











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Las especies leñosas, cultivadas en sistemas silvícolas de monte bajo de turnos cortos (Short Rotation Coppice “SRC”), han adquirido importancia en la oferta de biomasa vegetal (Hoogwijk et al., 2005), la cual puede utilizarse con fines dendroenergéticos. Eucalyptus globulus, E. nitens, E. denticulata (Camps y Marcos, 2002) y Acacia dealbata (Quiroz et al., 2014) son especies con potencial dendroenergético e importancia mundial. El interés por manejar los rodales de estos árboles con métodos de monte bajo ha aumentado, pues en los primeros ocho años producen el doble del volumen que con rodal de regeneración sexual o monte alto (Sixto et al., 2007).
Después de una poda varias especies aumentan considerablemente los rebrotes en los tocones. Esto puede representar competencia entre individuos y poblaciones, y dificultades para el manejo de las plantaciones comerciales, pero aumenta la biomasa por unidad de superficie en menos tiempo. Algunas especies de sauce, álamo, acacia y eucalipto son las más adecuadas para el manejo en monte bajo, por su brotación alta con el vigor suficiente para obtener biomasa con fines dendroenergéticos (Geldres et al., 2004). Los rebrotes de origen proventicio se desarrollan mejor y superan entre 10 y 20 % a los rebrotes adventicios (Vita, 1996).
En eucalipto se recomienda el aclareo a tres rebrotes por tocón después de 18 meses del corte de aprovechamiento, para obtener pulpa y leña; y para producir madera para aserrío se recomienda dejar uno a dos rebrotes por tocón (Geldres et al., 2004; Kenya Forest Service, 2009). La densidad básica de la madera es una característica tecnológica importante por su correlación significativa con la mayoría de las propiedades físicas y mecánicas de la madera (Pereyra y Gelid, 2002; Omonte y Valenzuela, 2011). Además, es uno de los indicadores principales de la calidad y el rendimiento en especies forestales, y se considera un atributo que depende del tipo de árbol, de la edad, del sitio y de los tratamientos silvícolas usados durante el turno (Arango et al., 2001).
El aumento en la producción de biomasa se observa en rodales de especies forestales en densidades altas de población, y se ha detectado que la mortalidad natural (auto-aclareo) regula el número de individuos (Mátyás y Varga, 2000). La información en eucalipto y acacia es limitada para evaluar el efecto de la densidad de plantación en el ajuste natural del número y densidad básica de los rebrotes a lo largo del turno (Miranda y Pereira, 2015; Petro et al., 2015). El estudio y uso adecuado de estas propiedades permite reducir los costos del manejo silvícola y mejora la calidad en la madera de plantaciones de monte bajo. El objetivo de este estudio fue evaluar la dinámica del número y la densidad básica en rebrotes de E. globulus Labill, E. denticulata Cook & Ladiges y A. dealbata Link en diferentes densidades de plantación. La hipótesis fue que todas las especies evaluadas muestran igualdad estadística en la dinámica de brotación y densidad básica en las tres densidades de plantación.
Materiales y Métodos
Descripción del área de estudio
El estudio se realizó en la octava región de Chile, en un experimento establecido en La Aguada (37° 11’ 23” S y 72° 26’ 04” O), comuna de Yumbel. El suelo predominante se conoce como arenal, tiene textura gruesa, es profundo, poco desarrollado, tiene procedencia aluvial y se derivó de arenas volcánicas con origen andesítico y basáltico. El clima predominante es sub-húmedo, con temperaturas entre 28.6 °C en enero y 4.4 °C en julio. La precipitación pluvial media anual es 1093 mm (Novoa y Villaseca, 1989).
Diseño experimental
El experimento se estableció en agosto de 2010, el diseño fue de bloques completos al azar con tres repeticiones. Cada bloque fue un cuadrado de 110 m de lado (12 100 m2) y formado por cuatro cuadrantes, cada uno con nueve parcelas de 18 m por lado (324 m2). Cada parcela tenía una zona de amortiguamiento del efecto de borde y una unidad experimental con 30 árboles útiles. Las especies plantadas fueron E. globulus, E. denticulata y A. dealbata, con tres densidades (5000, 10 000 y 15 000 árboles ha-1). Las tres especies se manejaron con tres frecuencias de corte. Los cuadrantes se cosecharon el primero, segundo y tercer año de edad y se dejó uno sin corte (Figura 1). Las variables se midieron en el cuadrante cosechado en diciembre de 2012 y correspondió a la frecuencia de corte de dos años (tocones de dos años).
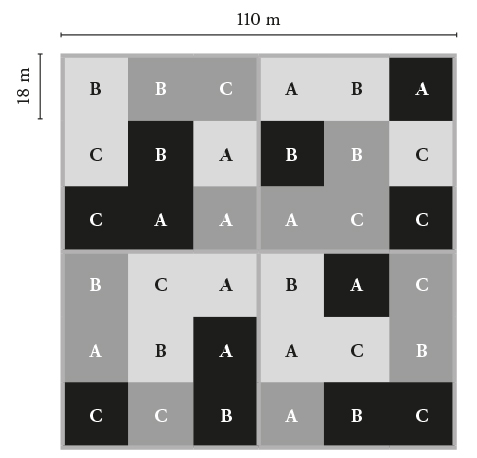
Variables evaluadas
En los tocones obtenidos en diciembre 2012 se evaluó la dinámica del número de rebrotes proventicios por tocón, el número de ellos se contó en la unidad experimental en septiembre de 2014 y 2015 y se midieron la altura y el diámetro a la altura del cuello (DAC) de cada rebrote. Para el análisis combinado de rebrotes se contó en cada tocón el tipo de rebrote (proventicio y adventicio) a los 10, 21 y 31 meses de crecimiento. El tipo de rebrote se definió según sus características fisiológicas y posición; los que brotaron en la corona del tocón y en posición horizontal formados de las yemas de cicatrización se identificaron como adventicios; los mejor unidos al tocón (planta madre) y en posición vertical, originados de yemas latentes dentro de los tejidos vivos del tocón, se identificaron como proventicios (De Simón y Bocio, 1999). En dos periodos de medición (2014 y 2015) se evaluó la densidad básica de cada vástago o rebrote, en rodajas de alturas diferentes a lo largo del fuste. Para esto, tres tocones se seleccionaron en el área de amortiguamiento de cada parcela y en cada una se cortaron todos los rebrotes, se empaquetaron y etiquetaron para identificar el bloque, especie, densidad, tocón y tipo de rebrote (adventicio y proventicio). Los rebrotes se llevaron al laboratorio para su análisis.
La densidad básica de la madera se determinó en todos los rebrotes de cada uno de los tocones. En cada brote se evaluó la altura total con un telescómetro (Messfixs®) de 12 m y el diámetro por sección con un pie de metro (Mitutoyo®) a los 10, 50, 130 cm (diámetro normal=DAP) y luego a cada metro, hasta que se registraron diámetros inferiores a 10 mm. De cada uno de estos puntos a lo largo del fuste se obtuvo una rodaja (de 2.5 cm de espesor). Éstas se ordenaron y etiquetaron en mallas plásticas, se colocaron en recipientes con agua, hasta su completa sumersión, para de esa forma lograr que el volumen de las rodajas sobrepasara el punto de saturación de las fibras.
Cuando las rodajas estuvieron completamente saturadas (peso constante) se determinó su volumen por el método de desplazamiento de agua por muestra (cm3 de agua=g) según la Norma NCh 176/2 (INN, 1986). El peso del agua desplazada se determinó en una balanza electrónica (Snowrex® con precisión 0.01 g), sobre ésta se colocó un recipiente con agua, en el que las muestras estuvieron completamente sumergidas sin tocar los costados ni el fondo; el peso del agua desplazada, que correspondió al volumen de la muestra de madera, se obtuvo al considerar que la densidad del agua es uno (Valencia y Vargas, 1997). Las muestras en bolsas de papel se deshidrataron a 105 °C en un horno (Riossa®) hasta peso constante. El peso del material seco (anhidro) se obtuvo en una balanza electrónica (precisión 0.01 g), inmediatamente después de extraerse del horno para evitar la absorción de humedad ambiental. La densidad básica de la madera correspondió al cociente del peso seco (anhidro) y el volumen saturado:
donde Db: densidad básica (g cm-3) PS: peso seco (g) Vh: volumen saturado (cm3).
Análisis de datos
Los datos se analizaron con estadística descriptiva e inferencial con base en la especie, densidad y fecha de evaluación. Para densidad básica se realizó ANDEVA y comparación de medias con la prueba de Tukey (p≤0.05). El modelo estadístico fue:
donde Yijkl: valor de la variable dependiente, observado, en la especie i, en el año j, en la densidad de plantación k y tipo de rebrote l; µ: media general; SPi: efecto de la i-ésima especie; Aj: efecto del j-ésimo año; Dk: efecto de la k-ésima densidad de plantación; Trl: efecto del l-ésimo tipo de rebrote, SP(A)ij : efecto de la interacción entre la i-ésima especie y j-ésimo año; SP(D)ik: efecto de la interacción entre la i-ésima especie y la k-ésima densidad; SP(Tr)il: efecto de la interacción entre la i-ésima especie y el l-ésimo tipo de rebrote; Eijkl: error aleatorio residual.
Los datos se analizaron con SAS Versión 9.2 (SAS Institute, 2008), Infostat® (Universidad Nacional de Córdoba, Argentina) y Sigma Plot® 8.1 (Systat Software, Inc. San José California, USA).
Resultados y Discusión
Rebrotes proventicios
Acacia dealbata mostró la cantidad mayor de rebrotes proventicios en la densidad de 5000 árboles ha-1, comparado con los otros tratamientos (Figura 2). En 2014 cerca de 85 % de los tocones tuvieron cuatro a cinco rebrotes en las densidades de 10 000 y 15 000 árboles ha-1. El número máximo fue siete rebrotes, y en promedio medían 2.46 de altura y 21 mm de diámetro a la altura del cuello (DAC) en las tres densidades. Con el tiempo la altura promedio de los rebrotes llegó a 3.2 m y DAC de 27 mm; el número de rebrotes en las densidades de 5000 y 10 000 árboles ha-1 se ajustó entre densidades de plantación en 2015, cuando 91 % de los tocones presentaron cinco rebrotes y alcanzaron un máximo de siete que representó el 9 % restante (Figura 2). Con 15 000 árboles ha-1, 87 % de los tocones mostraron solo tres rebrotes proventicios, y hasta cinco en el resto de los tocones (Figura 2). Con el tiempo, en esta densidad el número de rebrotes proventicios disminuyó; esto pudo ser efecto de la competencia por espacio y nutrientes inter e intra tocones. Valero y Picos (2009) obtuvieron resultados similares en parcelas invadidas de A. dealbata en Galicia; ellos registraron brotación promedio entre 7 y 21 rebrotes por tocón de las parcelas evaluadas en las primeras fechas de crecimiento.
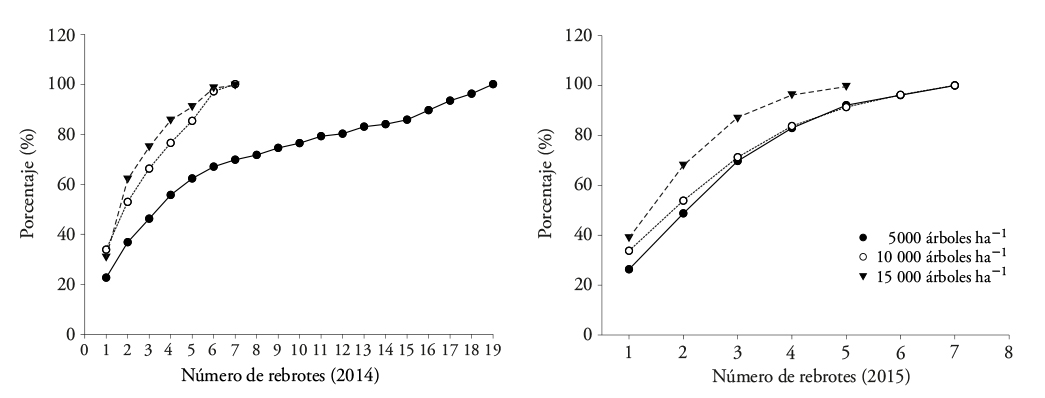
Figura 2 Proporción de tocones en Acacia dealbata que mostraron rebrotes proventicios en dos periodos de evaluación (2014 y 2015).
Eucalyptus denticulata mostró una capacidad considerable de rebrote en todas las densidades de plantación, en número de rebrotes, altura y diámetro. En 2014, cerca de 80 % de los tocones mostraron tres rebrotes proventicios, con máximo de siete y ocho en los otros tocones (Figura 3) con altura promedio de 2.33 m y un DAC de 21.57 mm en las tres densidades. El número de rebrotes proventicios se redujo en 2015; en las densidades de plantación de 10 000 y 15 000 árboles ha-1 94 % de los tocones mostraron dos rebrotes (Figura 3). En el caso de la densidad de plantación de 5000 árboles ha-1, 91 % de los tocones presentaron tres rebrotes y los demás desarrollaron hasta cuatro. La altura promedio de los rebrotes llegó a 4.77 m y el DAC a 42.91 mm.
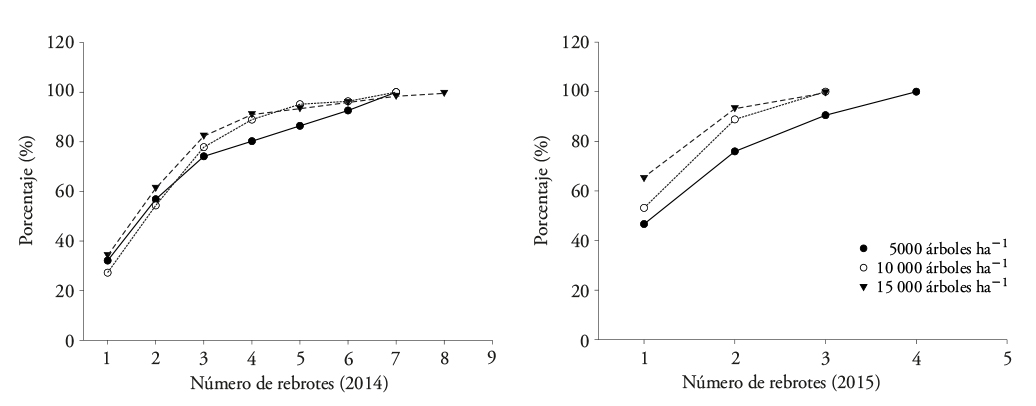
Figura 3 Proporción de tocones de Eucalyptus denticulata que mostraron rebrotes proventicios en dos periodos de evaluación (2014 y 2015).
En el 2014 el total de los tocones de E. globulus, en las tres densidades de plantación, mostró entre cuatro y cinco rebrotes proventicios (Figura 4). Aunque está especie presentó número menor de rebrotes, mostró características mejores en su porte (en ese año tuvo 3.35 m de altura y 29 mm de DAC). En el 2015, cerca del total de los tocones redujo entre dos y tres rebrotes, en altura promedio de 7.2 m y DAC de 55 mm. Además, una proporción alta de tocones presentó de uno a dos en la densidad de 15 000 árboles ha-1 (Figura 4). Esto afecta la producción de rebrotes por competencia de espacio y luz. Villegas (2003)4 evaluó el efecto del espaciamiento del tocón en el número de rebrotes en E. globulus a los 60 d de crecimiento. El efecto no significativo lo atribuyó a la evaluación temprana (2 meses), cuando el crecimiento inicial del rebrote depende de la planta madre (tocón), que aporta soporte y nutrientes. Esto no sucedió después de 24 meses de crecimiento en nuestro estudio, donde los rebrotes compitieron inter e intra tocones, con influencia significativa en la densidad de plantación.
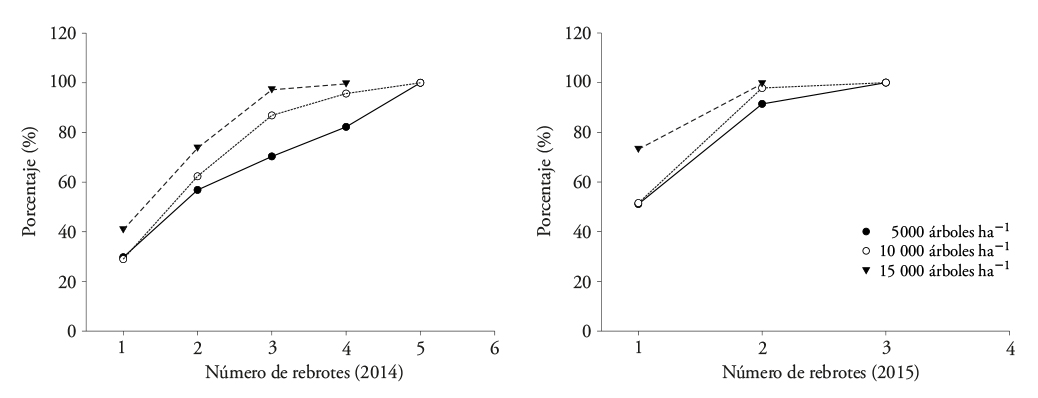
Figura 4 Proporción de tocones de Eucalyptus globulus que mostraron rebrotes proventicios en dos periodos de evaluación (2014 y 2015).
Las tres especies variaron en altura y diámetro como ajuste del número de rebrotes, en función del número de árboles ha-1 y del tiempo. Esta información podrá usarse para adecuar planes de manejo, reducir costos o evitar la replantación y utilizar el ajuste natural del número de los rebrotes en turnos cortos, para multiplicar la cantidad de biomasa por unidad de superficie, con fines dendroenergéticos o mejorar la calidad de la madera mediante podas, para ser aprovechada como aserrío u obtención de pulpa.
Análisis combinado de rebrotes adventicios y proventicios
La evaluación en fechas diferentes permitió visualizar la dinámica del número de rebrotes adventicios y proventicios en cada especie, densidad de plantación y periodo de crecimiento (Cuadro 1). En A. dealbata con 10 meses de crecimiento, en 15 000 árboles ha-1 se observó un número alto de rebrotes adventicios, ocho en promedio y hasta 37 rebrotes excepcionalmente. El número alto de rebrotes adventicios en la densidad de 15 000 árboles ha-1 disminuyó continuamente y a los 21 meses había cinco y tres a los 31 meses. En la densidad de 10 000 árboles ha-1 la respuesta fue irregular y se alcanzó en promedio dos rebrotes adventicios, con máximo de siete por tocón al final de la evaluación. En la densidad de 5000 árboles ha-1 el número de rebrotes adventicios fue menor, con fluctuación promedio de dos y tres en las tres fechas.
Cuadro 1 Número de rebrotes proventicios y adventicios en tocones de especies dendroenergéticas, evaluadas en tres fechas y densidades de plantación.
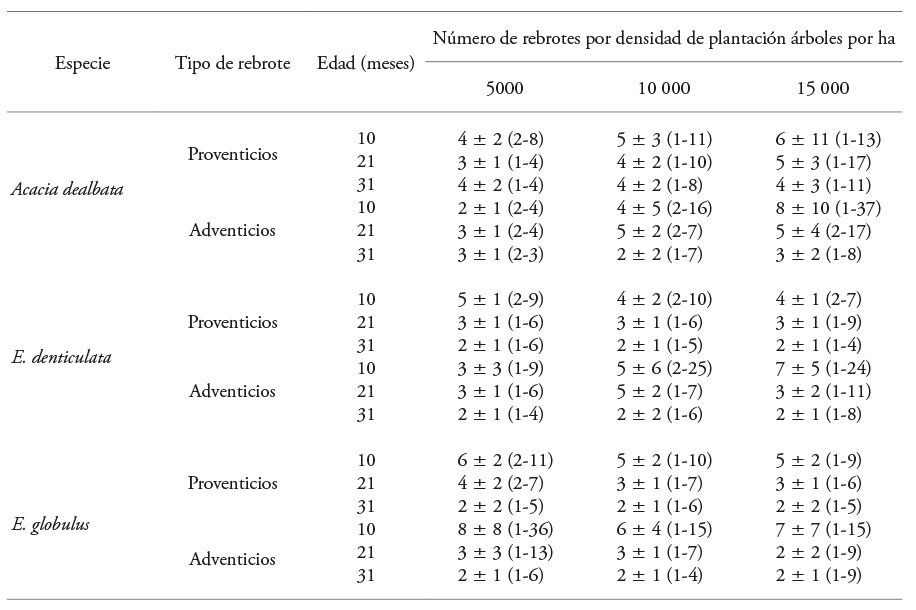
Valor medio, ±=desviación estándar y rango observado (mínimo-máximo).
En esta especie, en la primera evaluación (10 meses), en la densidad de 5000 árboles ha-1 el número de rebrotes de tipo proventicios mostró cuatro rebrotes y un intervalo de dos a ocho rebrotes. Además, en promedio se redujo este tipo de rebrote a los 21 meses, entre uno y cuatro y promedio de tres, y mantuvo cuatro en la evaluación final (31 meses). En las densidades mayores se redujo el número de rebrotes y la variabilidad entre tocones con el tiempo. Después del corte, en la densidad menor el tocón quedó expuesto a la radiación solar directa y al viento, lo que aceleró el proceso de cicatrización de la zona del corte, esto redujo la producción de tejido calloso, en el que se originan las yemas de tipo adventicio. Lo anterior favoreció la formación de rebrotes proventicios, con promedio de cuatro en todas las densidades de plantación, a los 31 meses después del corte.
Eucalyptus denticulata mostró cantidad alta de rebrotes adventicios en la primer fecha de evaluación, con cinco en la densidad de 10 000 árboles ha-1 y siete en la de 15 000 árboles ha-1 (Cuadro 1). Además, hubo variación amplia entre tocones (máximos de hasta 25 rebrotes adventicios por tocón). Esta respuesta disminuyó en las evaluaciones a los 21 y 31 meses, en todas las densidades. Este patrón también se observó en los rebrotes proventicios (de dos a cinco), con la cantidad mayor en 5000 árboles ha-1.
En la primera evaluación, después de 10 meses de crecimiento, en E. globulus se observó el número promedio mayor (ocho) de rebrotes adventicios en la densidad de 5000 árboles ha-1, pero el máximo fue 36 (Cuadro 1). En las evaluaciones siguientes (21 y 31 meses después del corte) los valores disminuyeron a tres y dos, respectivamente. También en la primera fecha el número de rebrotes proventicios en esta especie varió de cinco a seis en las tres densidades y disminuyó en el tiempo a dos rebrotes por tocón en las tres densidades (Cuadro 1). Con el tiempo el número de rebrotes, tanto adventicios como proventicios, se redujo en todas las especies y densidades de plantación principalmente por la eliminación natural de los rebrotes débiles, que disminuyen su crecimiento debido a la competencia por espacio y radiación solar (Sarmiento y Ávila, 2007; Goel y Singh, 2008). La disminución de la intensidad luminosa influye en el inicio del rebrote (Meier et al., 2012).
La importancia del diámetro del tocón se ha demostrado en la probabilidad de rebrote de algunas especies arbóreas, y los árboles con tallos más gruesos tienen menor capacidad de rebrote (Sands y Abrams, 2009). En nuestro estudio los tocones jóvenes de E. denticulata, de 2 años y diámetros entre 5 y 8 cm y 7 cm en E. globulus y A. dealbata, mostraron buen rebrote. Mc Carthy et al. (2014) reportan diferencias en el número de rebrotes entre clones, edad de corte y altura del tocón.
Densidad básica
La densidad básica de la madera mostró diferencias significativas entre especies y periodos de evaluación; pero los tipos de rebrotes (adventicios y proventicios) no fueron diferentes en las tres especies estudiadas. En 2014, E. denticulata mostró los valores mayores y los de A. dealbata fueron los menores (Cuadro 2). Los valores de E. globulus concordaron con los documentados por Igartúa y Monteoliva (2010) de densidad básica de la madera, en plantaciones adultas de tallo único (unifustales), entre 480 y 503 kg m-3.
Cuadro 2 Valores medios de densidad básica de rebrotes en tres especies con potencial dendroenergético, establecidas en tres densidades de plantación.
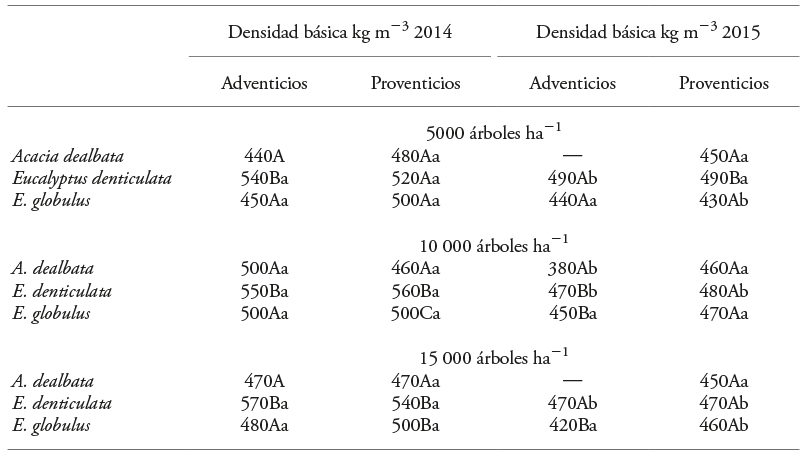
Letras mayúsculas diferentes muestran diferencias significativas entre especies dentro del mismo año; letras minúsculas diferentes muestran diferencias entre años (p≤0.05).
En 2015 los valores de la densidad básica se redujeron en las tres especies, aunque en E. denticulata fue superior. Los valores de A. dealbata (380 y 460 kg m-3) (Cuadro 2) fueron similares a los obtenidos (densidad básica de la madera desde 318 hasta 544 kg m-3) por Pinilla y Hernández (2010) en plantaciones de la misma especie con un tallo, en localidades de la región del Biobío. Ellos también observaron que los valores menores (414 a 466 kg m-3) correspondieron a los árboles jóvenes y variación entre los sitios experimentales; la sección muestreada del tallo presentó densidad básica mayor en la base del fuste (558 a 590 kg m-3).
Variación de la densidad básica de la madera dentro del árbol también se ha observado en eucalipto. Esta se relacionó con las edades formativas del tallo y la morfología de las fibras (Igartúa y Monteoliva, 2010). En E. regnans, la densidad básica disminuyó (404 kg m-3) entre la base y 4.5 % de la altura total del árbol y luego aumentó (462 kg m-3) hasta 70 % de la altura del árbol (Omonte y Valenzuela, 2011). La reducción de la densidad básica registrada entre años en nuestro estudio se debió principalmente al efecto del crecimiento primario, el que modificó la posición de la distancia desde la base en relación con los sitios de muestreo a diferentes alturas del fuste (Igartúa et al., 2002).
Conclusiones
Acacia dealbata, Eucalyptus denticulata y E. globulus presentan brotación en tocones de 2 años y pueden manejarse como cultivos de monte bajo (SRC). Los tallos por unidad de superficie aumentarán, a pesar de la disminución considerable de rebrotes (proventicios y adventicios). Aunque la densidad básica de la madera de E. denticulata es menor, las tres especies muestran características similares a las de plantaciones adultas de tallo simple, por lo que pueden usarse para la producción de biomasa dendroenergética desde rebrotes, lo que evita los costos de replantación y pueden usarse en varios turnos de aprovechamiento.

