











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Pinus pinceana Gordon es un pino piñonero endémico de México, su distribución está restringida a 17 poblaciones pequeñas y aisladas, en montañas semiáridas de la sierra Madre Oriental, en suelos de origen calcáreos, entre 1400 y 2300 msnm, en los estados de Coahuila, Zacatecas, San Luis Potosí, Nuevo León, Querétaro e Hidalgo (Ledig et al., 2001; Favela et al., 2009; Villarreal-Quintanilla et al., 2009). El sobrepastoreo con caprinos y la recolección de leña y conos han colocado a la especie en riesgo de extinción (Richardson y Rundel, 1998; Ledig et al., 20011). Este árbol piñonero está sujeto a protección especial por el gobierno mexicano, por lo que se requieren estrategias de conservación y manejo (Ledig et al., 2001; Molina-Freaner et al., 2001).
El límite sur de la distribución natural de P. pinceana se ubica en el estado de Hidalgo, con dos poblaciones, El Arenalito y San Cristóbal, en el Municipio El Cardonal (Ramírez-Herrera et al., 2010). Las poblaciones que habitan los márgenes de la distribución natural de una especie están fragmentadas y aisladas de otras, con baja capacidad reproductiva debido a la producción de semilla poco viable (Westemeier et al., 1998; Mápula-Larreta et al., 2007). En estas zonas las especies están limitadas por el ambiente que ejerce presión de selección fuerte (Mosseler et al., 2000). Según Ledig et al. (2002), el factor primario de la declinación gradual de muchas coníferas hasta los niveles actuales es el calentamiento global, desde el periodo de las glaciaciones. Así, este pino pudo haberse desplazado hacia el norte sobre la parte interior de la sierra Madre Oriental, quedando algunas poblaciones pequeñas en Hidalgo aisladas de la mayoría de las poblaciones con mayor extensión de terreno y ubicadas en Coahuila, Nuevo León y Zacatecas (Figura 1). Además, si los pronósticos de cambio climático se materializan a corto plazo, las especies tenderán a moverse hacia el norte, con la desaparición de las poblaciones sureñas (Sáenz-Romero et al., 2010), ya que la migración natural estará limitada por los mecanismos naturales de dispersión de semillas.
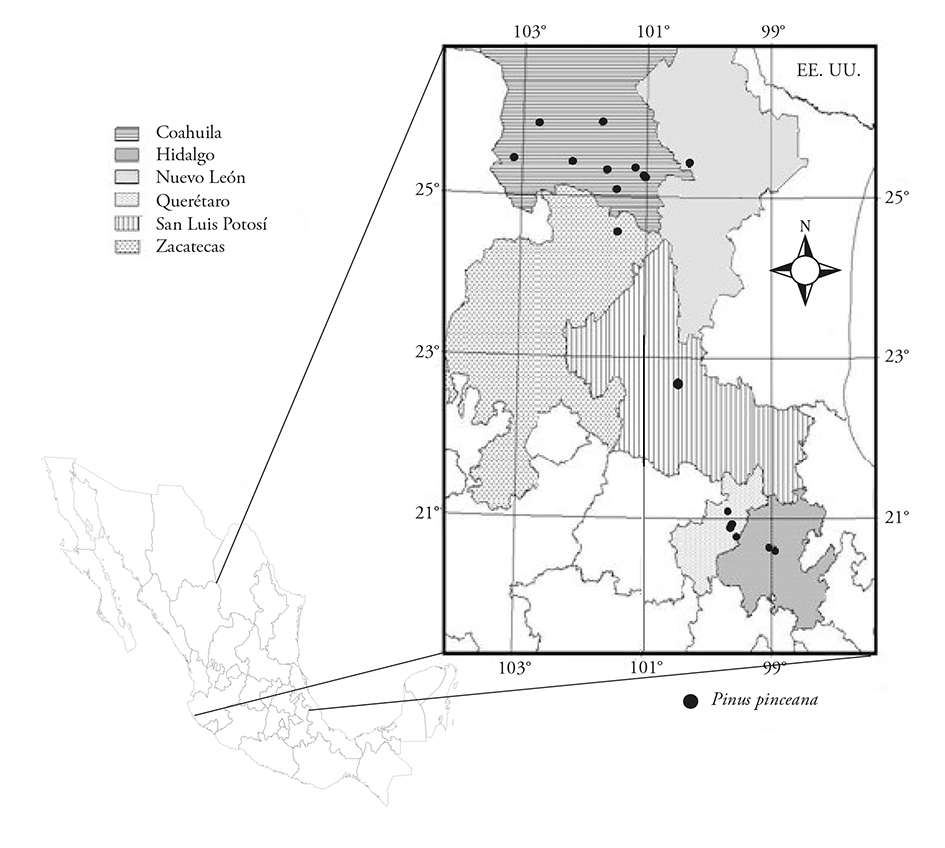
Figura 1 Distribución de poblaciones naturales de Pinus pinceana Gordon en México (basado en Favela et al., 2009; Ledig et al., 2001; Molina-Freaner et al., 2001, y datos propios).
La producción de semilla es irregular y en periodos variables de años (Cain y Shelton, 2000) y para monitorear la capacidad reproductiva de una población se usa el análisis de producción de semilla, obtenido por el potencial de producción de semilla, la cantidad de semilla llena y vana, y la relación entre el peso de semilla llena con el número total de semilla por cono (Bramlett et al., 1977; Mosseler et al., 2000). Los renuevos son escasos en las poblaciones de P. pinceana al sur de su distribución natural, por lo cual el objetivo de este estudio fue determinar la producción de conos por árbol y la proporción de semilla llena y vana en la cosecha de tres ciclos de producción consecutivos en dos localidades del sur de P. pinceana en Hidalgo para determinar posibles problemas en la conservación de este piñonero en estas localidades.
Materiales y Métodos
Las dos poblaciones de estudio están ubicadas en el municipio El Cardonal, Hidalgo, separadas por 8 km (Cuadro 1). La población de San Cristóbal se encuentra dentro de la Barranca de Tolantongo, con vegetación arbórea de bosque bajo de Juniperus flaccida Schltdl. (Hiriart y González, 1983). La población El Arenalito está en la cañada del mismo nombre, con vegetación en el estrato arbóreo dominado por Pinus cembroides Zucc. y Yucca filifera Chabaud (Villarreal-Quintanilla et al., 2009).
Cuadro 1 Caracterización de las poblaciones de Pinue pinceana Gordon localizadas al sur de su distribución natural.
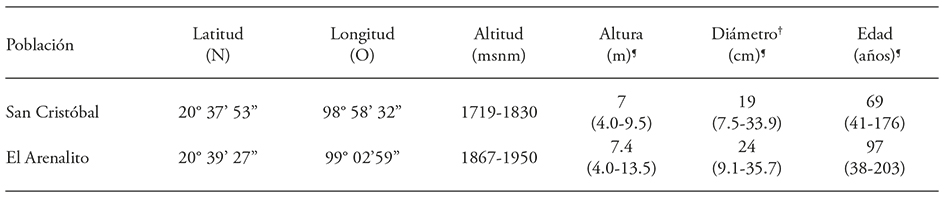
+Diámetro normal. ¶ Valores promedio y extremos (entre paréntesis) de los arboles muestreados.
En cada población se seleccionaron 25 árboles sanos con conos, separados entre sí por un mínimo de 50 m para cubrir una mayor superficie de cada localidad, los cuales se identificaron para recolectas posteriores. La altura total del árbol se midió con pistola Haga®, el diámetro normal con cinta diamétrica y para la edad se utilizó el taladro Pressler. En septiembre de 2001, 2002 y 2003 se contabilizó en el mismo árbol la producción total de conos y se seleccionaron al azar cinco conos por árbol para determinar capacidad reproductiva con la metodología de análisis de cono y producción de semilla (Bramlett et al., 1977; Mosseler et al., 2000). Los conos se midieron en largo y después se colocaron en invernadero (45 °C temperatura máxima) para la apertura de las escamas y extraer la semilla. Cada cono se disectó para contar las semillas llenas, vanas y óvulos abortados y las escamas fértiles, que se ubican en la porción central del cono con capacidad para producir óvulos fértiles. Los conos se colocaron en una estufa de secado a 70 °C por 48 h para obtener el peso seco. Por cada cono se pesó el total de semilla llena y se eligieron 20 semillas por árbol para medir el largo y ancho. Estas dos características y la longitud del cono sólo se obtuvieron en el primer año.
El método de análisis de conos proporciona información para identificar la producción de semillas y sus fallas (Bramlett et al., 1977). Los cálculos se realizaron por cono:
Potencial de semilla = no. de escamas fértiles x 2 (dos óvulos por escama).
Semillas llenas y vanas = aquellas que tuvieron megagametofito y embrión son semillas llenas, y como vanas aquellas que no lo tuvieron. La semilla llena se separó de la vana por el método de flotación en una mezcla de agua más etanol en proporción 1:1 en volumen.
Semillas desarrolladas (aquellas que completaron su desarrollo estructural = llenas + vanas + plagadas (no hubo plagadas en ningún cono de los árboles muestreados).
Porcentaje de óvulos abortados = (no. de óvulos abortados / potencial de semilla) x 100.
Porcentaje de semilla llena = (no. de semillas llenas / potencial de semilla) x 100. Esto se conoce como eficiencia de producción de semillas llenas (eficiencia de semillas).
Porcentaje de semilla vana = (no. de semillas vanas / potencial de semilla) x 100.
Peso de 100 semillas = (peso total de las semillas llenas / no. de semillas llena) x 100.
Semillas llenas por kilogramo de semillas llenas = 1,000 x (peso total de las semillas llenas en g / no. de semillas llena).
Eficiencia reproductiva = peso de las semillas llenas por cono / peso seco del cono, como una medida de eficiencia reproductiva que refleja la proporción de la energía utilizada en el esfuerzo reproductivo almacenado en la semilla (Mosseler et al., 2000).
Índice de endogamia = no. de semillas vanas / no. de semillas desarrolladas. Cuando ocurre autofecundación o cruzamientos muy emparentados se expresan genes deletéreos homocigóticos que impiden el desarrollo del embrión, lo cual genera semilla vana (Mosseler et al., 2000).
Para detectar las diferencias entre sitios y fechas de recolecta se usó el análisis de varianza de medidas repetidas (Gumpertz y Brownie, 1993), considerando esos dos efectos como fijos, excepto el error que fue aleatorio, con el modelo:
Yijk= μ+Si+Fj+SFij+εijk
donde Yijk: valor observado de la variable en la k-ésima muestra en la j-ésima fecha en el i-ésimo sitio; μ: media general; Si: efecto del i-ésimo sitio; Fj: efecto de la j-ésima fecha; SFij: efecto de la interacción del i-ésimo sitio con la j-ésima fecha de muestreo; εijk: error asociado a la j-ésima fecha en la k-ésima muestra aleatoria (árbol) del i-ésimo sitio (error de muestreo).
Los análisis se realizaron con PROC MIXED y con la opción REPEATED (Littel et al., 1998), para lo cual se usaron datos promedios por árbol. Entre las variables morfológicas y las de los árboles se realizaron análisis de correlación. Los análisis estadísticos se efectuaron con SAS (SAS Institute, 1998). Los datos de proporciones se transformaron con arcoseno (proporción)0.5 antes de los análisis para mejorar la normalidad (Sokal y Rohlf, 2012).
Resultados y Discusión
No hubo diferencias significativas entre las dos localidades, pero si entre años y la interacción de las poblaciones x años (Cuadro 2). La producción de conos en los 50 árboles (25 de cada población) fue variable durante los tres años de estudio (Figura 2). La producción de conos fue significativamente mayor en 2001, con 2,188 comparado con 747 y 695 conos en 2002 y 2003 al contabilizar los 50 árboles recolectados.
Cuadro 2 Valores de probabilidad obtenidos del análisis de varianza con medidas repetidas de diferentes características de producción de semilla en dos poblaciones de Pinus pinceana Gordon en el estado del Hidalgo, muestreados en tres años consecutivos (2001,2002 y 2003)
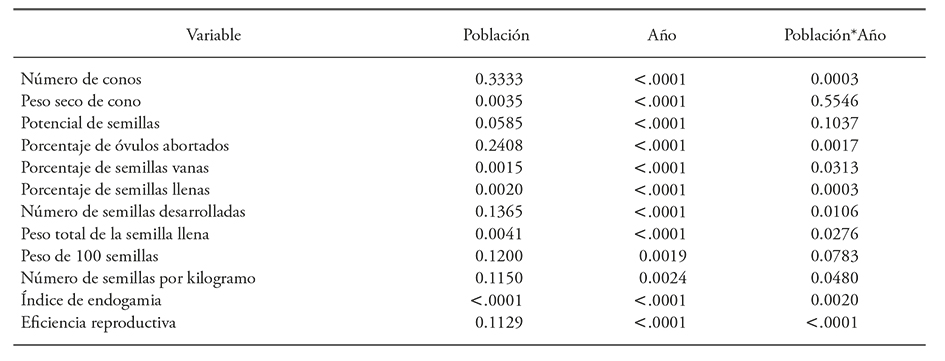
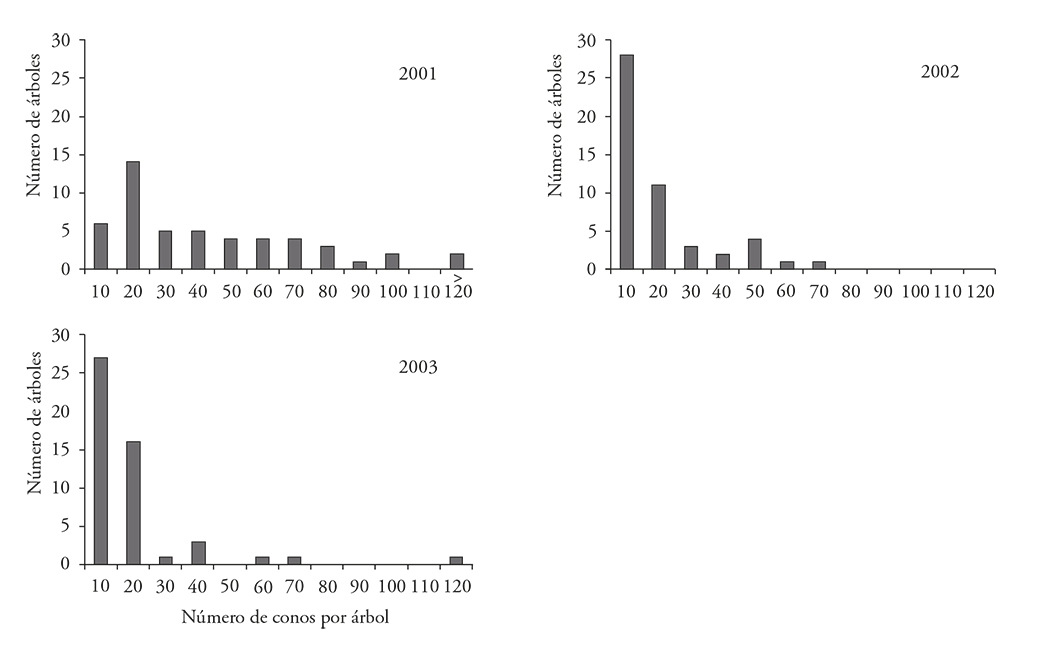
Figura 2 Número de árboles por categoría de producción de conos en dos localidades de Pinus pinceana Gordon del estado de Hidalgo durante tres años de muestreo (N=50, 25 de cada población; la categoría 120 comprende un árbol con 121 y otro con 297 conos en el año 2001, y un árbol con 117 conos en el 2003).
La producción de piñón es errática de un año a otro en un mismo árbol (Flores y Díaz, 1989; Richardson y Rundel, 1998), según la disponibilidad de recursos y condiciones ambientales propias para la reproducción (Romero et al., 1996). Hubo diferencias significativas entre años en San Cristóbal y en El Arenalito, con una reducción en el número de conos en el segundo y tercer año (Cuadro 3), con muy amplias diferencias entre los árboles muestreados (Figura 2). Las diferencias fueron más notorias en la segunda población. Durante los años de producción abundante, las semillas tienen una capacidad germinativa superior y conservarán su viabilidad durante más tiempo; además, el impacto proporcional por el ataque de insectos es menor que en otros años (FAO, 1991; Barner y Olsen, 1994; Kelly, 1994).
Cuadro 3 Valores promedios de diferentes características de producción de semilla en dos poblaciones de Pinus pinceana Gordon en el estado de Hidalgo, muestreados en tres años consecutivos (2001, 2002 y 2003).
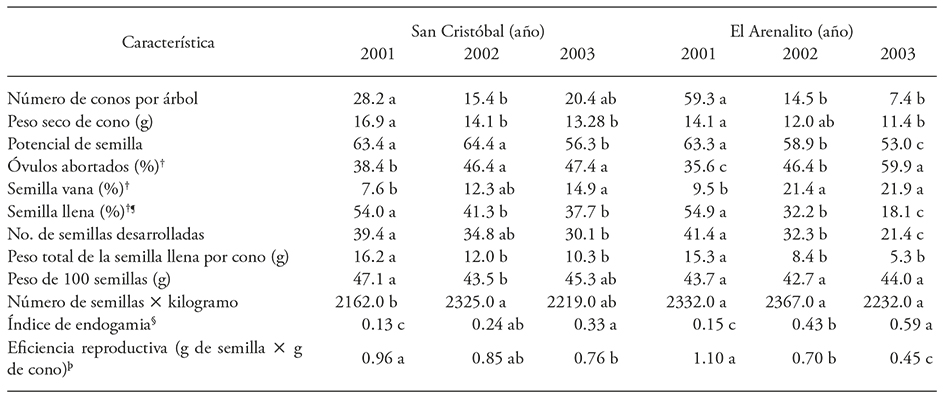
Valores promedio con letras diferentes son estadísticamente diferentes (p≤0.05) entre años dentro de cada población por línea. +Calculado sobre el número de semillas potenciales. ¶Eficiencia de semillas=número de llenas / potencial de semillas. § Relación de semillas vanas sobre las desarrolladas (vanas+llenas). ( Peso de semilla sobre el peso seco del cono.
En San Cristóbal se obtuvo en promedio 34 semillas llenas en 2001 (54% x 63.4 semillas potenciales, Cuadro 3), bajando a 26 y 21 semillas llenas por cono en 2002 y 2003. En el Arenalito se encontraron 35, 19 y 10 semillas llenas por conos en 2001, 2002 y 2003, respectivamente. Lo anterior indica diferencias drásticas en el Arenalito entre años, similares a lo ocurrido en la producción de conos por árbol. La relación semillas vanas sobre desarrolladas (índice de endogamia) varió de 0.13 y 0.15 en el primer año para las dos localidades, lo que es 5 a 6 semillas vanas por cono (7.6% x 63.4 del potencial en San Cristóbal y 9.5% x 63.3 del potencial en El Arenalito, Cuadro 3). Como no se detectó una sola semilla plagada, y la densidad de población es baja, se supone que ocurre cierto nivel de autopolinización o cruzamientos emparentados. En el 2002 y 2003 la proporción de vanas sobre semillas desarrolladas aumentó de 0.24 y 0.43 a 0.33 a 0.59, en San Cristóbal y El Arenalito, respectivamente, lo que representa 8 y 12.7 semillas vanas en el 2002 y 8.5 y 11.4 semillas vanas por cono en 2003 en San Cristóbal y en El Arenalito, respectivamente. La mayor proporción de semillas vanas en El Arenalito en 2003 se debe al menor potencial de semilla que en 2002.
La cantidad de óvulos abortados fue mayor en 2002 y 2003 (de 27 a 32 óvulos abortados por cono). Esto sugiere una falta de polen en la primavera de 2001 y mayor en la de 2002, en particular El Arenalito, lo cual resultó en muchos óvulos no fecundados en la tercer cosecha.
Al generar una curva acumulada de producción de conos se encontró que pocos árboles producen la mayor proporción de semilla y entre más pronunciado sea el arco, menos árboles producen la mayor cantidad de conos. Lo ideal es que cada árbol produjera la misma cantidad de conos y semillas (línea de producción equitativa, a 45°). Diez árboles o menos produjeron 50% de los conos en cada uno de los tres años (Figura 3). Dado que pocos individuos producen la mayor cantidad de germoplasma en un ciclo de producción, se esperaría una menor variabilidad genética en la progenie respecto a la posible, lo cual debe ser más drástico durante el segundo y tercer año, cuando menos árboles producen 50% del total de conos. Además hubo una correlación significativa entre la producción de conos por árbol entre 2002 y 2003 (r=0.53, p=0.003), pero no con la de 2001. Lo anterior indica que no todos los individuos producen la misma cantidad de conos en cada año y que un pequeño porcentaje de árboles contribuye más al establecimiento de plántulas en un año determinado, pero diferentes árboles pueden aportar semilla en tiempo (por eso la baja repetitividad entre años).
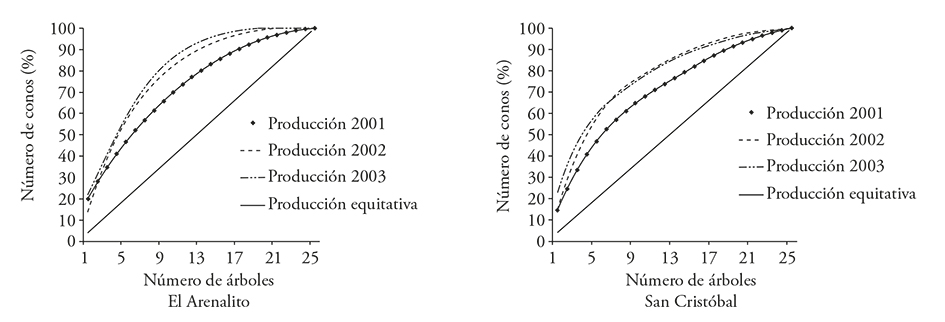
Figura 3 Contribución de árboles individuales a la producción total de conos (%) en cada una de las poblaciones de Pinus pinceana Gordon durante tres años de muestreo.
Aunque hubo diferencias significativas en algunas características, muy poca variación se atribuyó a diferencias entre las dos poblaciones naturales (Cuadro 3). No hubo diferencias significativas entre poblaciones para el número de conos, potencial de semilla, óvulos abortados, número de semillas por kilogramo y en eficiencia reproductiva. La interacción poblaciones x año de recolecta fue significativa en todas las características. Así, la producción de las semillas llenas fue notablemente superior en 2001 y fue escasa en 2002 y 2003, aunque entre estos dos años fue variable según la población. El peso de la semilla fue mayor en 2001 en San Cristóbal, sin diferencias entre años en El Arenalito.
La mayor contribución a la varianza total en 10 de las 11 características analizadas fue la del error, esto es, a la varianza entre árboles dentro de poblaciones y años. En el largo del cono la mayor contribución a la varianza total fue por el árbol; el factor árboles tiene mayor contribución a la varianza en longitud (54%) y ancho de semilla (37%). La variación entre árboles de Pinus es alta y con frecuencia las características de conos y semillas muestran una mayor variación entre árboles dentro de poblaciones que entre ellas (Mosseler, 1992; López-Upton y Donahue, 1995; Flores-López et al. 2005).
Durante el primer año las dos poblaciones presentaron mayores valores de eficiencia de semilla (54 y 55%) y eficiencia reproductiva (0.96 y 1.11 g g-1), lo que representa una mayor proporción de biomasa asignada a la semilla. En las siguientes cosechas los valores fueron reducidos entre 41% a 18% en eficiencia (Cuadro 3, proporción de semillas llenas). Según Flores-López et al. (2012), las coníferas de zonas áridas tienen una menor producción de semillas que aquellas en mejores condiciones ambientales. En Pinus orizabensis D.K. Bailey de Altzayanca, Tlaxcala, la eficiencia fue 29% en un año de evaluación (Sánchez-Tamayo et al., 2005) y 42 a 81% en 12 poblaciones de P. greggii Engelm. ex Parl. (López-Upton y Donahue, 1995). En huertos semilleros de P. banksiana Lamb., P. taeda L., P. elliottii Engelm., P. echinata Mill. y P. palustris Mill. la eficiencia fue hasta 60% usando control de plagas (Bramlett, 1987; De Groot y Schnekenburger, 1996). Por el contrario, es común encontrar problemas de producción de semillas en poblaciones pequeñas: 37% en P. arizonica Engelm. (Narváez, 2000) y 1.97% en P. leiophylla Schiede ex Schltdl. et Cham. (Morales et al., 2010), valor que mejora a 17% con el establecimiento de un huerto semillero sexual de esta especie (Gómez et al., 2010). Por lo tanto, se puede concluir que estas dos localidades de P. pinceana estudiadas producen aceptables cantidades de semilla cuando producen muchos conos, pero será relativamente escasa cuando hay pocos conos los que tendrán pocas semillas llenas.
En otras coníferas de poblaciones fragmentadas hay un comportamiento menos adecuado; en Picea mexicana Martínez el valor de eficiencia fue 7% y el índice de endogamia fue 0.73 a 0.84 (Flores-López et al. 2005; Flores-López et al. 2012), y en Pseudotsuga menziesii (Mirb.) Franco la eficiencia de producción de semilla fue 25.5% en nueve localidades mexicanas, lo que se relaciona al número reducido de árboles de esos rodales (Mápula-Larreta et al., 2007). Según Owens et al. (2005), la autofecundación y la escasez e inviabilidad del polen son las causas principales del aborto de semillas en coníferas. En las dos poblaciones de P. pinceana los valores de este índice variaron entre 0.13 y 0.15 en 2001 de abundante producción de conos, pero aumentó hasta 0.33 en San Cristóbal y 0.59 en El Arenalito, ambos en 2003.
El porcentaje de semillas llenas está determinado por la abundancia y calidad de polen que es variable de un año a otro (Sorensen, 1973; Todhunter y Polk, 1981). El promedio de óvulos abortados en el tercer año es superior a los primeros años y fue mayor en El Arenalito. También se obtuvo el porcentaje más alto de semillas vanas. La producción de éstas se ha relacionado con la autopolinización, el daño por insectos y hongos (Bramlett et al., 1977). Al utilizar conos sin daños externos, no se detectaron semillas dañadas por insectos, por lo que la autopolinización debe ser la principal causa de las semillas vanas aquí encontradas.
La reducción en la producción de semilla llena por cono en la segunda y tercera cosecha es consistente con el efecto esperado de depresión endogámica en años no semilleros y en poblaciones pequeñas y aisladas, donde la cantidad y calidad de polen es altamente variable (Mosseler et al., 2000). En un estudio de cinco poblaciones de P. pinceana en los estados de Querétaro, San Luis Potosí y Coahuila, se encontró un tamaño de población de 1000 a 3000 árboles totales (Molina-Freaner et al., 2001). Los rodales de Hidalgo tienen menor tamaño que los ya mencionados. En nuestro estudio la densidad fue 117 árboles ha-1 en El Arenalito y 107 árboles ha-1 en San Cristóbal. En poblaciones pequeñas y de baja densidad se puede esperar un incremento en endogamia y homocigosis, la expresión de alelos deletéreos y una reducción en la capacidad reproductiva y viabilidad (Mosseler, 2000).
En el análisis de correlación se esperaba que algunas de las relaciones como el largo y peso del cono, y el número de semilla llenas con el peso total de las semillas por cono, fueran significativas por la interdependencia de caracteres, pero no lo fue así. Otras, como la eficiencia reproductiva y la eficiencia de semilla no presentaron una correlación significativa; sólo algunas variables se correlacionaron (Cuadro 4).
Cuadro 4 Coeficientes de correlación de Pearson significativos (p≤0.05) entre características de producción de conos y semillas de Pinus pinceana Gordon (n=50)
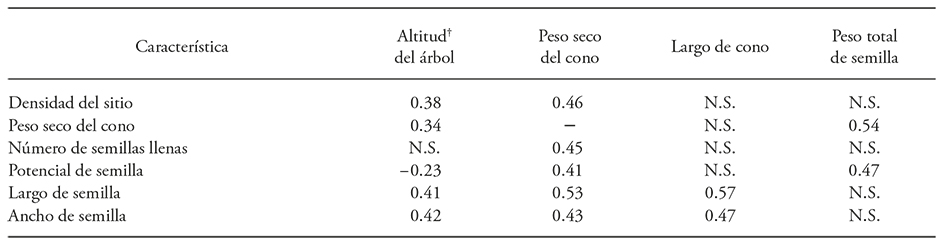
N.S. No significativa. +Altitud del árbol dentro de cada población en msnm.
El largo del cono tuvo correlación positiva con el tamaño (largo y ancho), pero no con la cantidad de semilla llena y el peso seco del cono con el peso total de la semilla; si se desea semilla grande deberán recolectarse conos con el tamaño mayor. Para el establecimiento de la regeneración es importante porque las semillas de tamaño mayor producen plántulas más vigorosas durante el primer año de vida (Castro, 1999; Mueller et al., 2005). El potencial de semilla tiene una correlación positiva con el peso de cono (r=0.41); esto indica que una mayor cantidad de energía destinada a las estructuras reproductivas es favorable para la producción de óvulos fértiles.
La población El Arenalito es muy sensible al estrés hídrico, ya que en una prueba de sequía y alta temperatura plantas de dos años de edad procedentes de esta localidad, fueron más sensibles que las de San Cristóbal y de otras 10 localidades de P. pinceana (Martiñón-Martínez et al., 2010). Los dos rodales de P. pinceana estudiados en Hidalgo son susceptibles al cambio climático dada la ubicación en la periferia austral, ya que de continuar el aumento de la temperatura y de mayor déficit hídrico (Sáenz-Romero et al., 2010). El escenario más probable es la extirpación de poblaciones de los extremos altitudinales inferiores y extremos sur en el Hemisferio Norte (Aitken y Withlock, 2013), como se ha pronosticado para las localidades extremas de Pseudotsuga menziesii de Oaxaca (Rehfeldt et al., 2014), o las sureñas de Pinus leiophylla (Sáenz-Romero et al., 2015) y la reducida plasticidad fenotípicas de las procedencias del sur de P. pinceana (Martiñón-Martínez et al., 2010). Por lo tanto, deben realizarse acciones de conservación in situ y ex situ (Ledig et al., 2001), aprovechando al máximo la recolecta en los años semilleros, cuando la calidad de la semilla es mejor y la obtención de la semilla es más barata. Además, es necesario efectuar actividades de restauración como la plantación de árboles procedentes de ambas poblaciones de manera recíproca, para fomentar el flujo genético entre éstas (Aitken y Withlock, 2013), e incluso considerar las poblaciones del estado de Querétaro, por sus similitudes ecológicas y morfológicas, por ejemplo, similares crecimientos y cantidades de cera en las acículas (Ramírez-Herrera et al., 2010) lo cual ayudaría a incrementar la variabilidad genética al fomentar el flujo de genes.
Conclusiones
El primer año fue de alta producción de conos y semillas, con valores de eficiencia de producción de semilla y la cantidad de semilla vana adecuados y mejores a los registrados en otras coníferas. En el segundo y tercer año disminuyó la producción de conos, se redujo el valor de las características reproductivas, entre ellos menos semillas llenas y más vanas. Existe un desbalance en la producción de conos entre los árboles; pocos de estos producen la mayor parte de la cosecha, y entre años fueron diferentes los más productores. La disparidad entre cosechas coincide con la periodicidad en la producción de semillas documentada en los piñoneros. No hay indicaciones de que las poblaciones sufran los efectos de depresión por endogamia, la que se relaciona más a la variación natural entre años de alta o escasa producción, y la disparidad entre árboles muy productores de conos y los que producen poco.

