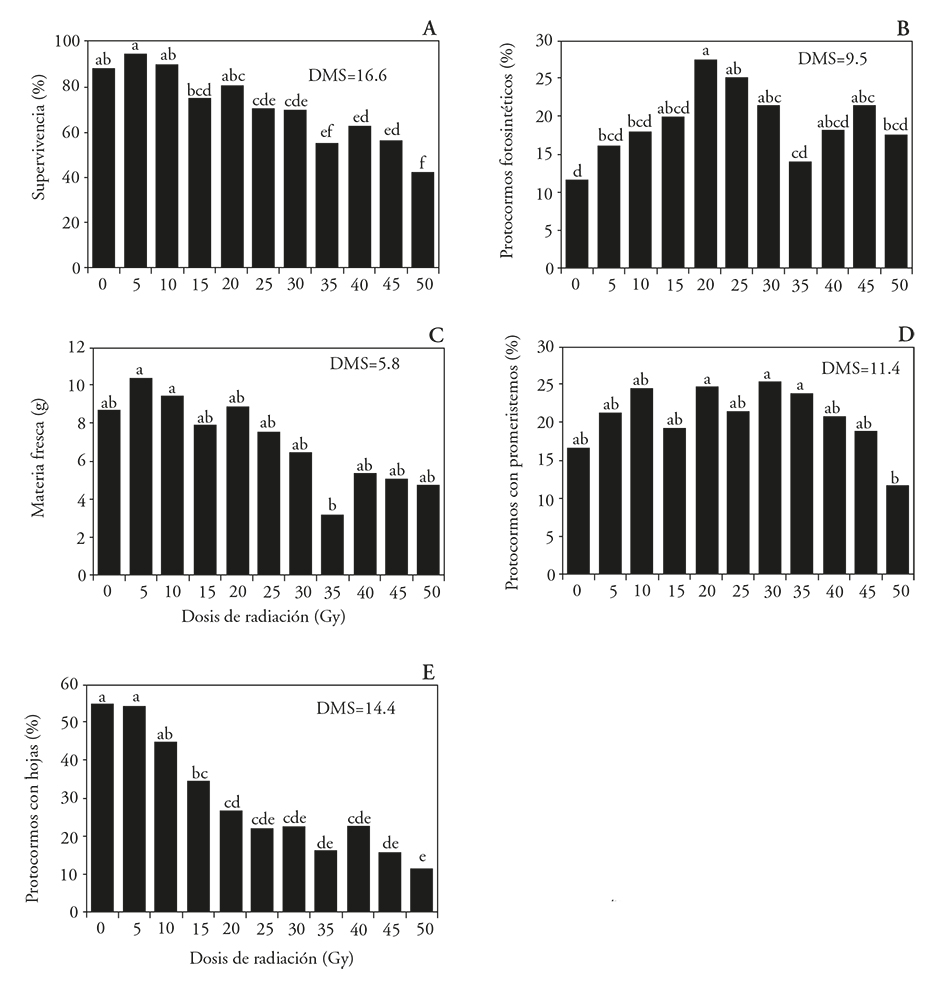
Figura 1 Efecto de la radiación gamma 60Co sobre la supervivencia y el desarrollo de protocormos de Laelia autumnalis, A) supervivencia de protocormos, B) protocormos fotosintéticos, C) materia fresca de protocormos, D) protocormos con promeristemos y E) protocormos con hojas. DMS=Diferencia Mínima Significativa. En cada figura, letras distintas indican diferencias significativas (Tukey; p≤0.05).
La formación de protocormos con hojas no difirió (p>0.05) entre las plántulas no irradiadas y las tratadas con 5 y 10 Gy; desde 15 Gy se redujo (p≤0.05) la formación de protocormos con hojas al aumentar la dosis de irradiación (Figura 1E). El desarrollo de los explantes se aceleró a dosis de radiación baja (radioestimulación), pero se redujo al aumentar la dosis de radiación gamma, como sucede en callos in vitro de Chrysanthemum morifolium (Soliman et al., 2014).
Aunque en nuestra investigación no hubo estudios a nivel molecular, los tejidos se dañan al aumentar la dosis de radiación gama, aunque las células presentan un mecanismo de reparación del ADN cuando están expuestas a cualquier fuente de radiación. Pero si la intensidad de la fuente de radiación es demasiado alta, este mecanismo ya no puede reparar todas las células dañadas, las cuales mueren como ocurre en plantas de jengibre (Zingiber officinale) (Yoon et al., 2014). En las células sobrevivientes se producen radicales libres que ocasionan desórdenes metabólicos (Pavan et al., 2013), se altera el patrón de expresión génica (Corthals et al., 2000), que regula ciertas rutas metabólicas y sistemas de defensa (Zolla et al., 2003), y causa cambios cualitativos y cuantitativos en el contenido total de proteínas solubles (Corthals et al., 2000). Estas proteínas tienen una función importante en la transducción de señales, en la defensa antioxidante, anticongelante, choque térmico, antipatogénesis o síntesis de osmolitos, que son esenciales para las funciones y el crecimiento de las plantas (El-Fiki et al., 2015). Estas alteraciones a nivel celular, causadas a nivel fisiológico o físico, incluidos daños en los cromosomas y la reducción de la supervivencia al aumentar la dosis de radiación, se reportan en Jatropha curcas L. (Dhakshanamoorthy et al., 2011), cuatro variedades comerciales de Lycopersicon esculentum (Ramírez et al., 2006), Withania somnifera (Bhosale y More, 2014), Orthosiphon stamineus (Kiong et al., 2008) y Chrisanthemun (Yamaguchi et al., 2009).
Los coeficientes de correlación fueron negativos y altamente significativos entre los niveles de radiación aplicados y la supervivencia de protocormos (-0.91), protocormos con hojas (-0.90) y materia fresca de protocormos (-0.69). Además hubo correlación positiva entre la supervivencia y los protocormos con hojas (0.89) y materia fresca de protocormos (0.71), y entre la formación de protocormos con hojas y la materia fresca de protocormos (0.65) (Cuadro 2).

Efecto de la radiación sobre el desarrollo de plántulas
El ANDEVA reveló que la radiación influyó en la materia fresca, biomasa y longitud de las plántulas y la anchura de hojas y raíces, pero no afectó el número de hojas, la longitud y ancho de pseudobulbos y el número de raíces. (Cuadro 3).
Cuadro 3 Análisis de varianza para el efecto de la radiación gamma 60Co sobre el desarrollo de protocormos de Laelia autumnalis
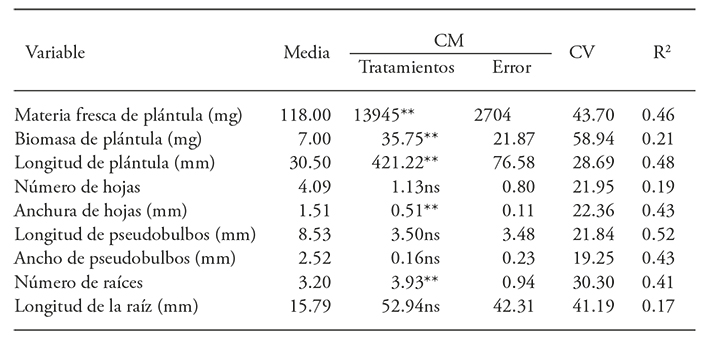
** p≤0.01; ns=no significativo.
La longitud de plántulas aumentó en 32 % al usar radiación a 5 Gy, con relación al tratamiento sin irradiar. Esto sugiere un efecto radioestimulante del crecimiento a dosis de radiación bajas; con 35, 40 y 50 Gy las plántulas fueron 31, 50 y 42 % menos largas que las plántulas sin irradiar (Figuras 2A y 3). La mayor acumulación de materia fresca (189 y 176 mg) se registró en las plántulas tratadas con 5 y 10 Gy, estadísticamente similares al testigo y a 15 Gy, pero superiores a los otros tratamientos (Figura 2B). La anchura de las hojas se redujo en 25 y 29 % con 40 y 50 Gy respecto al testigo, y el número de raíces disminuyó en 37 % con 45 Gy (Figura 2C y D). La acumulación de biomasa fue similar en las plántulas de todos los tratamientos, excepto en las tratadas con 5 y 15 Gy, las cuales acumularon casi el doble de biomasa que las irradiadas con 45 Gy (Figura 2E). La disminución de la biomasa al aumentar la dosis de radiación se ha observado en plántulas de Arabidopsis que inhiben su transporte de carbohidratos cuando se tratan con radiaciones mayores de 50 Gy (Bondada y Oosterhuis, 2003), porque hay daños y desorientación de los tilacoides del grana y aumenta la acumulación de gránulos de almidón dentro de los cloroplastos. Estos cambios en la ultraestructura no se observaron en los cloroplastos de células irradiadas con dosis bajas (Wi et al., 2007).
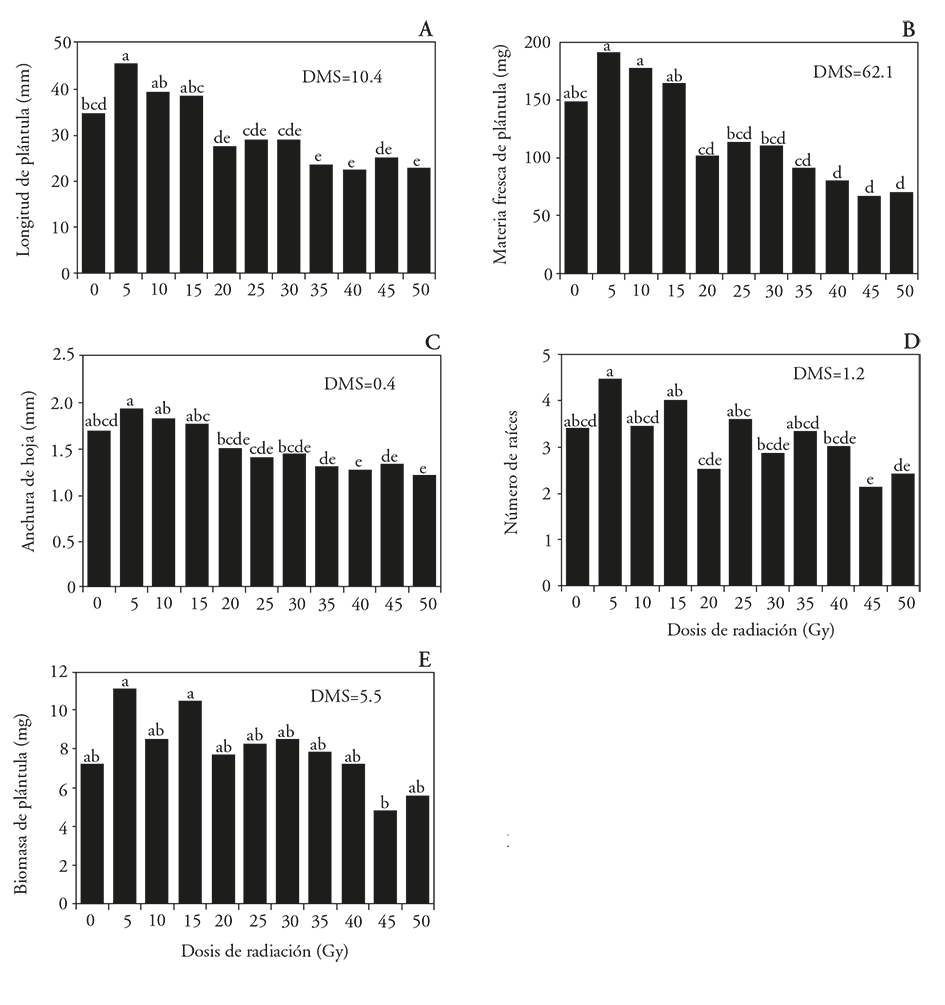
Figura 2 Efecto de la radiación gamma 60Co sobre plántulas de Laelia autumnalis procedentes de protocormos irradiados con 60Co, A) longitud, B) materia fresca, C) anchura de hojas, D) número de raíces y E) biomasa de plántulas. DMS=Diferencia Mínima Significativa. En cada figura, letras distintas indican diferencias significativas (Tukey; p≤0.05).

Figura 3 Plántulas de Laelia autumnalis procedentes de protocormos irradiados con rayos gama 60Co a diferentes dosis de radiación.
En contraste a la radioinhibición, el fenómeno de hormesis observado con 5 Gy sobre el crecimiento de las plántulas de L. autumnalis, se describe como el efecto estimulante de cualquier factor sobre el crecimiento de organismos (Szarek, 2005). Esto se atribuye al cambio en la red de señalización hormonal en las células vegetales o al aumentar la capacidad antioxidante de las células para superar los factores de estrés como las fluctuaciones de intensidad de la luz y la temperatura en las condiciones de crecimiento (Kim et al., 2004). Además, la radiación acelera la tasa de división celular y activa las auxinas, reportado en Gypsophilla paniculata en la cual al irradiar callos con 1 Gy aumentó la longitud de brotes (3.4 cm) (Barakat y El-Sammack, 2011). En L. esculentum hay un intervalo radioestimulante de 5 a 25 Gy para la altura de las plantas (Ramírez et al., 2006); en Triticum durum el impacto de la radiación gamma sobre las semillas aumentó 32 y 75 % en el número y longitud de raíces, al ser irradiadas con 20 Gy (Melki y Marouani, 2010). En la regeneración de callos embriogénicos de Heliconia psittacorum irradiados con rayos gamma 60Co, 30 Gy hubo el porcentaje mayor de callos regenerados (80.8 %), 20. 8 % más que en el tratamiento sin irradiar y 74.8 % más que en los callos tratados con 75 Gy (6 %) (Urrea y Ceballos, 2005).
Los coeficientes de correlación fueron negativos y altamente significativos entre los niveles de radiación aplicados y la materia fresca de plántula (-0.57), longitud de plántulas (-0.56) anchura de hojas (-0.52), número de raíces (-0.41) y longitud de pseudobulbos (-0.40) (Cuadro 4). Esto confirma una correlación negativa entre la energía absorbida por unidad de masa en la dosis aplicada y las variables de crecimiento y desarrollo de las plantas (Giovannini et al., 2015). En Arabidpsis esta inhibición es atribuida a la detención del ciclo celular en la fase G2/M durante la división de las células somáticas o por daños en el genoma (Preuss y Britt, 2003). En crisantemo esto se atribuye a la generación de aberraciones cromosómicas y pérdida de la capacidad de proliferación de las células (Patil et al., 2015) y a daños en los genes implicados en la morfogénesis de hojas verdaderas, como sucede en hojas de Rumex obtusifolius irradiadas con 500 Gy (Kitano et al., 2015). En Agave tequilana la radiación absorbida produce mutaciones en el ADN de las células y altera su desarrollo normal al aumentar la dosis de radiación (Ángeles-Espino et al., 2013).
Cuadro 4 Correlación del efecto de la radiación gamma 60Co sobre el desarrollo de plántulas de Laelia autumnalis
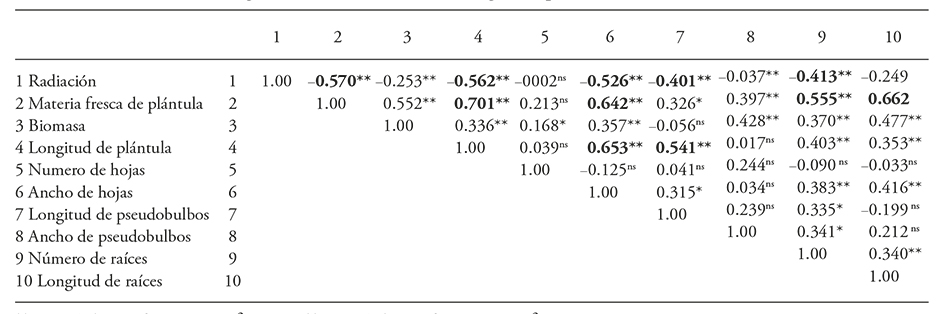
** p≤0.01; * ≤0.05; ns=no significativo.
Hubo coeficientes de correlación positivos entre materia fresca de plántula y longitud de plántula (0.70), longitud de raíz (0.66), anchura de hojas (0.64) y número de raíces (0.55). También hubo coeficientes de correlación significativos entre longitud de plántula y anchura de hojas (0.65) y longitud de pseudobulbo (0.54).
Determinación de la DL50 y GR50
El modelo de regresión lineal mostró el ajuste mejor para explicar el efecto de la radiación sobre la supervivencia de los protocormos (Cuadro 5). La materia fresca de protocormos y de plántulas mostraron una tendencia a disminuir conforme se aumentó la dosis de radiación; sin embargo, presentaron una R2 baja de 0.48 y 0.35 por lo que estas variables no se consideraron para calcular la GR50. La dosis letal media para la supervivencia de protocormos fue 53 Gy (Figura 4), lo cual se considera que favorece la aparición de mutaciones útiles. La dosis de reducción media del crecimiento (GR50) para la formación de hojas en protocormos fue 28 Gy (Cuadro 5 y Figura 4).
Cuadro 5 Análisis de regresión para la supervivencia de protocormos y emergencia de hojas en protocormos de Laelia autumnalis irradiados con rayos gamma 60Co.
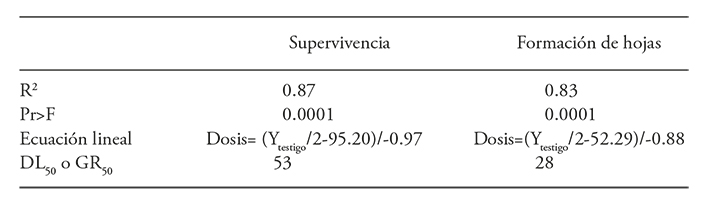
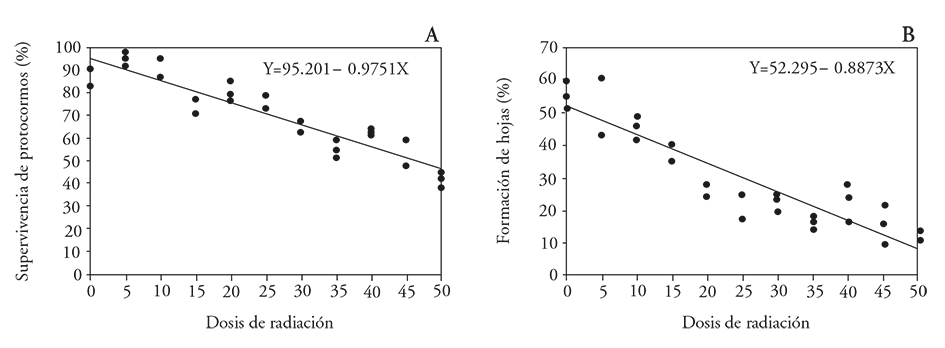
Figura 4 Efecto de la radiación gamma 60Co sobre protocormos de la orquídea Laelia autumnalis, A) porcentaje de supervivencia en protocormos y B) formación de hojas en protocormos.
La DL50 para L. autumnalis difiere a la reportada para rizomas de Alstroemeria aurea G. irradiados con rayos gamma a dosis de 0 a 40 Gy y la DL50 se determinó en 40 Gy; esta dosis redujo la supervivencia de explantes en 50 %, y se puede usar para mejoramiento genético (Aros et al., 2012). La GR50 para el porcentaje de germinación en semillas de dos variedades de arroz (Oryza sativa L.) irradiadas con rayos gamma 60Co de 20 a 200 Gy, fue 89 y 188 Gy en las variedades JGL y Vijetha (Pavan et al., 2013). En brotes de fresa irradiados con 30 a 325 Gy, la DL50 para supervivencia de brotes fue 177 Gy (Murti et al., 2013); en semillas de frijol largo (Vigna sesquipedalis) irradiadas con 300 a 800 Gy la DL50 para supervivencia fue 600 a 800 Gy y la GR50 para altura de planta fue de 400 a 500 Gy (Kon et al., 2007).
Con base en esos datos se confirmó que la DL50 y la GR50 son específicas para cada especie, genotipo e incluso tamaño y tipo de tejido irradiado, y que la sensibilidad a la radiación depende de la naturaleza y extensión del daño cromosómico (Kiong et al., 2008; Pavan et al., 2013). Además varía en función del contenido de ADN, tamaño y número de cromosomas (Sparrow et al., 1961), y el contenido de agua en el material vegetal irradiado, ya que la radiación interactúa con moléculas, especialmente de agua, para producir radicales libres en las células. Estos radicales modifican diferentes compuestos importantes de las células en las plantas, por lo cual las semillas toleran dosis más altas de radiación para llegar a la DL50, en comparación con los tejidos suculentos (Yunus et al., 2013).
La DL50 determinada para las plántulas de L. autumnalis confirma que esta especie es muy sensible a la radiación gamma. Esta sensibilidad también se observa con respecto a la radiación solar, lo que se atribuye a que L. autumnalis es una especie epífita que habita bajo el dosel de los árboles. En las orquídeas epifitas altas intensidades de radiación solar reducen la capacidad fotosintética y afectan el intercambio gaseoso (Stancato et al., 2002).
La dosis reductiva media de 28 Gy para la formación de hojas a partir de protocormos de L. autumnalis es similar a la de Agave fourcroides, en la que se determinó una GR50 de 20 Gy para el peso de materia fresca. Además, el peso de materia fresca de callo es un excelente indicador de la radiosensibilidad (González et al., 2007). En explantes de A. tequilana irradiados con rayos gamma de 0 a 50 Gy, la dosis reductiva media fue 20 a 25 Gy para la formación de plántulas y de 15 a 25 Gy para la formación de callos. Con estas dosis aumenta la probabilidad de inducir mutaciones favorables para fines de selección y mejoramiento genético (Ángeles-Espino et al., 2013). Sin embargo, la GR50 determinada para la formación de hojas a partir de protocormos de L. autumnalis es alta si se compara con los resultados en explantes de Etlingera elatior irradiados con rayos gama 60Co de 0 a 140 Gy, donde la DL50 para supervivencia de explantes se determinó en 10 Gy (Yunus et al., 2013). En tubérculos de nardo (Polianthes tuberosa L.), irradiados con rayos gamma de 0 a 30 Gy la DL50 fue 9.09 Gy para la supervivencia de plántulas aclimatadas procedentes de brotes cultivados in vitro; no así para los tubérculos establecidos in vivo en los que se determinó una DL50 de 25.91 Gy para la supervivencia de plantas (Estrada-Basaldua et al., 2011).
Conclusiones
La radiación gamma 60Co ejerció un efecto inversamente proporcional sobre el crecimiento y desarrollo de plántulas de Laelia autumnalis. Dosis bajas radio estimulan el crecimiento de las plántulas, pero dosis altas inhiben la supervivencia de protocormos, la formación de hojas y la materia fresca de plántulas.
La irradiación con dosis entre 28 y 53 Gy se puede emplear en programas de mejoramiento para L. autumnalis, con el fin de promover mutaciones que favorezcan la obtención de variantes de importancia ornamental.

