











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En México se reconocen más de 1200 especies y subespecies de orquídeas y se ubican al sur del Trópico de Cáncer, desde las costas del Pacífico a las del Golfo, en regiones de hasta 3500 msnm (Espejo y López-Ferrari, 1998). Oncidium y Laelia son dos géneros de orquídeas distribuidas en México y tienen valor comercial alto. La belleza de sus flores las hace muy atractivas, por lo cual en las poblaciones silvestres se extraen ejemplares de manera ilegal para satisfacer la creciente demanda de los mercados. Lo anterior, aunado a la deforestación de su hábitat y a cambios en el uso de suelo, son las causas principales de la disminución de sus poblaciones silvestres (Szeszko, 2011). La reproducción in vitro o micropropagación vegetal es una técnica viable para la multiplicación masiva de plantas, y se usa para la propagación y preservación de orquídeas nativas importantes, como Laelia y Oncidium (Lee et al., 2007; Mengxi et al., 2011).
En un laboratorio de micropropagación comercial, el control de las condiciones físico-químicas, nutricionales y de ambiente (temperatura, humedad y luz) son factores críticos (Loberant y Altman, 2010), ya que determinan el crecimiento y el desarrollo de los tejidos vegetales. Para la fijación de carbono, las plantas usan la radiación fotosintéticamente activa (PAR) que corresponde a las zonas azul (400 a 500 nm) y roja (superior a 600 nm) del espectro visible, que abarca de 400 a 700 nm. El resto del espectro se refleja y es responsable del color verde de las hojas. Por esto, al mejorar la calidad e intensidad de la luz se acelera la fotosíntesis de las plantas, sobre todo cuando se iluminan al irradiarlas con las longitudes roja y azul del espectro (Casierra Posada et al., 2011).
La fuente de luz tradicional usada en cultivo in vitro es de lámparas tubulares fluorescentes (LTFs) (Lin et al., 2011) que emiten un espectro amplio, por lo cual los efectos fisiológicos en los vegetales son poco específicos (Da Rocha et al., 2010). Su uso en un laboratorio de cultivo de tejidos representa 65 % del costo total de electricidad (Jao y Fang, 2004) y provee las longitudes de onda innecesarias para las plantas. Por lo tanto, se debe buscar fuentes alternativas de energía y formas más eficientes de iluminar los cultivos (Loberant y Altman, 2010).
Los diodos emisores de luz (LEDs) tienen un potencial alto para usarse como fuente de luz en la micropropagación (Loberant y Altman, 2010). Sus ventajas son la conversión eficiente de energía, su volumen pequeño, una vida más larga, la emisión de radiación de longitudes de onda específicas, que permiten eficientizar la fotosíntesis (Araujo et al., 2009) y ajustar la intensidad/calidad de luz; además, sus emisiones térmicas y sus costos de mantenimiento son bajos y protegen el ambiente al disminuir las emisiones de CO2 (Lee et al., 2010). En la micropropagación, la luz LED roja y azul, solas o combinadas, tienen una influencia significativa sobre el desarrollo de las plantas (Dutta y Jatothu, 2013). La luz LED roja se asocia con el crecimiento de las plantas (alargamiento de entrenudos), como se observó en Oncidium (Mengxi et al., 2011) y con la baja concentración de pigmentos fotosintéticos probada en Dendrobium officinale (Lin et al., 2011). Pero hay poca información acerca del efecto de la luz azul sobre la fisiología de plantas superiores; con frecuencia se le relaciona con el crecimiento vigoroso, la diferenciación y los contenidos altos de clorofila y carotenoides en plantas in vitro (Lin et al., 2011; Mengxi et al., 2011).
La luz LED roja aumenta la biomasa húmeda y seca de raíces de plántulas de Paphiopedilum en comparación con la luz blanca fluorescente y la azul (Lee et al., 2011), y aumenta el peso fresco y seco de brotes en Dendrobium officinale (Lin et al., 2011). Mientras que la combinación con luz azul aumentó la acumulación de biomasa en PLBS de Oncidium (Mengxi et al., 2011). Esto indica que la respuesta de las plantas a la calidad de luz LED cambia entre especies y su etapa de desarrollo, y se necesita estudiar su efecto en cada especie para lograr determinados propósitos: promover o inhibir brotes, raíces, bulbos y controlar floración (Kim et al., 2004; Poudel et al., 2008). Con base en estas investigaciones, se plantea que es posible sustituir la luz fluorescente por luz LED durante la micropropagación de O. tigrinum y L. autumnalis; además, las respuestas morfogénicas de las plantas a la calidad de la luz, se pueden usar para el control del desarrollo de los tejidos in vitro. Por tal motivo, el objetivo del presente estudio fue determinar la calidad de luz favorable, a partir de diferentes combinaciones de luz LED, requerida para el desarrollo óptimo in vitro de plántulas de las orquídeas O. tigrinum y L. autumnalis.
Materiales y métodos
Material vegetal
En este estudio se usaron plántulas cultivadas in vitro, con una altura aproximada de 1.0 cm y tres meses de edad, obtenidas de la germinación de semillas de plantas de O. tigrinum y L. autumnalis pertenecientes al Sistema Nacional de Recursos Fitogenéticos (SINAREFI), bajo resguardo de la Facultad de Agrobiología Presidente Juárez, de la Universidad Michoacana de San Nicolás de Hidalgo, en Uruapan, Michoacán.
Medio de cultivo
Para el cultivo de las plántulas se usó el medio Murashige y Skoog (MS) sin fitohormonas más tiamina (0.4 mg L-1), inositol (100 mg L-1), sacarosa (30 g L-1) y agar (6 g L-1) ajustado a pH 5.7. Este medio se esterilizó en autoclave 15 min a 121 °C.
Establecimiento y diseño experimental
En este estudio se usaron frascos de vidrio de 100 mL de capacidad con 20 mL de medio MS, y en cada uno se colocaron 30 plántulas. El procedimiento se realizó en una campana de flujo laminar. Los frascos etiquetados se incubaron en cajas de triplay de 0.10125 m3 pintadas en su interior de color blanco. En la parte superior de cada caja se colocó una base de aluminio con cuatro focos LED de alta eficiencia de 1 W marca SiLed® de colores rojo, azul o blanco, acoplados en serie a una fuente de alimentación externa de 12 V para conectarse a la corriente alterna de 110 V.
El diseño experimental fue completamente al azar y los tratamientos fueron: LEDs rojo y azul en proporción 3:1, 2:2 y 1:3, y LEDs blancos (Cuadro 1). Las plántulas del tratamiento testigo se incubaron con una lámpara blanca fluorescente marca Phillips de 39 W, cada tratamiento se repitió seis veces, y la unidad experimental fue un frasco con 30 plántulas.
Cuadro 1 Tratamientos para evaluar el efecto de la calidad de la luz LED en el desarrollo in vitro de plántulas de Oncidium tigrinum y Laelia autumnalis.
| Tratamiento | Descripción |
| 1 | LEDs rojo y azul (3:1) |
| 2 | LEDs rojo y azul (2:2) |
| 3 | LEDs rojo y azul (1:3) |
| 4 | LEDs blancos |
| 5 (testigo) | Luz blanca fluorescente |
Las longitudes de onda emitidas por las fuentes se midieron con un espectro radiómetro de campo (Field Spec Pro) Marca ASD® (Analitical Spectral Devices). Los LEDs azules y rojos emitieron espectros caracterizados por un único pico de emisión, centrado a 458 y 636 nm, respectivamente, de aspecto gaussiano y con una dispersión (desviación estándar) de 13 nm. Los LEDs blancos presentaron un pico de emisión a 444 nm (también con una dispersión de 13 nm) y una amplia zona de emisión centrada alrededor de 553 nm (pero con una dispersión de 50 nm). En cambio, las lámparas fluorescentes presentaron una serie de picos estrechos a 436, 490, 545, 586 y 612 nm (Figura 1). La intensidad lumínica se determinó con el sensor de radiación de una pinza a fotosíntesis (IRGA-6400, LI-COR®), fue de 31±2 µmol m-2 s-1 en todos los tratamientos, con fotoperiodo de 16 h luz y 8 de oscuridad y temperatura promedio de 25 °C.

Figura 1 Características de las longitudes de onda emitidas por focos LED (azul, rojo y blanco) y lámpara fluorescente, medidas con un espectro radiómetro de campo (Field Spec Pro) Marca ASD ® (Analitical Spectral Devices). La emisión normalizada equivale al porcentaje de la energía visible total que se encuentra en cada banda de 1 nm.
Variables evaluadas
Longitud de plántula y de raíz (cm) se midió desde la base y el ápice de la plántula o la raíz, en su caso, con papel milimétrico, en 10 plántulas de cada tratamiento y repetición.
Número de hojas y raíces, también se registraron en 10 plántulas por tratamiento y repetición.
Peso fresco de plántulas (mg), se seleccionaron 10 plántulas de cada tratamiento y se pesaron en una balanza analítica digital marca OHAUS® con legibilidad de 0.01 mg.
Materia seca de plántulas (%). Esta variable se determinó en una muestra de 100 mg de tejido fresco de las plántulas de cada tratamiento y repetición, mediante el método gravimétrico. El secado se realizó en una estufa marca FELISA, a 70 °C durante 72 h. Las muestras secas se pesaron en una balanza analítica digital marca OHAUS® y los resultados se expresaron como porcentajes de materia seca en proporción a su peso fresco.
Contenido de pigmentos en las plántulas (mg g-1). La concentración de clorofila a, b y total se determinó en muestras de 100 mg de cada tratamiento y repetición, mediante su extracción de las hojas y su medición espectrofotométrica (Porra et al., 1989). Las hojas se colocaron en frascos de vidrio con 3 mL de N-Dimetilformamida cubiertos con papel aluminio, los cuales se almacenaron 48 h a 4 °C. Después se colocó 1 mL de muestra en una celda para medir su absorbancia en un espectrofotómetro JENWAY 6320D; las longitudes de onda utilizadas en la medición fueron: 664 mm para clorofila a y 647 nm para clorofila b. Los datos de absorbancia se usaron para calcular la concentración de cada una de las clorofilas con las siguientes ecuaciones: clorofila a=12.70 (A664)-2.97 (A647); clorofila b=20.70 (A647)-4.62 (A664) y clorofila total=17.90 (A647)-8.08 (A664).
Porcentaje de plantas con necrosis y albinas sólo se midió para O. tigrinum. Cada unidad experimental (frasco) se dividió en cuadrantes para facilitar los conteos, se contaron las plantas normales, necróticas o con albinismo en cada cuadrante y se sumaron para obtener los totales de cada condición por frasco. Con la suma de plantas normales, necróticas y albinas de cada frasco se obtuvo el número total de plantas por unidad experimental, que se usó como denominador para obtener los porcentajes de plantas necrosadas o albinas: porcentaje de plantas necrosadas o albinas por unidad experimental=(Número de plantas necrosadas o albinas por frasco/Número total de plantas contenidas en el frasco)×100.
El registro de variables en O. tigrinum se realizó 30 d después de iniciar los tratamientos, mientras que para L. autumnalis fue a los 90 d.
Análisis de datos
Con los datos se realizó un análisis de varianza y la prueba de Tukey (p≤0.05) con SAS versión 9.0. Para ilustrar mejor las comparaciones de medias entre tratamientos, se usó el software R versión 3.1.1, que da gráficas para las diferencias. Para comprobar la normalidad de la distribución de datos se usó la prueba Shapiro (R Core Team, 2014).
Resultados y discusión
Desarrollo de Oncidium tigrinum
Las plántulas de O. tigrinum no mostraron diferencias significativas entre tratamientos en el peso de materia fresca, el número de hojas y los contenidos de clorofila a, b y total. Aunque se esperaba que la exposición a la combinación de los espectros rojo y azul incrementara la concentración de pigmentos y peso fresco debido a que: 1) las clorofilas a y b tienen su máxima absorción en el rango de azul y rojo (Pimentel et al., 2007); 2) la luz influye considerablemente en la multiplicación celular y el crecimiento de los tejidos vegetales cultivados in vitro (Araujo et al., 2009); 3) ya se tenía registrado en plántulas de otra especie de Oncidium que la combinación de luz roja y azul (4:1 y 7:3) incrementó la concentración de pigmentos fotosintéticos (clorofila a, b y total) y la biomasa (Mengxi et al., 2011).
En nuestro estudio se detectaron diferencias significativas en longitud de plántula, número y tamaño de raíces, así como en los porcentajes de materia seca, de plantas albinas y de plantas necróticas. Las plántulas más largas (1.64 cm) se obtuvieron con combinación de luz roja y azul (1:3); pero fueron estadísticamente similares a los otros tratamientos a excepción del T2, con el cual las plántulas redujeron su tamaño en 28 % (Figura 2). El T2 también inhibió la producción y longitud de raíces (Figura 3A). El T1 sólo redujo la longitud de raíz (Figura 3B).

Figura 2 Efecto de los tratamientos de luz T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2), T3 (LEDs rojo y azul 1:3), T4 (LEDs blancos) y, T5 (luz blanca fluorescente), en la longitud de plántulas de Oncidium tigrinum cultivadas in vitro. En cada figura, letras distintas indican diferencias significativas (Tukey; p≤0.05).
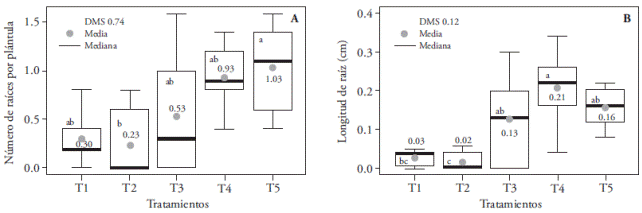
Figura 3 Efecto de los tratamientos de T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2), T3 (LEDs rojo y azul 1:3), T4 (LEDs blancos) y, T5 (luz blanca fluorescente), en el número (A) y longitud de raíces (B) de plántulas de Oncidium tigrinum cultivadas in vitro. En cada figura, letras distintas indican diferencias significativas (p≤0.05).
La menor longitud de raíces en el T1 y T2 (mayor e igual proporción de luz roja respecto a la azul), puede atribuirse a lo señalado por Correll y Kiss (2005), quienes afirman que los receptores de luz roja (fitocromo A y B) tienen efecto inhibitorio sobre la elongación de raíces. La luz blanca emite menor proporción de radiación en la región roja, estimula el enraizamiento de plántulas de Cattleya loddigessi (Araujo et al., 2009) y junto con la luz verde promueve el enraizamiento de brotes Dendranthema×grandiflora ‘Lilac Wonder’ (Miler y Zalewska, 2006). Nuestro estudio confirma que la luz blanca emitida por los LEDs y las lámparas fluorescentes promovió el enraizamiento in vitro de plántulas de O. tigrinum y estas fuentes de luz emitieron la menor cantidad de radiación en la región roja, comparado con los otros espectros estudiados.
Las plántulas micropropagadas fueron de buena calidad en todos los tratamientos de iluminación y no se presentaron cambios degenerativos en hojas y pseudobulbos (Figura 4); excepto por la presencia de algunas con ausencia total de clorofila. La incidencia mayor de plántulas albinas (33.7 %) se encontró en el T5 (luz fluorescente) y fue similar a T4 (15.8 %), pero significativamente diferente a los demás tratamientos, que en promedio mostraron 2.16 % de plántulas albinas (Figura 5). Esto parece indicar que la cantidad de luz que emiten las lámparas fluorescente y LEDs blancas en las regiones roja y azul del espectro, no es suficiente para que algunas de las plántulas de O. tigrinum realicen en forma normal la fotosíntesis.

Figura 4 Plántulas de Oncidium tigrinum in vitro iluminadas durante 30 d con diferentes combinaciones de luz: T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2), T3 (LEDs rojo y azul 1:3), T4 (LEDs blancos) y, T5 (luz blanca fluorescente).

Figura 5 Efecto de los tratamientos de luz T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2), T3 (LEDs rojo y azul 1:3), T4 (LEDs blancos) y, T5 (luz blanca fluorescente), en la incidencia de plántulas albinas de Oncidium tigrinum cultivadas in vitro. DMS: Diferencia mínima significativa. En la figura, letras distintas indican diferencias significativas (Tukey; p≤0.05).
El albinismo es una característica genética evolutiva no documentada para el género Oncidium, aunque ocurre en individuos en poblaciones de orquídeas de los géneros Epipactis (Stöckel et al., 2011) y Cephalanthera (Roy et al., 2013) que crecen bajo el dosel de los árboles. En las demás plantas verdes el proceso fotosintético es reducido debido a la poca luz ambiental o a la baja eficiencia de su aparato fotosintético o ambas (Roy et al., 2013). Además, durante las primeras etapas (formación de protocormos) las plántulas de Oncidium sphacelatum son completamente micoheterótrofas, es decir requieren de la simbiosis con un hongo micorrícico que le proporcione una fuente de carbono (Valadares et al., 2014).
El porcentaje mayor (12.2 %) de materia seca se obtuvo con el T1, estadísticamente superior de T3 al cual superó con 4 % de biomasa, pero fue similar a los otros tratamientos (Figura 6A).

Figura 6 Efecto de los tratamientos de luz T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2),T3 (LEDs rojo y azul 1:3),T4(LEDs blancos) y, T5 (luz blanca fluorescente), en la acumulación de materia seca (A) e incidencia de plántulas con necrosis (B) de Oncidium tigrinum cultivadas in vitro. En cada figura, letras distintas indican diferencias significativas (p≤0.05).
Respuestas similares se han encontrado en condiciones in vitro con intensidades de luz de 70 µmol m-2 s-1. Así, en Lilium oriental hybrid ́Pesaro ́, la iluminación con luz LED azul+roja (1:1) y luz fluorescente, incrementaron la materia seca en bulbillos (Lian et al., 2002) y en Dendrobium officinale, la luz roja+azul (1:1) mejoró la concentración de biomasa en protocormos (Lin et al., 2011). Además, la iluminación sólo con luz azul reduce la materia seca (Lian et al., 2002); por lo cual, en nuestro estudio, el valor menor registrado con el T3 se puede deber a la proporción más alta de luz azul con relación a la roja. Con el T3 se presentó mayor porcentaje de plántulas con necrosis (57.7 %), y es posible que la necrosis de tejidos se deba a daños por fotoinhibición o fotoxidación, como lo señalan Araujo et al. (2009) y Casierra-Posada et al. (2011). Sin embargo, fue similar (p>0.05) a los demás tratamientos, con excepción del T4 que disminuyó significativamente la incidencia de plántulas necrosadas. Es decir, con luz LED 100 % blanca se obtuvieron plántulas casi sin daño aparente (Figura 6B).
Desarrollo de Laelia autumnalis
En esta especie se detectaron diferencias significativas entre tratamientos en longitud de plántula y raíz, número de hojas y de raíces, en el peso fresco y seco de plántulas, así como en el contenido de clorofila a, b y total. Las plántulas más largas se obtuvieron con los tratamientos con mayor emisión de luz en la región azul: T3 (3.26 cm), T1 (3.04 cm) y T2 (2.93 cm), estadísticamente similares, pero superiores a T4 (2.65 cm) y, T5 (1.88 cm) (Figura 7A). Sin embargo, estas plantas alargadas presentaron cambios morfológicos degenerativos, con tallos y hojas más delgados que las plantas del resto de los tratamientos (Figura 8). Esto puede explicarse en parte porque la luz azul, aumenta el consumo de agua por las plantas hasta su máxima capacidad celular, lo cual puede incidir en mayor alargamiento de los tejidos (Ribeiro et al., 2009). Contrario nuestros resultados, Ribeiro et al. (2009) no encontraron diferencias en la longitud de las plántulas de Zantedeschia aethiopica por efecto de la calidad de la luz blanca, roja, azul y verde; en cambio, en brotes de Paphiopedilum. Lee et al. (2011) observaron una pérdida de dominancia apical (plantas pequeñas) por efecto de la luz azul.
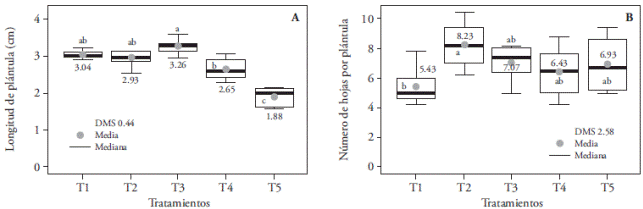
Figura 7 Efecto de los tratamientos de luz T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2), T3 (LEDs rojo y azul 1:3), T4 (LEDs blancos) y, T5 (luz blanca fluorescente), en la longitud (A) y número de hojas (B) de plántulas de Laelia autumnalis cultivadas in vitro. En cada figura, letras distintas indican diferencias significativas (p≤0.05).

Figura 8 Plántulas de Laelia autumnalis in vitro irradiadas durante 90 días con diferentes combinaciones de luz: T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2), T3 (LEDs rojo y azul 1:3), T4 (LEDs blancos) y, T5 (luz blanca fluorescente).
El número de hojas promedio de todos los tratamientos fue mayor de seis, excepto en el T1 que sólo tuvo cinco, aunque estadísticamente sólo fue diferente del T2, con 8.23 hojas por planta (Figura 7B). Este efecto pudo deberse a la baja proporción de la luz azul con relación a la roja en el T1 (1:3) en comparación con T2 (2:2). La inducción de brotes laterales y hojas en plántulas de Anthurium (Budiarto 2010) y PLBs de Dendrobium officinale (Lin et al., 2011) se estimula cuando se exponen a mayor cantidad de luz azul respecto de la roja. Sin embargo, esta opción no es viable para aumentar la producción de hojas en las plántulas de L. autumnalis debido a los cambios morfológicos degenerativos ocasionados por el exceso de luz azul.
El mayor peso de materia fresca se obtuvo con el T3 (rojo y azul 1:3) y el T2 (rojo y azul 2:2), que fueron superiores a los demás tratamientos (Figura 9A). Estos resultados son similares a los obtenidos por Wu y Lin (2012) quienes indican que la iluminación con luz roja y azul (1:1) a 50 µmol m-2 s-1 de intensidad, incrementa la acumulación de peso fresco de plántulas de Protea cynaroides; pero difieren con lo reportado en Cymbidium, donde el incremento de peso fresco de PLBs se atribuye a la luz verde (Nahar et al., 2012). La luz roja y la azul afectan la apertura estomática, pero Squeo y León (2007) consideran que sólo la azul es responsable de este mecanismo, por lo que las plantas expuestas a este espectro de iluminación consumen más agua, lo que puede ayudar a explicar nuestros resultados (mayor peso fresco con el T3).

Figura 9 Efecto de los tratamientos de luz T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2), T3 (LEDs rojo y azul 1:3), T4 (LEDs blancos) y, T5 (luz blanca fluorescente), en el peso fresco (A) y seco (B) de plántulas de Laelia autumnalis cultivadas in vitro. En cada figura, letras distintas indican diferencias significativas (p≤0.05).
Con T4 se registró el mayor porcentaje de materia seca (10.83), mayor a T1, T3 y T5 (8.83) (Figura 9B). Estos resultados son diferentes a los reportados por Lin et al. (2011), quienes indican que la acumulación de peso seco en plántulas de Dendrobium officinale se favorece con 70 µmol m-2 s-1 de luz roja y azul (1:2) y disminuye a la mitad cuando se iguala su proporción (1:1).
La luz LED blanca (T4) y el testigo (T5) incrementaron la acumulación de clorofila a (Figura 10A), clorofila b (Figura 10B) y clorofila total (Figura 11) respecto a los demás tratamientos, con hasta 16.6 mg g-1 más clorofila total (en T1). Esto puede deberse a la composición espectral de la luz blanca (400-700 nm), que incluye las longitudes de onda azul (430480 nm), verde (495-570 nm) y roja (590-670 nm) que absorben los fotosistemas II (≤680 nm) y I (≤700 nm), pues para una máxima eficiencia deben participar ambos fotosistemas (Solarte et al., 2010). Hay que considerar que las hojas absorben fotones principalmente de los espectros azul y rojo, y de manera más débil lo hacen en el verde, aunque la mayoría de esos fotones son reflejados por las plantas en forma de radiación difusa (Lazo y Ascencio, 2010).

Figura 10 Efecto de los tratamientos de luz T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2), T3 (LEDs rojo y azul 1:3), T4 (LEDs blancos) y, T5 (luz blanca fluorescente), en la acumulación de clorofila a (A) y clorofila b (B) de plántulas de Laelia autumnalis cultivadas in vitro. En cada figura, letras distintas indican diferencias significativas (p≤0.05).
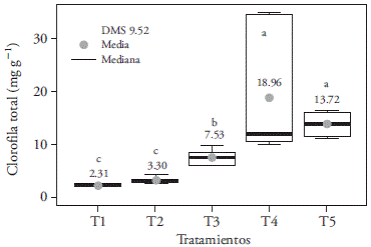
Figura 11 Efecto de los tratamientos de luz T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2), T3 (LEDs rojo y azul 1:3), T4 (LEDs blancos) y, T5 (luz blanca fluorescente), en la acumulación de clorofila total de plántulas de Laelia autumnalis cultivadas in vitro. En cada figura, letras distintas indican diferencias significativas (p≤0.05).
Como en nuestro estudio, Lee et al. (2011) observaron una disminución en el contenido de clorofila total de hojas de Paphiopedilum bajo condiciones de luz azul, roja y roja más azul (9:1); mientras que Lin et al. (2011), en Dendrobium officinale, encontraron que el espectro azul incrementó más del doble la concentración de pigmentos fotosintéticos.
El número mayor de raíces por plántula (0.90) se observó con la luz fluorescente (T5) y fue similar a la luz LED blanca (T4); mientras que las combinaciones de luz roja y azul (3:1, 2:2 y 1:3) no indujeron la formación de raíces (Figura 12). Esta inhibición de la rizogénesis puede estar asociada con un incremento en la emisión de luz azul de los espectros en T1 (3:1), T2 (1:1) y T3 (1:3). Con esta tendencia coinciden los resultados obtenidos en plántulas de Chrysanthemun (Kurilčik et al., 2008), donde se inhibe la formación de raíces cuando aumenta la luz azul con respecto a la roja y roja lejana; aunque difieren con lo observado en Paphiopedilum, donde el desarrollo de raíces se dio con luz roja y azul, con pesos frescos y secos de hasta 387.7 y 31.2 mg más que la luz blanca, respectivamente (Lee et al., 2011).
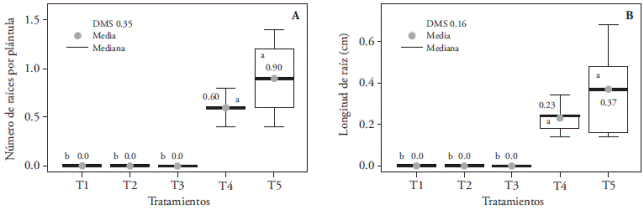
Figura 12 Efecto de los tratamientos de luz T1 (LEDs rojo y azul 3:1), T2 (LEDs rojo y azul 2:2), T3 (LEDs rojo y azul 1:3), T4 (LEDs blancos) y, T5 (luz blanca fluorescente), en el número (A) y la longitud (B) de raíces de plántulas de Laelia autumnalis cultivadas in vitro. En cada figura, letras distintas indican diferencias significativas (p≤0.05).
La luz azul reduce el enraizamiento in vitro de brotes de Protea cynaroides porque estimula la biosíntesis de compuestos fenólicos; además, en combinación con luz roja incrementa la acumulación de ácido 3,4-dihidroxibenzoico, ácido gálico y ácido ferúlico. Al contrario, la formación de raíces en las plántulas iluminadas con LEDs rojos es el resultado de la baja concentración endógena de estos fenoles (Wu y Lin, 2012). Durante la iluminación simultánea con luz azul y roja, ocurre una interacción sinérgica entre criptocromos y fitocromos (fotorreceptores del azul y rojo, respectivamente) y estos pigmentos pueden ser responsables de la percepción y activación del proceso de rizogénesis. Los fitocromos posiblemente regulan el proceso de rizogénesis vía sistemas de fitohormonas (Kurilčik et al., 2008).
Con base en nuestros resultados, es factible iluminar cultivos in vitro de O. tigrinum y L. autumnalis con luz LED blanca, porque produce efectos sobre el desarrollo similares a los obtenidos con luz blanca fluorescente. Además, el calor que emiten las lámparas LED es bajo y se pueden colocar directamente sobre las plantas (Schroeter-Zakrzewska y Kleiber, 2014). Esta solución permite instalar varios niveles en un estante, incrementar el número de plántulas por unidad de área y reducir los costos de energía por iluminación y enfriamiento. Pero aún se debe estudiar el efecto del fotoperiodo y la densidad de flujo fotónico fotosintético, que dependen de la especie en cuestión y es una adaptación ecológica particular (Kurilčik et al., 2008). Por ejemplo, para el desarrollo de plántulas de fresa se requiere 60 µmol m-2 s-1 (Nhut et al., 2003) y en explantes de crisantemo sólo 40 µmol m-2 s-1 (Kurilčik et al., 2008).
Conclusiones
El espectro de iluminación emitido por LEDs blancos fue el más favorable para el desarrollo in vitro de plántulas de O. tigrinum y L. autumnalis. Por lo tanto, en un laboratorio de micropropagación de orquídeas es posible reemplazar las lámparas de luz blanca fluorescente por LEDs blancos, ya que ambos tipos de iluminación son altamente efectivos como fuentes de radiación fotosintética y promueven en las plantas respuestas fotomorfogénicas similares. La luz blanca reduce la longitud y la materia fresca de las plántulas de L. autumnalis, pero aumenta el contenido de pigmentos fotosintéticos y promueve su enraizamiento. Mientras que la iluminación de plántulas de O. tigrinum con mayor proporción de luz roja con respecto a la azul inhibe el enraizamiento, pero disminuye la formación de plantas albinas.

