










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgrociencia
versão On-line ISSN 2521-9766versão impressa ISSN 1405-3195
Agrociencia vol.51 no.3 Texcoco Abr./Mai. 2017
Crop Science
Influence of woody species in the dynamics of weeds in agroforestry systems in Yucatan, Mexico
1Campus de Ciencias Biológicas y Agropecuarias, Universidad Autónoma de Yucatán, México.
2Instituto Tecnológico de la Zona Maya, Quintana Roo, México. (fkzanov@gmail.com)
3Instituto Tecnológico de Chiná, Campeche, México.
The factors that limit maize production in Yucatán, México are decreasing yields and a constant increase of weedy species. A strategy to deal with these problems is to develop diversified productive schemes, such as agroforestry systems (SAF). This study determined the cover dynamics and biomass of weeds in an SAF composed of forage trees and maize during two cropping cycles (CC). Leucaena leucocephala, Guazuma ulmifolia, and Moringa oleifera, associated with maize, and a plot of maize monocrop were evaluated with a complete random block design with three replications. Weed cover, biomass, diversity and density were measured at the beginning, middle and end of the two CC. The weeds were collected in squares delimited by 50x50 cm frames distributed randomly in each experimental unit, separated by species and dried. With this data, an ANOVA was performed, and when significance was detected, the Tukey test (p≤0.05) was used. At the beginning of the first CC, the SAF Guazuma + maize had lower weed cover (41%), while Moringa + maize was higher at the middle and end (71 and 49%). At the beginning of the second CC, the Guazuma + maize SAF had less weed cover, and Guazuma + maize and Leucaena + maize were lower (54 and 48%). In both CC, weed biomass in the SAF was almost half that of monocropped maize. At the end of both CC, the Guazuma + maize system had less weed diversity than the maize monocrop. At the beginning and end of the two CC, weed density was greater in the monocrop. Eighteen botanical families were identified and, of these, 21% were leguminous. The suppression of weeds in the SAF was 23 to 36%. Woody species in SAF reduce weed cover and biomass, relative to maize monocrop.
Key words: Weeds; agroforestry systems; cover; biomass; suppression
Los factores limitantes para la producción de maíz en Yucatán, México son la reducción en el rendimiento y aumento constante de especies arvenses. Una estrategia para enfrentar esos problemas es desarrollar esquemas productivos diversificados, como los sistemas agroforestales (SAF). En nuestro estudio se determinó la dinámica de la cobertura y biomasa de arvenses en un SAF integrado por árboles forrajeros y maíz durante dos ciclos de cultivo (CC). Para ello se evaluaron Leucaena leucocephala, Guazuma ulmifolia, y Moringa oleifera asociadas a maíz, y un monocultivo de esta especie en una parcela, con un diseño de bloques completos al azar y tres repeticiones. La cobertura, biomasa, riqueza y densidad de arvenses se midieron al inicio, a la mitad y al final de dos CC. Las arvenses recolectadas estaban en cuadros de 50x50 cm, distribuidas aleatoriamente en las unidades experimentales; luego, se separaron por especie y secaron. Con los datos se usó un ANDEVA y cuando hubo significancia se realizó la prueba de Tukey (p≤0.05). Al inicio del primer CC, el SAF Guazuma + maíz tuvo menor (41%) cobertura de arvenses y Moringa + maíz fue mayor a la mitad y al final (71 y 49%). Al inicio del segundo CC el sistema Guazuma + maíz tuvo menor cobertura de arvenses y al final, Guazuma + maíz y Leucaena + maíz fueron menores (54 y 48%). En ambos CC la biomasa de arvenses en los SAF fue casi la mitad que en maíz, en ambos CC la biomasa de arvenses en los SAF fue casi la mitad que en maíz en monocultivo. Al final de ambos CC el sistema Guazuma + maíz tuvo menor riqueza que en monocultivo de maíz. En los dos CC, al inicio y al final la densidad de arvenses fue mayor en el monocultivo. Dieciocho familias botánicas se identificaron y 21% de ellas son leguminosas. La supresión de arvenses en los SAF fue 23 a 36%. Las especies leñosas en SAF reducen la cobertura y biomasa de arvenses en comparación con el maíz en monocultivo.
Palabras clave: Arvenses; sistemas agroforestales; cobertura; biomasa; supresión
Introduction
The milpa, or slash-and-burn system, is a diversified productive scheme practiced on more than 30% of the cultivated area of Yucatán, Mexico (Bellon et al., 2011). However, recently, the bases that made it sustainable have been lost (Caamal-Maldonado et al., 2001) because agricultural policy has established that land use should be intensive and crops changed (Pacini et al., 2003). These changes can lead to soil degradation, with decrease in crop yield and weed increase that grow in areas disturbed by agricultural management (Caamal-Maldonado et al., 2001; Charudattan 2001). These are the two main factors that limit maize production in Yucatán (Latournerie et al., 2006).
Despite farmers’ efforts to control weeds manually and chemically, they persist as a problem for maize production. For this reason, new options were developed, such as using live, or dead, cover crops (mantillas), particularly legumes, or interspersing bush or tree species (Caamal-Maldonado et al., 2001; Bainard et al., 2013). These species can keep moisture, add organic matter, improve nutrient cycling and fix nitrogen (Segura-Rosel et al., 2012; Bainard et al., 2013).
Agroforestry systems that integrate woody species (leguminous and non-leguminous) with traditional crops or animals, or both, are another productive, environment-friendly option (Casanova-Lugo et al., 2014) that can contribute to weed control (Caamal-Maldonado et al., 2001). In this regard, Casanova-Lugo et al. (2010) mention that tree species can produce a large amount of foliage and limit weed growth because of the shade they provide. In fact, because they have greater biomass, tree species prevent weed species from having the space, light, moisture or nutrients they need for growth. Moreover, some woody plants can have allelopathic effects that inhibit development of herbaceous species (Rippin et al., 1994; Lin et al., 2004; Cummings et al., 2012).
Balandier et al. (2006) point out that tree species that have rounded, extended crowns as well as simple oval leaves, such as Guazuma ulmifolia, can reduce diversity and abundance of weeds under their canopy because they intercept a large quantity of light. The objective of our study was to determine the dynamics of cover and biomass of weeds in an agroforestry system integrated by forage trees and maize during two cropping cycles.
Materials and Methods
Study site
The study was conducted on the Campus of Biological and Agricultural Sciences of the Universidad Autónoma de Yucatán (21° 51’ N y 89° 41’ W), in Xmatkuil, Yucatán, Mexico (Figure 1), from January to April 2012. The region’s climate is Aw0 (García, 1988); in the last two years average annual rainfall was 967 mm, most concentrated in the period from June to October. Average high and low temperatures are 37.8 and 24.5 °C (Figure 2). Predominant soils are moderately fertile rendzinas with 1 to 1.5% organic carbon (CO) and pH 7.5 to 7.8 (Bautista et al., 2011).
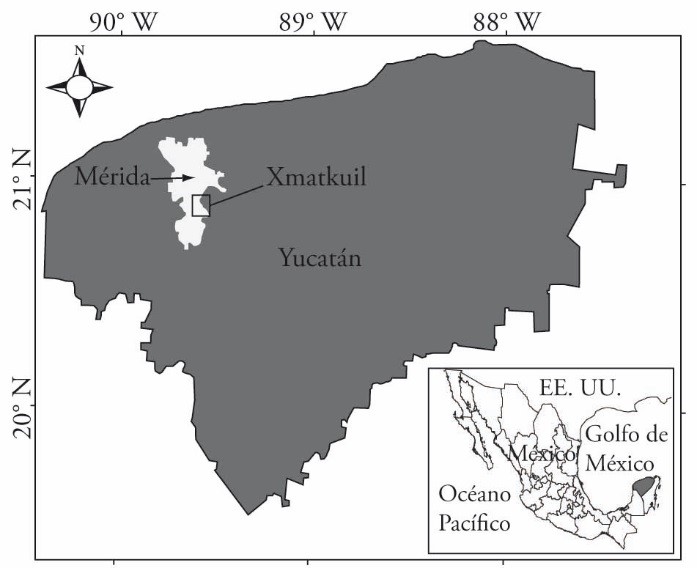
Fuente: (http://www.seduma.yucatan.gob.mx/ordenamiento-ecologico/index.php).
Figure 1 Location of the study site in Xmatkuil, Yucatán, Mexico.
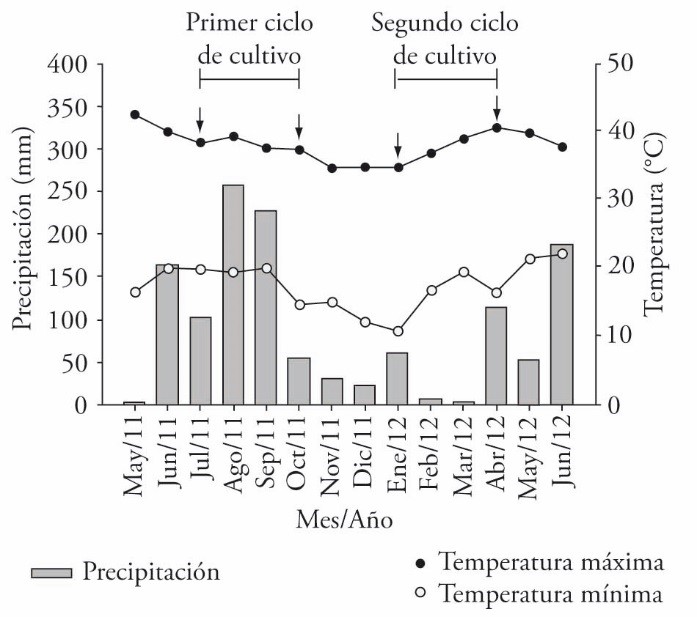
Figure 2 High and low temperatures and rainfall in the study site. Arrows indicate harvest dates of forage species biomass. (Data were taken from the mini-weather station of the Campus of Biological and Agricultural Sciences of the Universidad Autónoma de Yucatán, Mexico, from May 2011 to June 2012).
The area’s vegetation is classified as low deciduous tropical forest, in which bushes reach heights o 4 to 7 m. The most abundant species are Gymnopodium floribundum, Sapranthus campechianus, Bourreria pulchra, Diospyros bumeloides, Neomillspaughia emarginata, Gouania lupuloides, Karwinskia humboldtiana, Exostema caribaeum, Guettarda elliptica, Hamelia patens, Machaonia lindeniana, Randia aculeata, R. longiloba, Thouinia paucidentata, Sideroxylon americanum, Celtis iguanaea, Callicarpa acuminata, Petrea volubilis, and Hybanthus yucatanensis. In addition, there are larger species (8 and 12 m tall) such as Bursera simaruba, Havardia albicans, Lysiloma latisiliquum, Piscidia piscipula and Alvaradoa amorphoides (Gutiérrez-Báez and Zamora-Crescencio, 2012).
Experimental plot
The experimental plot was established in 2004 and experimental units were located; each plot had a woody species: L. leucocephala, G. ulmifolia or M. oleifera. The woody plants were established in rows (three per experimental unit): 2.0 m between rows and 0.5 m between plants. Trees were completely pruned (>90% of the foliage) to 1.0 m above ground level with a use frequency of three months, from the first year of establishment. Auxiliary irrigation was applied during the dry season.
For this study, two crop cycles (CC) were evaluated. In both cycles, maize planting density was 20,000 plants ha-1, using seeds of the local variety “Nal-Xoy”, a random cross of “Nal-Tel” with the variety PR7822 (Graefe, 2003). This maize cross has been used since establishment of the experimental plot. Seeds were germinated in plastic trays with 70 wells 3 cm in diameter and 5 cm deep. The seedlings were transplanted 15 d after germination. Before transplanting the maize, the woody species were pruned, as was done at establishment of the experimental plot (Caamal-Maldonado et al., 2012).
The experimental design was complete random blocks with three replications. The treatments were the following: L. leucocephala associated with maize (Leucaena + maize), G. ulmifolia associated with maize (Guazuma + maize), M. oleifera associated with maize (Moringa + maize), and a control (maize monocrop).
Weed sampling
To understand weed dynamics, samples were taken at three times: 1) at the beginning of each CC (July 2011 and January 2012) when maize was transplanted; 2) at the middle of each CC after the first pruning of forage tree species (August 2011 and March 2012); and 3) at the end of each CC before pruning forage tree species (October 2011 and April 2012).
Response variables
Total weed cover was estimated in 0.5x0.5 m metal frames, six per experimental unit distributed in zig-zag on both sides of the center row of the respective tree species, using high-resolution digital photographs, 14.0 megapixels, taken with a Canon Rebel EOS XS camera. These images were taken at a perpendicular angle (90°) at a height of 1.20 m. Each image was then analyzed with the software CobCal version 2.1 for Windows© to estimate cover (Ferrari et al., 2006).
The weeds inside the metal frame were harvested with roots to determine fresh weight of the total biomass. They were separated by species to determine their diversity (number of species) and counted to quantify their density (individuals m-2). All samples were dried in a forced air circulation oven at 70 °C until constant weight to determine dry matter content. Weed suppression potential was also determined with the formula of Rippin et al. (1994):
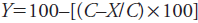 (1)
(1)
where C is weed biomass in the control treatment (g DM m-2), and X is equal to the weed biomass in the treatment corresponding to the agroforestry systems (g DM m-2).
Statistical analysis
The data were analyzed with an ANOVA using GLM (SAS Institute, 2004) to examine the effect of agroforestry systems on the cover dynamics, biomass, diversity and density of the weeds in these systems (SAS Institute, 2004). For significant differences, the Tukey test (p≤0.05) was used.
Results and Discussion
Weed cover
At the beginning of the first CC the Guazuma + maize system had less weed cover (41%), relative to the other treatments. At the middle of this CC (August) the Leucaena + maize system had less weed cover, followed by the plot with Guazuma + maize and the maize monocrop (control). The latter two were similar to the system with Moringa + maíz, which had more weed cover (71%). At the end of this CC, the systems Leucaena + maize, Guazuma + maize, and the monocrop continued to have the lowest weed cover (49%) (Table 1).
Table 1 Weed cover (m2) in agroforestry systems formed by forage trees and maize compared with a maize monocrop in two crop cycles, Xmatkuil, Yucatán, México.
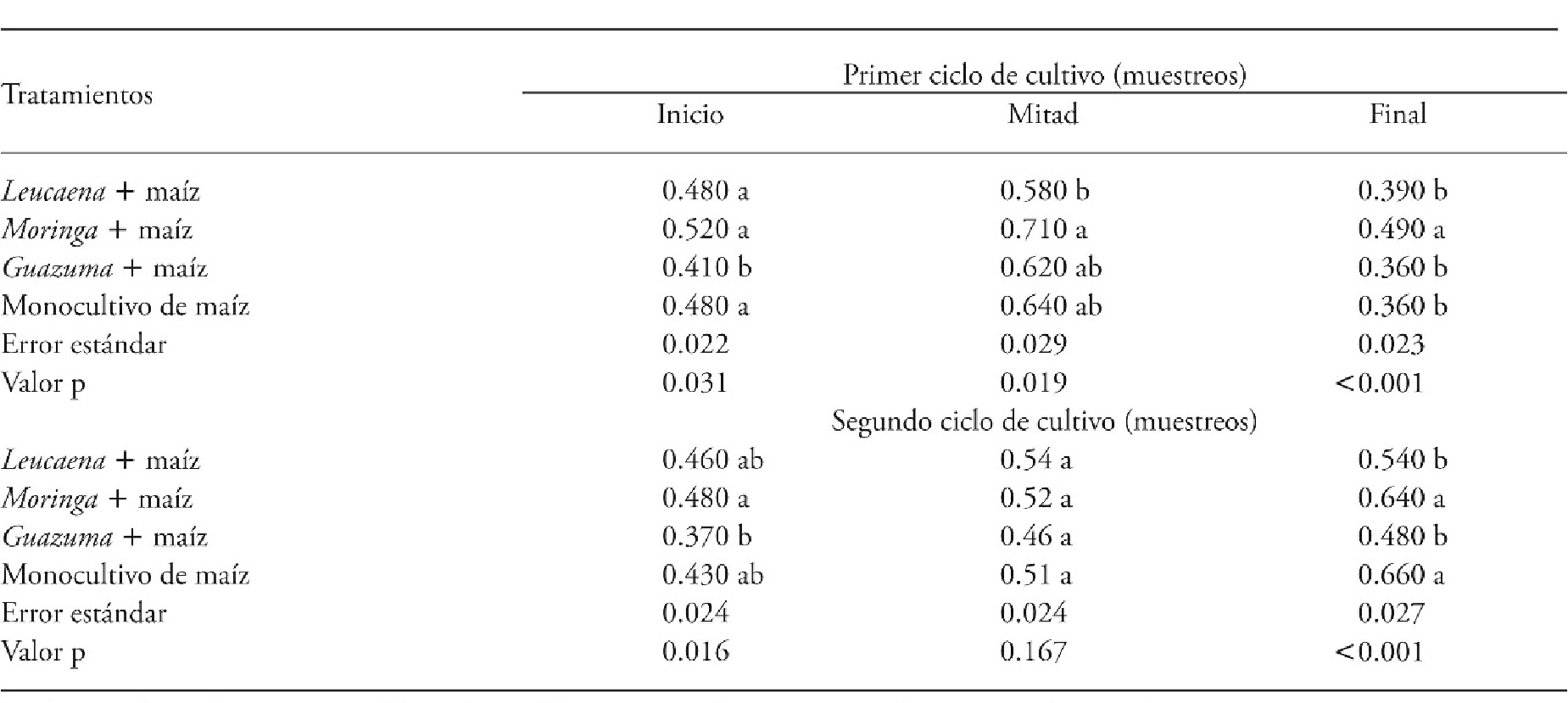
Means in a row with different letters indicate significant differents for each sampling period (p≤0.05).
At the beginning of the second CC the Guazuma + maize system had the least weed cover (37%) followed by Leucaena + maize and the maize monocrop. The latter two were not different from the system with Moringa + maize. At the middle of this cycle, weed cover was similar in all treatments (51% on average), but at the end of the second cycle, Guazuma + maize and Leucaena + maize had less weed cover (48 and 54%), (Table 1).
In our study, the association of maize with woody species showed positive results for weed control. This agrees with that reported by Rippin et al. (1994), Caamal-Maldonado et al. (2001) and Cummings et al. (2012). This is true particularly in reference to biomass. Even though the trees were pruned at the beginning of each cycle, later branch development provided shade, especially in Guazuma. Moreover, its continuous loss of leaves created permanent cover on the soil, forming a kind of mulch, which limited weed growth. (Caamal-Maldonado et al., 2001). In this way, crop associations with leguminous and non-leguminous species can contribute to reducing weed biomass.
It is important to underline that the methods for measuring cover differ among the studies reviewed; in some cover is estimated using indirect (visual) measurements but in our study digital photographs were used because they offer greater precision in measuring this variable (Ferrari et al., 2006).
Although we used a more precise photographic method to assess cover, the results did not show a direct relationship to weed biomass. In fact, the percentage of cover does not permit discriminating whether there is more or less biomass; it only expresses whether the surface is more or less occupied by weeds. Thus, there may be sampling frames with more or less biomass that have the same cover. Indeed, species with horizontal growth habits (i.e. creepers, procumbent or ascendant) will cover more ground than species with vertical growth (i.e. straight). Likewise, the shape and angle of the leaves are fundamental elements of this variable (Rippin et al., 1994).
Weed biomass
During the first and second CC, the agroforestry systems (i.e. Guazuma + maize, Moringa + maize and Leucaena + maize) had less weed biomass than the maize monocrop (Table 2).
Table 2 Weed biomass (g DM m-2) in agroforestry systems consisting of forage trees and maize compared with a maize monocrop in two crop cycles, Xmatkuil, Yucatán, México.
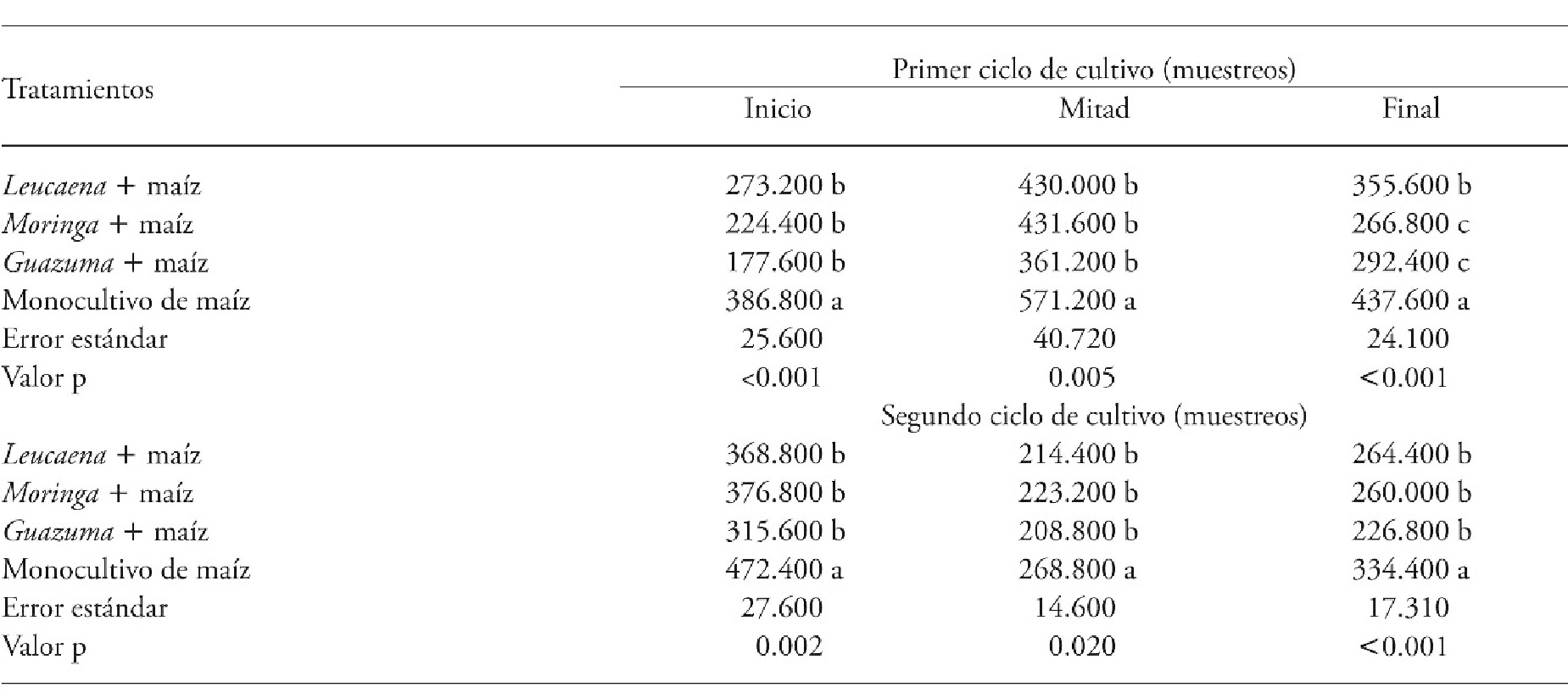
Means in a row with different letters indicate significant differents for each sampling period (p≤0.05).
In both CC the association of forage tree species with maize decreases weed biomass, relative to the monocrop. The leaves of these trees intercepted sunlight, so that weeds did not germinate or did not develop to their reproductive stage. This agrees with Jama et al. (1991), who showed that the association of L. leucocephala with maize can reduce weed biomass up to 90%. The broader leaves of G. ulmifolia may also have contributed to reducing the biomass of these species. It is important to consider that pruning the trees at the beginning of each CC, to obtain forage in a cut and carry scheme, permitted the maize seedlings to receive the light necessary to begin their development. Regrowth of tree leaves gradually shaded the soil surface, affecting weed germination and establishment. We can add the fact of the litter of the trees, among which G. ulmifolia is especially relevant, covered the space like a true carpet with greater effect on weed development.
At the beginning of the experiment, we proposed assessing forage production and maize yield in agroforestry arrays. That is, the trees were not established as a method of weed control. Nevertheless, the effect of woody species on herbaceous plants is important and can contribute to success of the system and reduce the use of external inputs such as herbicides.
Weed diversity
Weed diversity at the beginning and middle of the first and second CC was similar in the evaluated treatments. However, at the end of both cycles, the agroforestry system Guazuma + maize was outstanding because it had fewer weed species than the maize monocrop (Table 3). This agrees with the dynamics of the biomass of these species in the respective treatments: lower values of this variable coincide with fewer species in the treatment.
Table 3 Number of species (in 0.5 m-2) in agroforestry systems consisting of forage trees and maize compared with a maize monocrop in two crop cycles, Xmatkuil, Yucatán, México.
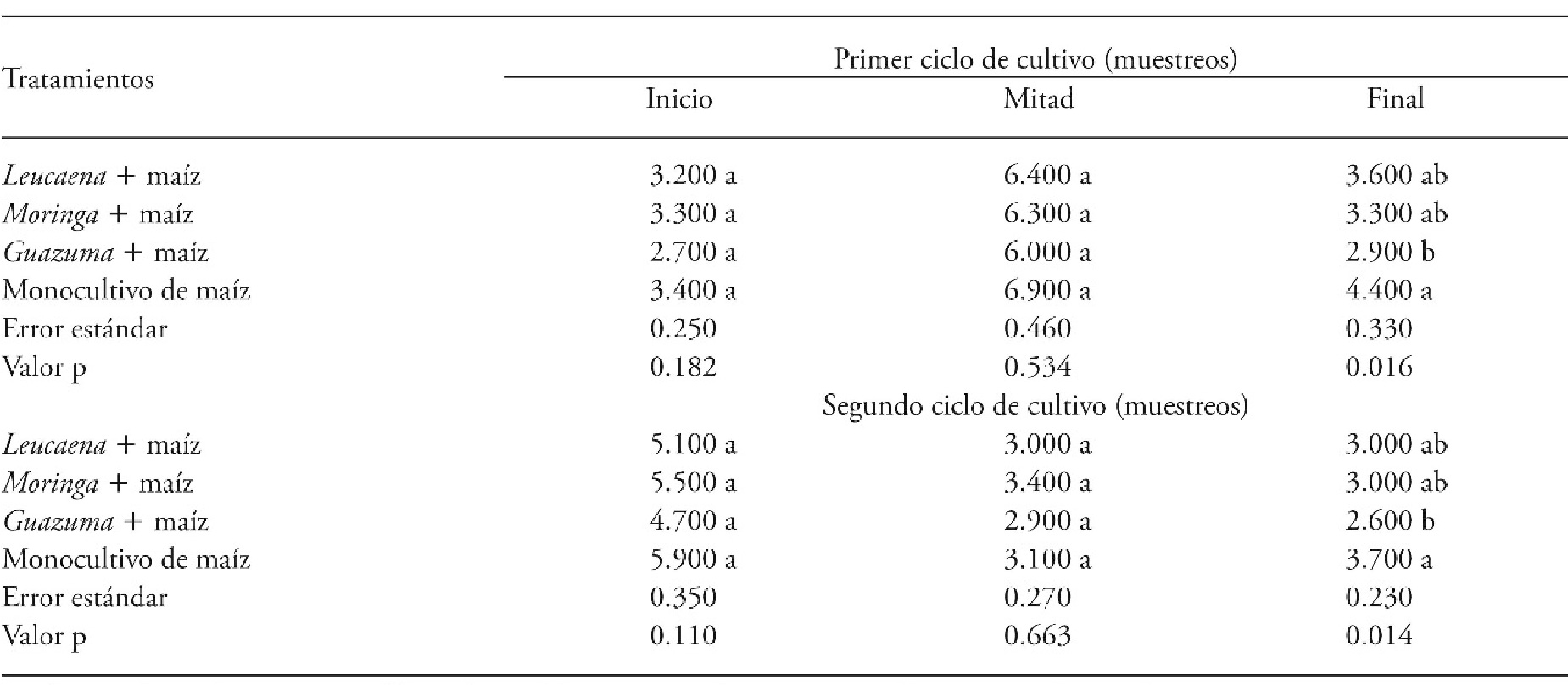
Means in a row with different letters indicate significant differents for each sampling period (p≤0.05).
Forty-seven species belonging to 18 families were identified. Of these, more than 21% were leguminous (Fabaceae), followed by the families Poaceae (12.8%), Asteraceae, (10.6%), Malvaceae, (8.5%), y Euphorbiaceae (8.5%), which together account for more than 61% of all the species found (Figure 3).
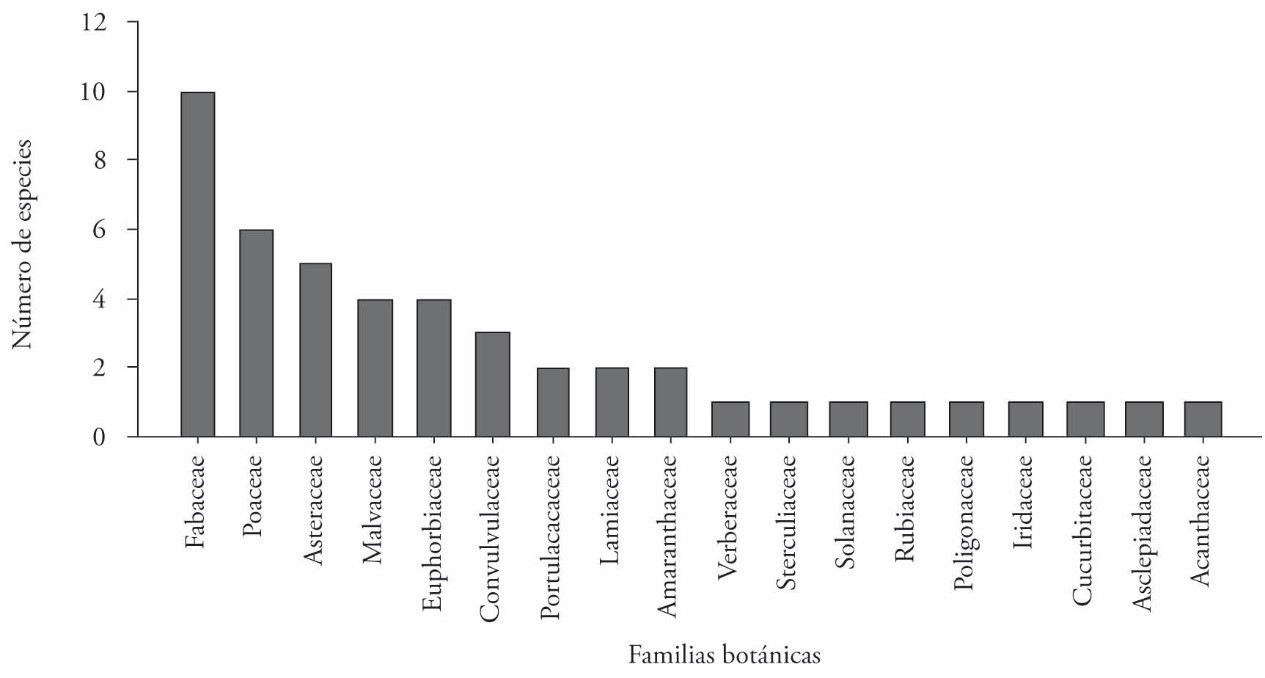
Figure 3 Botanical families of the weeds associated with agroforestry systems consisting of forage trees and maize in Xmatkuil, Yucatán, México.
Diversity of the weed species at the end of the two CC was lower where G. ulmifolia was present than with the presence of other tree species and the maize monocrop. This can be explained by G. ulmifolia’s architecture: its leaves form a dense crown that limits entry of sunlight. Although cover of forage plants was not measured, we could observe that, unlike G. ulmifolia, L. leucocephala, even when branches sprout from the trunk base, permits entry of light, which is necessary for heliophilous plants such as weeds. Moreover, M. oleifera has peculiar growth after pruning: the trunk lengthens and the branches sprout in the upper part, similar to the shape of a feather duster. This, together with the fact that its branches do not form a dense crown, permits the emergence of weed species. In this regard, Segura-Rosel et al. (2012) indicate that G. ulmifolia has large simple leaves that, after falling, persist on the ground as litter for more than four weeks. Besides, Casanova-Lugo et al. (2014) report that the biomass composition of G. ulmifolia has an influence in the number and seasonality of the species under its canopy. Thus, it can create a moist shaded microclimate that would limit the presence of weeds.
Fabaceae, Poaceae and Asteraceae were the predominating botanical weed families in the agroforestry systems formed by forage species and maize. This is in agreement with Caamal-Maldonado et al. (2001), who found that the most important weeds were Asteraceae, Poaceae, Fabaceae and Acanthaceae in their study of the allelopathic effects of four leguminous species, Mucuna deeringiana, Canavalia ensiformis, L. leucocephala and Lysiloma latisiliquum, on weed control in maize crops in Yucatán. But in Havana, Cuba, in a maize crop preceded by a transitory fallow after growing potatoes (Solanum tuberosum), the dominant weed families were Poaceaes and Euphorbiaceaes (Blanco and Leyva, 2010). This suggests that the diversity of weeds is related to edaphic-climatic conditions of the region, as well as to preceding cultural practices related mainly to residual damage by selective herbicides.
Weed density
At the beginning of the first CC, the treatments were similar in terms of weed density. In contrast, at the middle of this cycle, the highest weed density occurred in the maize monocrop. In the same way, at the end of this first cycle, the maize monocrop accumulated a higher weed density; the lowest density corresponded to the agroforestry systems Guazuma + maize and Moringa + maize, with statistically similar values (Table 4).
Table 4 Weed density (individuals m-2) in agroforestry systems consisting of forage trees and maize compared with a maize monocrop in two cropping cycles, Xmatkuil, Yucatán, México.
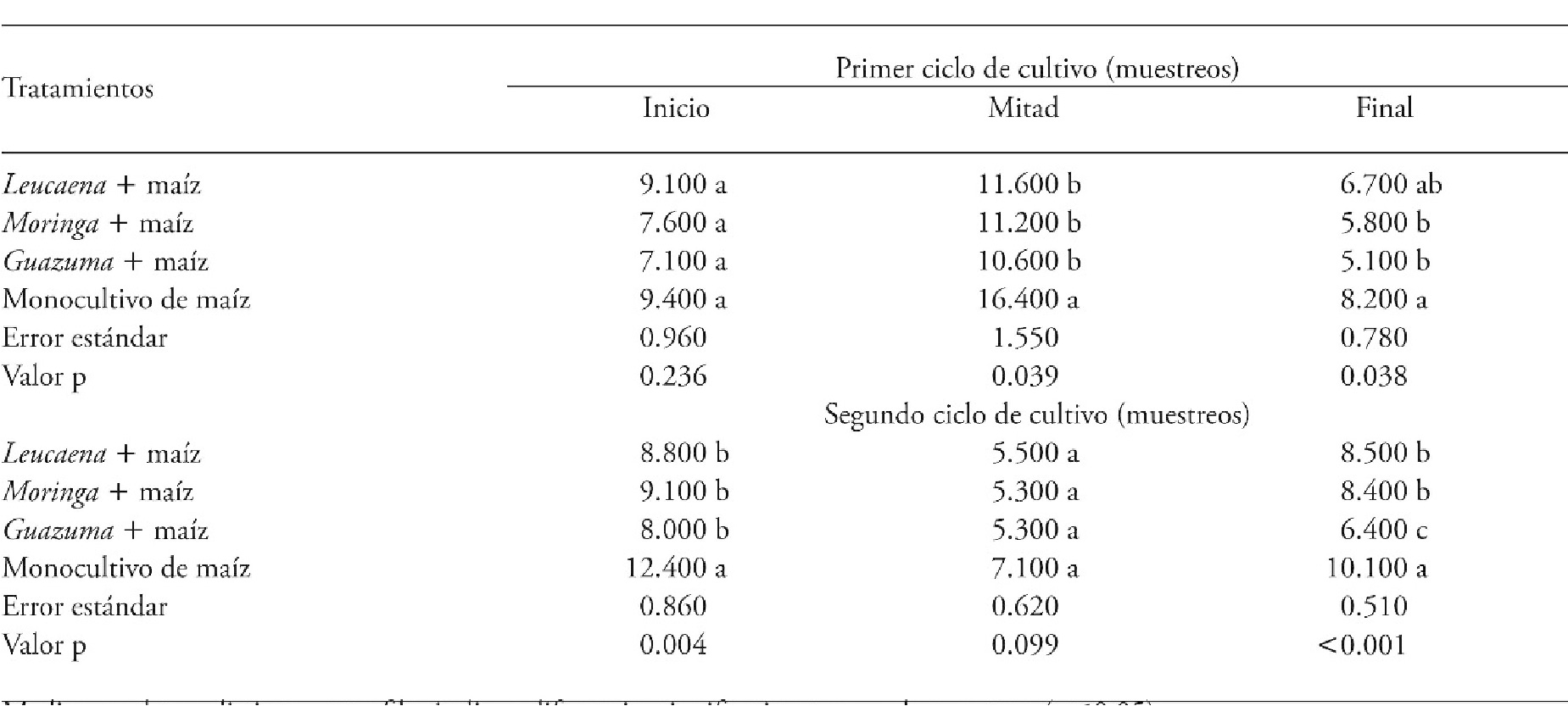
Means in a row with different letters indicate significant differents for each sampling period (p≤0.05).
At the beginning and end of the second CC, the maize monocrop had the highest density of weeds. Nevertheless, at the middle of this second period, there were no significant differences among the assessed systems (Table 4).
Regarding weed density, at the middle and end of the first cycle, the maize monocrop had the highest value, as was expected, given that the presence of the trees in the other systems can reduce shade-intolerant weeds. In the treatments with trees, pruning increases the presence of weed species, but when the trees recover, they again limit the conditions for growth of herbaceous plants. Barbier et al. (2008) indicate that weed density decreases because of the variation in availability of resources such as light and other effects caused by the characteristics of tree species.
In the second CC, the maize monocrop had a larger quantity of weeds, and since there were no tree species in this treatment, weeds exerted more pressure. In this sense, Ayala-Sánchez et al. (2007) mention that in the traditional systems, there is greater presence of weeds because, when there is activity in the soil, the latent seeds germinate and their competitive potential is greater. This was more evident in our study since the site had been cultivated for more than eight years (Caamal-Maldonado et al., 2012). In addition, the trees associated with maize limited weed development. It is also important to mention that in another experiment in the same plot, yield of maize monocrop increased when L. leucocephala and G. ulmifolia foliage was applied as cover (Gallegos-Pérez et al., 2013). This coincides with Jama et al. (1991), who sustain that the use of leguminous trees (i.e. L. leucocephala) as green organic fertilizer in maize cultivation increases yield by 24 to 76%. This was also corroborated by Rippin et al. (1994), who used other leguminous trees, such as Erythrina poeppigiana and Gliricidia sepium. In this context, it is possible to point out that agroforestry systems have the potential to generate sustainable production systems by promoting better weed control.
Potential weed suppression
Potential weed suppression varied from 23% to 36% in the agroforestry systems evaluated, relative to the maize monocrop. The Guazuma + maize agroforestry system had the highest potential, followed by the Moringa + maize and Leucaena + maize systems. Consequently, the maize monocrop had the highest relative proportion of weeds (Figure 4) of the four treatments.
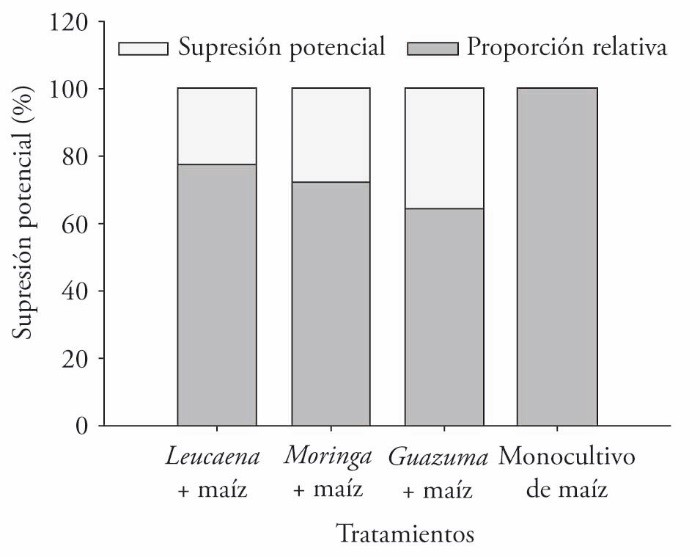
Figure 4 Potential suppression and relative proportion of weeds in agroforestry systems consisting of forage trees and maize compared with a maize monocrop, in two crop cycles in Xmatkuil, Yucatán, México.
The weed-suppressing effect of trees reported by Jama et al. (1991) was also observed in our study, although to a lesser degree; the agroforestry systems we evaluated produced 23% to 36% less weed biomass than the monocrop. This suppressing effect was more evident with maize associated with G. ulmifolia for the reasons given above: leaf size limiting the entry of light, permanence of leaf litter on the soil, and modification of the microclimate. It is also feasible that the nature and content of chemical compounds, especially of secondary metabolites in the plant leaves, have significant effects on weed germination and growth. For example, L. leucocephala leaf litter contains compounds that limit weed development (Caamal-Maldonado et al., 2001). The amino acid mimosine is possibly the main compound responsible for this effect. However, few studies focus on determining whether said compounds act individually or in conjunction, in order to promote them for weed control, particularly in agroforestry systems.
The search for sustainable alternatives for weed management involves maximizing beneficial ecological processes in agroecosystems to maintain weed populations at low thresholds so they do not negatively affect crop growth and development (Caamal-Maldonado et al., 2001; Cummings et al., 2012). To this end, it is necessary to identify the type of biodiversity that is desirable to maintain or broaden in order to propose better practices (Lin et al., 2004; Barbier et al., 2008) and reduce the use of herbicides to minimal levels (Cummings et al., 2012). Although agricultural systems are simpler than natural ecosystems, there are options for redesigning and managing agroecological systems and reduce weed populations (Charudattan, 2001). Among these alternatives are included agroforestry systems, which offer benefits for both the farmer and the environment.
Conclusions
The presence of tree species in agroforestry systems contributes to decreasing weed biomass and cover in associated maize crops, as compared with maize monocrop. In particular, the shape, angle and placement of the leaves seems to be the key element in controlling weeds. The woody species G. ulmifolia, associated with maize, reduces weed biomass, density and diversity, relative to the maize monocrop, contributing to weed control in productive systems. Moreover, they offer an alternative to the use of agrochemicals (i.e. herbicides), which could promote production of safe foods and contribute to the conservation of natural resources
Literatura Citada
Ayala-Sánchez, A., L. Krishnamurthy, J. A. Basulto-Graniel, and J. A. Leos-Rodríguez. 2007. Native tree legumes for improvement of slash-and-burn maize system in Yucatan, Mexico. Terra Latinoam. 25: 195-202. [ Links ]
Bainard, L. D., A. M. Koch, A. M. Gordon, and J. N. Klironomos. 2013. Growth response of crops to soil microbial communities from conventional monocropping and tree-based intercropping systems. Plant Soil 363: 345-356. [ Links ]
Balandier, P., C. Collet, J. H. Miller, P. E. Reynolds, and S. M. Zedaker. 2006. Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighboring vegetation. Forestry 79: 3-27. [ Links ]
Barbier, S., F. Gosselin, and P. Balandier. 2008. Influence of tree species on understory vegetation diversity and mechanisms involved a critical review for temperate and boreal forests. For. Ecol. Manage. 254: 1-15. [ Links ]
Bautista, F., G. Palacio-Aponte, P. Quintana, and J. A. Zinck. 2011. Spatial distribution and development of soils in tropical karst areas from the Peninsula of Yucatan, Mexico. Geomorphology 135: 308-321. [ Links ]
Bellon, M. R., D. Hodson, and J. Hellin. 2011. Assessing the vulnerability of traditional maize seed systems in Mexico to climate change. Proc. Natl. Acad. Sci. 108: 13432-13437. [ Links ]
Blanco, Y., y Á. Leyva. 2010. Abundancia y diversidad de especies de arvenses en el cultivo de maíz (Zea mays, L.) Precedido de un barbecho transitorio después de la papa (Solanum tuberosum L.). Cultivos Trop. 31: 12-16. [ Links ]
Caamal-Maldonado, J. A., J. J. Jiménez-Osornio, A. Torres-Barragán, and A. L. Anaya. 2001. The use allelopathic legume cover and mulch species for weed control in cropping systems. Agron. J. 93: 27-36. [ Links ]
Caamal-Maldonado, J. A., F. Casanova-Lugo, A. González-Moreno, J. Caamal-Caamal, P. Xiu-Canché, A. J. Navarro, J. B. Castillo-Caamal. 2012. Producción de alimento, forraje y servicios ambientales de sistemas agroforestales en Yucatán, México. In: Castelán O., A. Álvarez-Macías, A. Bernués-Jal, J. C. Ku-Vera, y V. Silveira (eds). Avances de Investigación en Producción Animal en Iberoamérica. Ediciones de la UADY, México. pp: 79-104. [ Links ]
Casanova-Lugo, F., J. Petit-Aldana, F. Solorio-Sánchez, D. Parsons, and L. Ramírez-Avilés. 2014. Forage yield and quality of Leucaena leucocephala and Guazuma ulmifolia in mixed and pure fodder banks systems in Yucatan, Mexico. Agrofor. Syst. 88: 29-39. [ Links ]
Casanova-Lugo, F., L. Ramírez-Avilés, F. J. Solorio-Sánchez. 2010. Effect of pruning interval on foliage and root biomass in forage tree species in monoculture and in association. Trop. Subtrop. Agroecosyst. 12: 33-41. [ Links ]
Charudattan, R. 2001. Biological control of weeds by means of plant pathogens: Significance for integrated weed management in modern agro-ecology. BioControl 46: 229-260. [ Links ]
Cummings, J. A., I. M. Parker, and G. S. Gilbert. 2012. Allelopathy: a tool for weed management in forest restoration. Plant Ecol. 213: 1975-1989. [ Links ]
Ferrari, H., C. Ferrari, y F. Ferrari. 2006. CobCal v 2.1. Instituto Nacional de Tecnología Agropecuaria. Paraguay. (Consulta: Marzo 2014). [ Links ]
Gallegos-Pérez, M., A. Caamal-Maldonado, F. Casanova-Lugo, y R. Tzuc-Martínez. 2013. Rendimiento de forraje de especies arbóreas en sistemas agroforestales y su uso como abono verde para el maíz. In: Chay-Canul, A.J., y F. Casanova-Lugo (eds). La contribución del sector pecuario a la seguridad alimentaria en México.. UJAT, Villahermosa, Tabasco, México. pp: 943-946. [ Links ]
García, E. 1988. Modificaciones al Sistema de Clasificación de Köppen. Universidad Nacional Autónoma de México, D.F. [ Links ]
Graefe, S. 2003. Crop and Soil Variability in Traditional and Modern Mayan Maize Cultivation of Yucatán, Mexico. Kassel University Press. [ Links ]
Gutiérrez-Báez, C., y P. Zamora-Crescencio. 2012. Especies leñosas de la selva baja caducifolia de Xmatkuil, Yucatán, México. Foresta Veracruzana 14: 9-14. [ Links ]
Jama, B., A. Getahun, and D. N. Ngugi. 1991. Shading effects of alley cropped Leucaena leucocephala on weed biomass and maize yield at Mtwapa, Coast Province, Kenya. Agrofor. Syst . 13:1-11. [ Links ]
Latournerie, L., J. Tuxill, E. Yupit-Moo, L. Arias-Reyes, J. C. Alejo, and D. I. Jarvis. 2006Traditional Maize Storage Methods of Mayan Farmers in Yucatan, Mexico: Implications for seed selection and crop diversity. Biodivers. Conserv. 15: 1771-1795. [ Links ]
Lin, D., E. Tsuzuki, Y. Dong, H. Terao, and T. D. Xuan. 2004. Potential biological control of weeds in rice fields by allelopathy of dwarf lilyturf plants. Biocontrol 49: 187-196. [ Links ]
Pacini, C., A. Wossink, G. Giesen, C. Vazzana, and R. Huirne. 2003. Evaluation of sustainability of organic, integrated and conventional farming systems: a farm and field-scale analysis. Agric. Ecosyst. Environ. 95: 273-288. [ Links ]
Rippin, M., J. P. Haggar, D. Kass, and U. Kopke. 1994. Alley cropping and mulching with Erythrina poeppigiana (Walp.) O. F. Cook and Gliricidia sepium (Jacq.) Walp.: effects on maize/weed competition. Agrofor. Syst . 25: 119-134. [ Links ]
SAS Institute. 2004. Statistical Analysis System, Users. SAS Institute, Cary, N.C. USA. [ Links ]
Segura-Rosel, A., F. Casanova-Lugo, F. J. Solorio-Sánchez , and A. J. Chay-Canul. 2012. Association of woody species in fodder banks: influence on litter production, decomposition and nitrogen release. Trop. Subtrop. Agroecosyst. 15: 61-69. [ Links ]
Received: January 2016; Accepted: May 2016
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
