











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Entre los sistemas de apareamiento, los diseños dialélicos, definidos como conjuntos de cruzamientos simples posibles entre un grupo de n progenitores, son los más usados en maíz (Zea mays L.). Éstos los presentó Schmidt (1919; citado por Martínez, 1983) y se emplean para estimar las componentes de la variación genética en el rendimiento, otras características de las propias cruzas y su capacidad productiva (Martínez, 1983).
Los cruzamientos dialélicos son herramientas valiosas en el estudio de poblaciones biológicas, para obtener información que permite conocer la acción génica en caracteres cuantitativos de importancia agrícola y para establecer esquemas de mejoramiento en la selección de genotipos superiores (Gardner y Eberhart, 1966; Ávila et al., 2009).
Los estudios de Sprague y Tatum (1942) y Griffing (1956) han contribuido más al uso frecuente de los diseños dialélicos en investigaciones genéticas, especialmente relacionados con el maíz híbrido. En el primer caso, los autores dividieron la aptitud combinatoria (AC) en general (ACG) y específica (ACE). La AC es la capacidad que poseen los individuos o una población de combinarse con otros, la ACG es el comportamiento medio de una línea en combinaciones híbridas y la ACE se refiere al efecto en donde ciertas combinaciones se expresan mejor o peor de lo esperado, con base en el comportamiento promedio de las líneas involucradas. Con respecto al tipo de acción génica que determina la AC de las líneas, la ACG está predominantemente asociada a la aditividad, en tanto que la ACE incluye a los efectos de dominancia y epistasis (Hallauer et al., 2010). Griffing (1956) sistematizó el análisis estadístico para la estimación de los componentes de varianza cuando se utiliza un diseño dialélico basado en ACG y ACE.
El dialélico completo, de interés para nuestro estudio, fue propuesto formalmente por Griffing (1956), quien definió a las cruzas dialélicas como esquemas de apareamiento de un conjunto de p líneas progenitoras, donde se obtiene un máximo de p2 combinaciones, que se dividen en tres grupos: 1) los p progenitores; 2) las p(p-1)/2 cruzas F1 directas; y 3) las p(p-1)/2 cruzas F1 recíprocas. Con estos tres grupos desarrolló cuatro métodos de análisis dialélicos que incluyen: I) las p2 combinaciones posibles; II) los progenitores y las cruzas F1 directas; III) las cruzas F1 directas y recíprocas; y IV) las cruzas F1 directas.
Los diseños de Griffing más usados son el II y el IV. Estos excluyen las cruzas recíprocas y a éstas y a los progenitores, respectivamente. Sin embargo, cuando por alguna razón el investigador supone efectos maternos, es indispensable estudiarlas, para lo que el modelo lineal básico para el análisis de los experimentos dialélicos (Y ijk = µ+g i +gj +sij +rij +eijk) puede desarrollarse aún más: Yijk = µ+gi +gj +sij +mi +mj +rij +eijk; donde Yijk es el valor fenotípico observado de la cruza, con los progenitores i y j, en el bloque k; µ es un efecto común en todas las observaciones; gi y gj son los efectos de ACG de los progenitores i y j; sij es el efecto de ACE de la cruza (i, j); mi y mj representan los efectos maternos de los progenitores (i, j); rij es el efecto recíproco de la cruza (i, j) y eijk es el efecto ambiental aleatorio correspondiente a la observación (i, j, k).
El efecto materno, asociado con ADN extra nuclear del progenitor femenino, con frecuencia es ignorado al emplear el sistema de cruzas dialélicas. Sin embargo, Mahgoub (2011) considera que el fenotipo de un individuo puede estar determinado también por este componente materno. Una forma de identificar tal contribución, de los progenitores a su descendencia, es mediante el estudio de las cruzas recíprocas; de ellos, el dialélico completo es el sistema de apareamiento más común para demostrar este fenómeno (Martínez, 1983).
Con base en lo anterior, el objetivo de este estudio fue determinar la aptitud combinatoria, e identificar los efectos maternos de un grupo de líneas endogámicas (S6-S8) a través de sus cruzas, determinar el tipo de acción genética (dominancia o aditividad) de éstas y valorar su potencial genético, para su aprovechamiento en el programa de mejoramiento MMR (Dr. Moisés Mendoza Rodríguez†) que permita incrementar la producción de grano. La hipótesis plantea la predominancia de los efectos no aditivos sobre los aditivos, así como la existencia de efectos maternos y la expresión de cruzas de comportamiento superior, desarrollados a partir de líneas sobresalientes.
Materiales y Métodos
Las combinaciones posibles entre 10 líneas endogámicas (S6-S8), desarrolladas para la producción de maíz, se evaluaron en condiciones de riego en los Valles Altos del Centro de México. El estudio se realizó durante el ciclo primavera-verano 2014, en tres localidades de esa área ecológica (Cuadro 1).
Cuadro 1 Localidades para la evaluación de cruzas dialélicas.
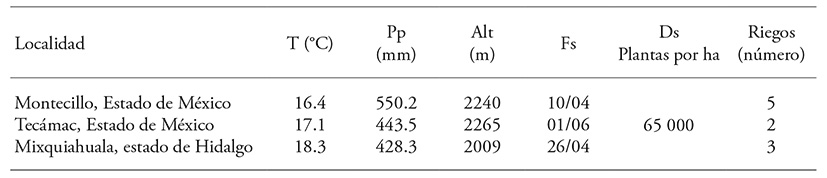
T: temperatura media anual; Pp: precipitación anual; Alt: altura sobre el nivel del mar; Fs: fecha de siembra; Ds: densidad de siembra.
En el ciclo primavera-verano de 2013, las 10 líneas progenitoras de maíz se sembraron en el Campo Agrícola Experimental del Colegio de Postgraduados, Campus Montecillo, las líneas se seleccionaron previamente, en una prueba de mestizos durante 2012; con estos genotipos se desarrolló el esquema de cruzas dialélico; con la semilla obtenida (F1) en 2014 se establecieron los ensayos de evaluación bajo el diseño experimental látice simple 10𝙭10, con cuatro repeticiones, en tres localidades (Cuadro 1); el experimento se desarrolló bajo punta de riego en Tecámac. La unidad experimental fue un surco de 5 m de longitud y 0.8 m de separación.
Durante la estación de crecimiento y en la cosecha se obtuvo información de características cuantitativas asociadas con la fenología, morfología y el rendimiento. Los valores fueron promedio de cinco mazorcas de cada unidad experimental. Los datos se analizaron con correlación de Pearson (Restrepo y González, 2007), los caracteres con correlación positiva mayor con el rendimiento de grano se identificaron. Ellos fueron el peso de las mazorcas primarias (Pmz1), el peso del grano por mazorca (Pgr) y la longitud de la mazorca (Lmz); estos caracteres se analizaron con componentes de la variación.
El rendimiento de grano por unidad experimental (Rto1) se determinó con la ecuación: Rto1=((PMz1+PMz2) X Id X (100 - %Hum)/100)/0.86 donde PMz1 y PMz2 fueron los pesos de las mazorcas primarias y secundarias cosechadas por unidad experimental; Id, el índice de desgrane; % Hum, la humedad de la muestra en campo y 0.86 un factor de corrección, para reajustar el rendimiento a 14 % de humedad (Pérez, 2001). Con este resultado y el tamaño de la unidad experimental se estimó el rendimiento de grano en t ha-1 (Rto).
El análisis genético se realizó con el rendimiento de grano mediante el PROC GLM, y se efectuó con el Modelo I (efectos fijos) del Método I (dialélico completo) de Griffing (1956), considerando que los progenitores fueron seleccionados con base en su comportamiento en una prueba de mestizos.
El análisis de varianza combinado se realizó con el modelo lineal: Yijkl =µ +αl +δ k(l) +νij +υij + (αν) ijl +(αυ)ijl +eijkl donde: Y ijkl es el valor fenotípico observado de la cruza (i, j) dentro de la repetición k en la localidad l; µ es la media general; αl es el efecto de la localidad l; δk(l) es el efecto del bloque o de la repetición k en la localidad l; νij es el efecto de la cruza =gi +gj +sij, donde: gi y g j son los efectos de la ACG de los progenitores i y j, respectivamente; sij es el efecto de ACE de la cruza (i, j); uij =mi -m j +rij, donde: mi y mj son los efectos maternos de los progenitores i y j, correspondientemente y rij es el efecto recíproco de la cruza (i, j); (αν)ijl =(αg)il +(αg) jl +(αs)ijl, donde: (αν)ijl es el efecto de interacción entre las cruzas y el ambiente, (αg)il y (αg)jl es el efecto de interacción entre el efecto de g i y gj y el ambiente, respectivamente, (αs)ijl es el efecto de interacción entre el efecto sij y el ambiente; (αu)ijl =(αm)il +(αm)jl +(αr)ijl, donde (αu)ijl es el efecto de interacción entre υij y el ambiente; (αm)il y (αm)jl son los efectos de interacción entre mi y mj con el ambiente, (αr)ijl es la interacción entre el efecto rij y el ambiente y eijkl es el efecto aleatorio del error correspondiente a la observación (i, j, k, l).
El análisis dialélico se realizó con SAS versión 9.3 (SAS, 2010). La prueba de significancia de los efectos principales (ACG, ACE, EMat, ERec) e interacciones (ACG×Loc, ACE×Loc, EMat×Loc y ERec×Loc) se realizaron con las pruebas de F propuestas por Martínez (1983) para el Diseño I de Griffing.
Para conocer la estabilidad de la expresión de cada genotipo, el análisis se realizó con el valor medio entre las localidades; no obstante, dada la interacción significativa localidades y aptitud combinatoria, se incluyó el análisis específico por localidad.
Resultados y Discusión
Análisis de varianza
Los cuadrados medios del análisis de varianza combinado fueron significativos (p≤0.01) entre cruzas y localidades (Cuadro 2) para rendimiento de grano (Rto) y las variables Pmz1, Pgr y Lmz mostraron correlación mayor con éste. Las diferencias entre las cruzas se atribuyen a la expresión de la variabilidad genética, asociada con los tipos de acción génica que se expresaron en cada una y a la heterosis, producto de la interacción de las líneas participantes.
Cuadro 2 Cuadrados medios del análisis de varianza combinado para el rendimiento de grano y tres variables con mayor correlación con éste en un esquema completo de cruzas dialélicas.
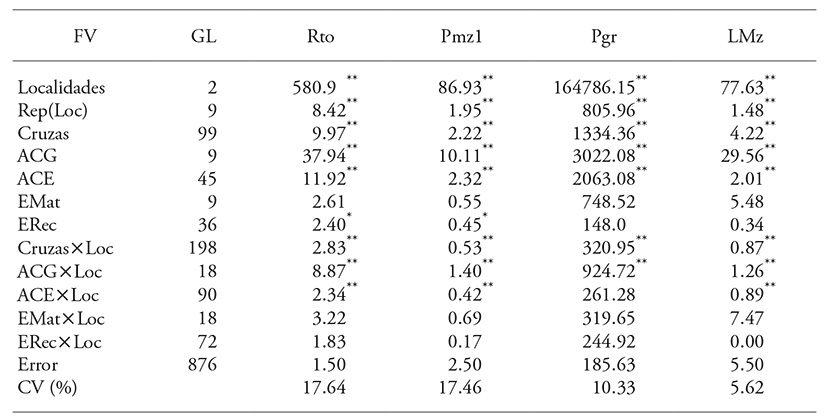
** p≤0.01 y * p≤0.05, FV: fuente de variación, GL: grados de libertad, Rto: rendimiento de grano, Pmz1: peso de mazorcas primarias, Pgr: peso de grano por mazorca, Lmz: longitud de mazorca, ACG: aptitud combinatoria general, ACE: aptitud combinatoria específica, EMat: efectos maternos, ERec: efectos recíprocos, Loc: localidad y CV: coeficiente de variación
La expresión contrastante de las variables entre las localidades se atribuyó al efecto ambiental sobre los genotipos; es decir, a la incapacidad de algunos para amortiguar el efecto ecológico diferencial asociado con las condiciones hídricas, térmicas, fitopatológicas y edáficas. Esto se manifestó en la interacción significativa de cruzas y localidades (Cruzas×Loc). Al respecto, Hallauer et al. (2010) señalan que las cruzas simples, comparadas con las cruzas trilineales y dobles, generalmente presentan mayor interacción con el ambiente, dada su homogeneidad.
Las diferencias entre las cruzas fueron significativas por lo cual, la suma de cuadrados fue fraccionada entre ACG y EMat de los progenitores y ACE y ERec de los cruzamientos (Cuadro 2). Para ACG y ACE se encontraron diferencias significativas (p≤0.01) en todas las variables, lo cual indica contrastes debidos a efectos aditivos y no aditivos.
Los EMat no fueron significativos (p>0.05), es decir, los caracteres Rto, Pmz1, Pgr y Lmz expresaron sólo herencia nuclear y no mostraron efectos significativos debidos a información genética extra nuclear. Los ERec presentaron diferencias significativas (p≤0.05) en Rto y Pmz1, las que se atribuyen a los efectos de interacción entre el ADN nuclear y el citoplásmico. Estos los denominó Cockerham (1963) efectos no maternos.
La interacción significativa Cruzas×Loc condicionó el fraccionamiento de los efectos de interacción: ACG×Loc, ACE×Loc, EMat×Loc y ERec×Loc (Cuadro 2). Las interacciones ACG×Loc y ACE×Loc fueron significativas (p≤0.01), excepto en la variable Pgr y ERec×Loc; las interacciones Emat×Loc y Erec×Loc no fueron significativas. Las interacciones mostraron que los efectos de ACG y ACE fueron específicos para los ambientes de evaluación; por lo tanto, se incluyó el análisis en cada localidad.
La suma de cuadrados del ANAVA indicó que la contribución a la varianza del rendimiento de grano, atribuible a las cruzas, estuvo explicada por ACE 54 %, ACG 34 %, ERec 8.7 % y EMat 2.3 %; es decir, para Rto los efectos no aditivos fueron más importantes que los aditivos, aunque estos últimos fueron también significativos en la expresión de la variación genética del rendimiento. Una tendencia similar se observó para Pgr. Según Ávila et al. (2009) la superioridad de los efectos no aditivos se puede deber a la heterosis resultante de las combinaciones híbridas entre los progenitores, la cual pudo asociarse con divergencia genética entre las líneas (Legesse et al., 2009). Para Pmz1 los efectos de ACG y ACE fueron similares, mientras que para Lmz el efecto de ACG fue mayor que el de ACE; la contribución a la varianza de ERec fue limitada (<6 %). Estudios similares también muestran que el rendimiento de grano está controlado principalmente por efectos no aditivos (Ávila et al., 2009; Pérez-López et al., 2014; Rodríguez-Pérez et al., 2016).
Los resultados mostraron que los efectos de ACG, ACE y ERec difirieron con respecto al carácter evaluado y al genotipo estudiado. No obstante, son deseables aquellas líneas cuya ACG para el rendimiento de grano esté correlacionada con la ACG para componentes del rendimiento (Fan et al., 2008).
Los efectos maternos (EMat) no significativos en este estudio mostraron que el aprovechamiento de las cruzas podría realizarse en ambos sentidos (directo y recíproco), sin que haya diferencias significativas. Esto se apoya en lo señalado por Cockerham (1963) y Hallauer et al. (2010), respecto a que en plantas los efectos maternos son mínimos y que generalmente es innecesaria su consideración. Sin embargo, podrían haber interpretaciones erróneas y generar incertidumbre de los resultados si no se hubieran evaluado (Mahgoub, 2011). Al contrario, Fan et al. (2013) consideran los efectos maternos importantes para cultivos como el maíz, donde el rendimiento de grano está determinado ampliamente por el endospermo.
Rendimiento de grano y ACG de las líneas
La expresión fenotípica de las líneas fue sobresaliente, ya que aun cuando son altamente homocigóticas (S6-S8) mostraron un valor medio de rendimiento superior al mínimo necesario (5.6 t ha-1, valor promedio general; Cuadro 3). De acuerdo con Espinosa et al. (2002), para la rentabilidad económica de la producción de semilla, las líneas utilizadas como progenitores femeninos, de híbridos de cruza simple, deben expresar una producción mínima necesaria de 3.0 t ha-1.
Cuadro 3 Rendimiento de grano y efecto de aptitud combinatoria general de las líneas progenitoras en tres localidades de los Valles Altos del Centro de México.
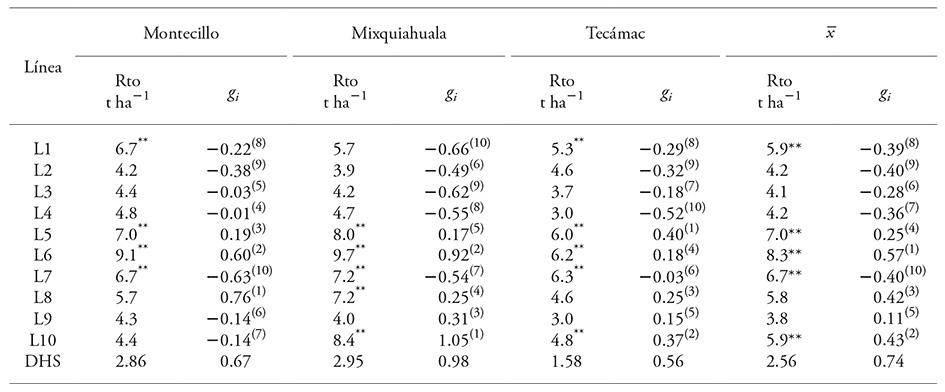
** p≤0.01 y * p≤0.05; Rto: rendimiento de grano, g i : efecto de ACG y DHS: diferencia honesta significativa. El valor en paréntesis muestra el orden de las líneas con respecto al efecto de su ACG (1: mayor y 10: menor)
Con base en el valor medio de las tres localidades, el rendimiento de las líneas mostró un intervalo de variación de 8.3 a 3.8 t ha-1. En particular, el rendimiento mostró potencial mayor en Mixquiahuala, Hidalgo (6.3 t ha-1 en promedio) y menor en Tecámac, Estado de México (4.8 t ha-1 en promedio).
El comportamiento anterior se asoció con particularidades de las localidades. En Mixquiahuala, aunque con condiciones hídricas menores a las comunes, dada la reducción de 6 a 3 riegos (con “aguas negras”) por la disponibilidad limitada de agua en el distrito de riego, suministrados en los momentos oportunos (en siembra, durante la floración y en el llenado de grano), se obtuvo la mejor expresión de los genotipos (líneas y cruzas simples). En Montecillo, la incidencia “atípica” de Fusarium spp. en el lote experimental, causó reducción significativa en el rendimiento (20 %), inesperada porque el rendimiento podría haberse igualado al de Mixquiahuala, por las condiciones óptimas para el cultivo. En Tecámac el sistema de riego estuvo afectado, por lo que la aplicación de dos riegos únicamente, en la siembra y en la floración, causó restricción hídrica y provocó rendimiento de grano menor, respecto al resto de las localidades.
En Mixquiahuala las líneas mostraron variación de Rto, de 9.7 (L6) a 3.9 (L2) t ha-1 (Cuadro 3); en Montecillo el intervalo fue 9.1 (L6) a 4.2 (L2) t ha-1 y en Tecámac 6.3 (L7) a 3.8 (L9) t ha-1. A pesar del rendimiento satisfactorio de las líneas, fue posible discriminar su expresión e identificar a las superiores. Acorde al valor medio de las tres localidades, las líneas L5, L6 y L7 mantuvieron un Rto igual o superior a 6.0 t ha-1 y el de L2, L3, L4 y L9 fue menor a 5 t ha-1.
Para ACG, dada la interacción con localidades, el comportamiento en cada sitio se analizó. En Montecillo (Cuadro 3) las líneas L6, L5, L1 y L7 mostraron Rto superior (9.1, 7.0, 6.7 y 6.7 t ha-1, respectivamente) y solo L6 y L5 fueron sobresalientes en Rto y ACG. La línea L8 con Rto de 5.7 t ha-1 presentó la ACG mayor (0.76). Por lo tanto, las líneas L5, L6 y L8 de efectos positivos se consideraron de ACG superior.
En Mixquiahuala (Cuadro 3) las líneas L6, L10, L5, L7 y L8 presentaron Rto mayor (9.7, 8.4, 8.0, 7.2 y 7.2 t ha-1). De acuerdo con Rto y los efectos de ACG, para esta localidad se determinó que las líneas de ACG alta fueron L5, L6, L8, L9 y L10.
En Tecámac (Cuadro 3), las líneas de Rto mayor fueron: L7, L6, L5, L1 y L10; pero sólo tres (L5, L6 y L10) presentaron coeficientes de ACG positivos, a las cuales se adhirieron L8 y L9 como líneas de ACG alta, pero de rendimiento medio.
De acuerdo con lo anterior y el valor medio de Rto y ACG fue posible diferenciar y clasificar el comportamiento de las líneas en: 1) ACG alta y Rto superior L5 y L6, 2) ACG alta y Rto medio L8 y L10, 3) ACG positiva y Rto bajo L9, 4) ACG baja y Rto medio L1 y L7, y 5) ACG baja y Rto bajo L2, L3 y L4.
La clasificación de las líneas con base en su ACG y Rto fue adecuada porque se espera (Reyes et al., 2004; Legesse et al., 2009; Escorcia-Gutiérrez et al., 2010; Pérez-López et al., 2014) que las de ACG alta formen cruzas con rendimiento superior, por la probabilidad mayor de interaccionar positivamente entre ellas debido a la contribución de sus alelos favorables. Al contrario, las líneas de ACG baja se esperaría que apareadas entre sí originaran cruzamientos poco sobresalientes.
Rto y ACG no se relacionaron directamente, pues en Montecillo L1 y L7 presentaron Rto alto y g i negativo y L8 Rto medio pero g i positivo. Es decir, entre las líneas con Rto mayor solo algunas tuvieron los efectos de ACG mayores. Estos resultados contrastaron parcialmente con los de Reyes et al. (2004) quienes encontraron que las líneas endogámicas con Rto mayor fueron también las de ACG alta. En contraste, los resultados coincidieron con los de Pérez-López (2014), quienes asociaron tal respuesta a la selección integral de las líneas, que juntó con el criterio de rendimiento superior se seleccionaron genotipos con características complementarias importantes para el fitomejoramiento, como precocidad, altura de la planta, peso de la mazorca, número de hileras, lo que pudo conducir a las discrepancias entre Rto y ACG.
Rendimiento de grano y ACE de las cruzas
Aunque, la interacción ACE×Loc fue significativa (Cuadro 2), el análisis de la expresión del rendimiento de las cruzas y su ACE se hizo con base en el valor medio de las tres localidades, esto permitió reconocer la estabilidad o adaptación específica de las cruzas y el efecto de la ACE.
Por localidad, el valor medio de rendimiento de las 90 cruzas posibles, siguió la misma tendencia que en las líneas: Mixquiahuala (8.5 t ha-1) >Montecillo (6.8 t ha-1) y Tecámac (6.0 t ha-1). Este comportamiento fue dependiente de las condiciones ambientales de las localidades (Cuadro 4).
Cuadro 4 Efecto de aptitud combinatoria específica en veintidós cruzamientos con mayor y cinco con rendimiento menor de grano en cada localidad de estudio y el valor medio entre las mismas.
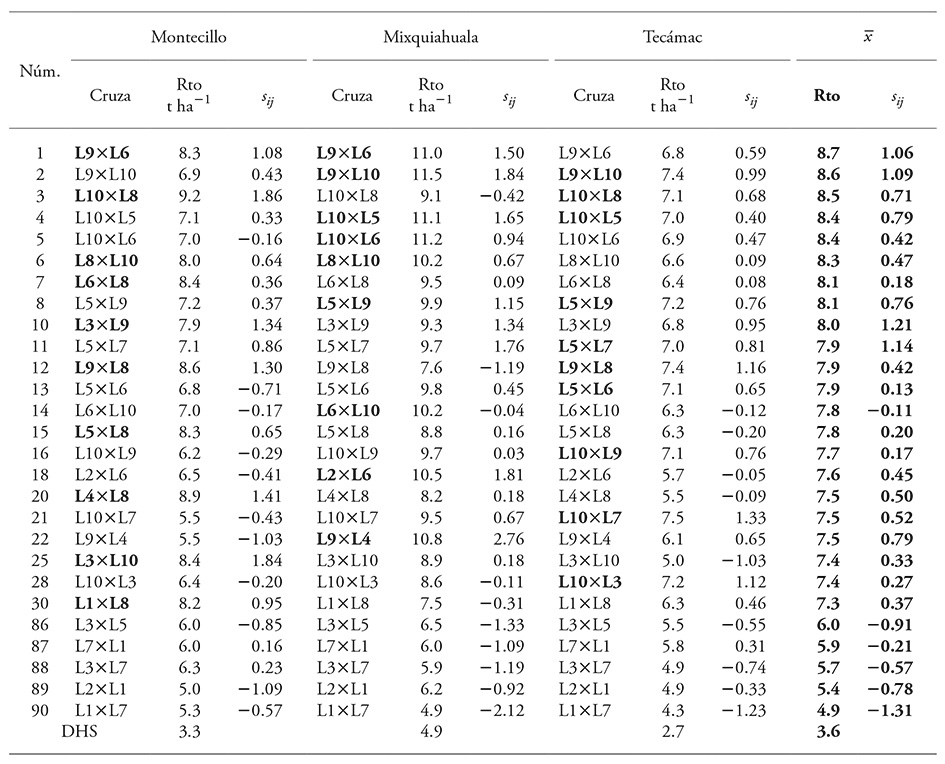
Rto: rendimiento de grano, sij: efecto de ACE, DHS: diferencia honesta significativa, Núm: orden ascendente de los genotipos (1-90) con base en el valor medio de rendimiento de las tres localidades estudiadas. En negritas, las diez cruzas de rendimiento superior en cada localidad
En general y con base en el valor medio, el rendimiento de grano de las 22 cruzas con rendimiento sobresaliente en el estudio, que incluyeron las diez cruzas de rendimiento superior en cada localidad, fue entre 8.7 y 7.3 t ha-1. Al contrario, aquélla con rendimiento inferior mostró 4.9 t ha-1 (Cuadro 4).
Los valores de ACE fueron predominantemente positivos para las cruzas de rendimiento superior y negativos para las de rendimiento inferior, esta expresión coincidió con estudios de aptitud combinatoria similares (Reyes et al., 2004; Pérez-López et al., 2014). El intervalo de variación de la ACE fue de 1.21 a -1.31, que en el valor positivo no coincidió con la expresión de Rto superior pero sí en el caso negativo con el Rto inferior. No obstante, a pesar de la variación para este efecto, los valores fueron pequeños comparados con los que obtuvieron Reyes et al. (2004) y Escorcia-Gutiérrez et al., (2010), pero similares a los de Legesse et al. (2009). Esto pudo deberse a la divergencia genética entre las líneas, que también se observó mediante la comparación del rendimiento de las mejores líneas y de sus cruzas respectivas.
De acuerdo con el valor medio de rendimiento, las ocho cruzas superiores (>8.0 t ha-1) sobresalen por presentar Rto superior en al menos dos localidades; ninguna cruza con rendimiento entre las diez cruzas superiores en cada localidad. Entre la ACE y el Rto no se observó relación directa, pues las cruzas L9×L6 y L9×L10, con Rto superior, mostraron valores positivos y altos; pero L3×L9 y L5×L7, con Rto medio, mostraron valores ACE mayores. Al contrario, L6×L10 con Rto medio mostró ACE negativa.
Con base en lo anterior, los valores de ACE no permitieron explicar completamente la expresión del rendimiento; pero se observó una tendencia asociada con ACG de las líneas para las cruzas. De las noventa cruzas posibles, 22 fueron sobresalientes (Cuadro 4); de estas, ocho con Rto superior (>8.0 t ha-1) se formaron solo por líneas de ACG alta (L5, L6, L8 y L10) o al menos positiva (L9). En el resto de las cruzas sobresalientes (<8.0 t ha-1), en la mayoría participó al menos una línea de ACG baja, aunque también hubo algunas en las que participaron dos líneas de ACG alta, como L9×L8, L5×L8), e incluso las dos líneas con Rto mayor y ACG alta, L5×L6.
La expresión genética del rendimiento de grano, para las cruzas sobresalientes estuvo fundamentado en dos posibilidades: 1) que la cruza esté formada por ambas líneas de ACG alta, en cuyo caso los efectos de ACE pueden ser altos o no, y 2) que la cruza se forme con al menos una línea de ACG alta pero cuyos efectos de ACE sean altos, al menos en una localidad. Este resultado coincidió parcialmente con los de Reyes et al. (2004) y Perez-López et al. (2014) y forma parte de la variación genética en la expresión de los caracteres del maíz, donde también son posibles rendimientos satisfactorios a partir del cruzamiento de líneas con ACG baja, como lo mostraron Rodríguez-Pérez et al. (2016).
En cuanto al rendimiento inferior, los efectos de ACE fueron negativos, y las cuatro cruzas inferiores estuvieron formadas por líneas de ACG baja: L1, L2, L3 y L7. También se encontró una donde participó una línea de ACG alta: L3×L5.
De acuerdo con la expresión del rendimiento de los 22 cruzamientos sobresalientes se identificó y clasificó la ACG de las líneas con base en el número de cruzas sobresalientes. Así, la línea L10 participó casi en 50 % (10 cruzas), le siguieron L9, L8 y L6 que, con excepción de la última (con Rto alto), fueron clasificadas previamente como de ACG alta (L8 y L10) o positiva (L9) pero con rendimiento medio o bajo. Además, considerando la expresión del rendimiento de la cruza L5×L6, cuyas líneas fueron de ACG alta y Rto alto, se determinó que el rendimiento per se de una línea no fue indicativo de su ACG y que tampoco garantiza que mediante su cruzamiento se obtenga expresión superior de rendimiento.
Los efectos maternos limitados dentro del grupo de líneas se confirmó (Cuadro 4), pues dentro de las cruzas existieron al menos cinco pares con rendimiento similar. Es decir, tanto la cruza directa y recíproca mostraron expresiones equivalentes, como L9×L10 y L10×L9, L10×L8 y L8×L10, L1×L7 y L7×L1.
Estructura genética de las cruzas
La estructura genética de una cruza permite conocer los tipos de acción génica que en ella operan; si en un genotipo los efectos de aditividad son más importantes que los efectos no aditivos (dominancia y epistasis), se recomienda hacer mejoramiento por selección recurrente. Al contrario, si los efectos no aditivos son predominantes, la hibridación es la mejor opción (Rodríguez-Pérez et al., 2016). Con base en lo anterior se realizó el siguiente análisis, con las diez cruzas con la media de rendimiento superior entre localidades y aquélla con la expresión inferior, de las 90 posibles (Cuadro 5). En este análisis no se consideraron los efectos maternos y recíprocos, por su aportación baja a la variación del rendimiento.
Cuadro 5 Estructura genética de diez cruzas con rendimiento de grano mayor y la del menor, con el valor medio de la evaluación en tres localidades de los Valles Altos del Centro de México.
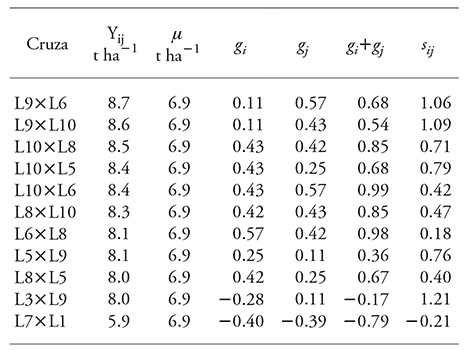
Y ij : valor fenotípico observado de la cruza (i, j), µ: media general, g i y g j : efecto de la ACG del progenitor i y j, respectivamente, s ij : efecto de ACE de la cruza (i, j).
Con excepción de L3×L9, los efectos de ACG y ACE fueron positivos para las cruzas de rendimiento superior, y para la cruza con rendimiento inferior todos los efectos fueron negativos.
La estructura genética de las cruzas mostró comportamientos contrastantes lo cual resaltó la posibilidad de emplear específicamente algunos genotipos. Las cruzas L9×L6 y L9×L10, L5×L9 y L3×L9, donde s ij >g i +g j , en las que los efectos aditivos fueron menores que los no aditivos, sólo pueden usarse como híbridos, porque se esperarían efectos de depresión endogámica en las generaciones filiales, que es una características indeseables en la producción de maíz.
Las cruzas L10×L8 y L10×L5, donde g i +g j =s ij , pueden emplearse como híbridos o incorporarse a un proceso de selección recurrente. Las cruzas L10×L6, L8×L10, L6×L8 y L8×L5, donde g i +g j >s ij , en las que los efectos aditivos fueron mayores que los de dominancia, pueden aprovecharse como variedades de polinización libre, sin detrimento del rendimiento en generaciones avanzadas.
Los resultados de la estructura genética coincidieron con la explicación de la suma de cuadrados del análisis de varianza, la cual indicó que la contribución a la varianza del rendimiento de grano estuvo principalmente determinada por efectos no aditivos. Sin embargo, los efectos aditivos no fueron despreciables.
Conclusiones
La predominancia del efecto no aditivo (dominancia y epistasis) sobre el aditivo en el sistema completo de cruzas permitió definir que el aprovechamiento de los progenitores a través del sistema de hibridación es el más indicado para el programa de mejoramiento genético.
Las líneas no mostraron efectos maternos (extranucleares); por lo que, pueden aprovecharse para beneficiar la producción de semilla de las cruzas sobresalientes, utilizándose indistintamente como hembras o machos.
Las líneas y las cruzas con rendimiento superior mostraron potencial de aprovechamiento en la producción de grano para las localidades de evaluación ubicadas en los Valles Altos del Centro de México.

