











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El enraizamiento de estacas es el medio de propagación vegetativa y producción masiva más importante. En árboles permite transferir todo el potencial genético del material donante y generar uniformidad genética en los individuos obtenidos, entonces se captura y aprovecha la varianza genética total y se mantiene la ganancia genética mediante el mejoramiento genético (Zalesny et al., 2003). Además el método tiene costo bajo, es rápido y sencillo (Capuana et al., 2000; Husen, 2004; Tarragó et al., 2005) y contrasta con la propagación in vitro, que requiere inversión mayor en infraestructura y equipo de laboratorio, medios de cultivo y protocolos especiales de trabajo (en particular con especies forestales). El enraizamiento de estacas consiste en obtener segmentos de ramas de un árbol y mantenerlas en condiciones ambientales favorables para inducir rizogénesis y organogénesis (Thanuja et al., 2002; Husen y Pal, 2003; Liu et al., 2012). En Chile, España, Portugal, China, Canadá, Brasil, Australia, Nueva Zelanda y EE.UU. este método se utiliza para desarrollar plantaciones clonales comerciales de Pinus radiata D. Don, Pinus taeda L., Gmelina arborea Roxb., Populus, Platanus, Salix y Eucalyptus (Bedon et al., 2011).
La capacidad de enraizamiento tiene relación directa con el estado de madurez de la planta madre, y la facilidad para formar raíces secundarias disminuye con la edad del material a propagar (Klein et al., 2000). Para obtener brotes juveniles con capacidad mayor para enraizar se deben realizar podas (Ruíz et al. 2005; Trobec et al., 2005; Castillo et al., 2013). El tejido juvenil responde mejor a los reguladores de crecimiento que el de células viejas y totalmente diferenciadas, probablemente porque las zonas del genoma que controlan la formación de órganos están menos reprimidas y pueden desbloquearse con los tratamientos con auxinas (Druege y Kadner, 2008; Klopotek et al., 2010; Denaxa et al., 2012).
Los factores genéticos y el estado fisiológico de las plantas controlan la inducción de raíces y los factores ambientales determinan su elongación (Thomas y Schiefelbein, 2002; Castrillón et al., 2008). El éxito del enraizamiento de las estacas depende de la especie, la edad de la planta donadora, la sección de la rama de la que se extraigan las estacas, el medio de enraizamiento y los reguladores de crecimiento empleados (Itoh et al., 2002; Klopotek et al., 2012).
Entre los factores fisiológicos principales están las auxinas endógenas que son traslocadas del meristemo apical hacia la base de la planta (Ruíz et al., 2005). En algunas especies estos compuestos endógenos no son suficientes para inducir enraizamiento y se deben aplicar auxinas exógenas. La auxina más usada es el ácido indolbutírico (AIB) y es necesario conocer la dosis adecuada (Ruíz et al., 2005), la que se relaciona con el contenido endógeno, y, a la vez, con la posición original de la estaca en la planta.
Respecto a los factores ambientales, el tipo de sustrato que dará soporte y adherencia a las raíces se debe considerar. La formación de raíces requiere cantidades altas de oxígeno, por lo que los sustratos deben tener suficiente aire, permitir el intercambio de gases, tener drenaje, temperatura y pH adecuados y libres de patógenos (Bonfil et al., 2007; Saranga y Cameron, 2007).
En México Cedrela odorata L. (cedro rojo) es una especie prioritaria por su importancia industrial (Sánchez et al., 2003; Ramírez et al., 2008; Quinto et al., 2009), y la más utilizada para plantaciones comerciales (CONAFOR, 2015). Pero Hypsipyla grandella (Zeller) puede dañar su desarrollo y limitar el éxito de las plantaciones comerciales (Navarro et al., 2004). Árboles resistentes a la invasión de esta palomilla (Sánchez et al., 2003) se detectaron y son candidatos para reproducirlos masivamente. El enraizamiento de estacas es una opción recomendable para multiplicar genotipos con características superiores (Marrón et al., 2002; Kraiem et al., 2010), pero no hay un protocolo para el enraizamiento de estacas de C. odorata.
Así, con la finalidad de desarrollar una metodología de propagación clonal mediante enraizamiento de estacas, se evaluó el efecto del ácido indolbutírico (AIB), la posición de la estaca en la planta y el tipo de sustrato en el enraizamiento de estacas juveniles de C. odorata en condiciones controladas de humedad y temperatura. La interacción tipo de sustrato y AIB favorecerá, en diferente proporción, la producción de raíces adventicias según el tipo de estaca.
Materiales y métodos
Localización y material vegetal
Para la producción de planta se usó semilla recolectada en mayo del 2012 de la familia 99 del huerto semillero ubicado en el Campo Experimental El Palmar, Veracruz, del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. La siembra se realizó en abril de 2013 en un invernadero del Colegio de Postgraduados, Texcoco, Estado de México. El sustrato fue una mezcla de turba, agrolita y vermiculita (2:1:1) en tubetes de polietileno de 1 L, y 10 g de fertilizante de liberación controlada por tubete. La germinación se inició a los 15 d, finalizó a los 35 d y alcanzó 95 %. Después se aplicaron riegos frecuentes y fungicida (captan® 1 g L-1) cada mes a la planta.
Producción de miniestacas
Para estimular la formación de material juvenil se realizaron podas a los 6 y 12 meses, a 6cm sobre el cuello de la planta. Para evaluar la incidencia de la posición de la estaca en el enraizado de cada brote se separaron las secciones apical, intermedia y basal. La longitud de los brotes seleccionados fue mayor a 25 cm y diámetro de 1 cm. Las hojas de cada vareta se cortaron y se obtuvieron tres estacas de 7 cm de longitud.
Preparación del sustrato
Los sustratos fueron: agrolita y la mezcla de turba, agrolita y vermiculita (2:1:1). Estos se esterilizaron 40 min con vapor de agua, se enfriaron y colocaron en la cámara para enraizamiento.
Establecimiento del experimento
En una cámara de enraizamiento semiautomática (2.7x1.2x0.7 m de longitud, anchura y altura) se controló el riego y la temperatura. El riego se programó para distribuir 2250 mL de agua destilada en 10 riegos por día. La humedad relativa se mantuvo sobre 80 % y la temperatura entre 18 y 25 °C en el día, y si llegaba a 25 °C un ventilador automático enfriaba a 18 °C la cámara.
Como testigo de asepsia, las estacas se colocaron 15 min en una solución de Captán® al 5 %. Para determinar las mejores dosis para enraizamiento se realizó una inmersión por 5 s en soluciones con 1500, 5000 y 10 000 ppm de Radix®, y como testigo se utilizó talco industrial.
Los sustratos se humedecieron con agua destilada y en hoyos de 4 cm de profundidad y espaciamiento de 10x10 cm se colocaron las estacas presionando el sustrato alrededor de la estaca, sin dañar los tejidos.
Diseño experimental y evaluación de variables
El diseño experimental fue completamente al azar, con arreglo factorial 2x3x4: dos tipos de sustratos, tres posiciones de corte de la estaca y cuatro concentraciones de AIB. Hubo diez repeticiones (10 estacas por tratamiento, 240 estacas en total). El experimento se realizó en el periodo otoño-invierno (del 26 de agosto de 2014 al 13 de enero de 2015), con evaluaciones periódicas se monitoreó la presencia de patógenos, y 20 semanas después se evaluó el porcentaje de estacas vivas, estacas enraizadas por tratamiento, número y longitud de raíces de primero y segundo orden por estaca.
Previo al análisis de varianza, la variable en porcentaje (Y) se transformó con la función arcoseno de la raíz cuadrada del valor original expresado en fracción decimal
Para obtener los estimadores de varianza y del error para cada variable se realizó el análisis estadístico con el modelo lineal siguiente:
donde: Y ijkl es la variable respuesta del i-ésimo tipo de estaca, con la j-ésima dosis de AIB, en el k-ésimo tipo de sustrato; μ es el promedio general; E i es el efecto fijo del i-ésimo tipo de estaca (apical, intermedia y basal); D j es el efecto fijo de la j-ésima dosis de AIB (0, 1500, 5000 y 10 000 ppm); S k es el efecto fijo del k-ésimo tipo de sustrato (mezcla turba-agrolita-vermiculita 2:1:1 vs. agrolita); ED ij es la interacción entre la i-ésimo tipo de estaca y la j-ésima dosis de AIB; ES ik es el efecto de la interacción entre el i-ésimo tipo de estaca y del k-ésimo tipo de sustrato; DS jk es el efecto de la interacción entre la j-ésima dosis de AIB y del k-ésimo tipo de sustrato; EDS ijk es la interacción entre el i-ésimo tipo de estaca, la j-ésima dosis de AIB y el k-ésimo tipo de sustrato; y E ijkl es el error aleatorio correspondiente a la observación Y ijkl .
Resultados y discusión
El efecto del sustrato, dosis de AIB y tipo de estaca fue significativo (p≤0.05) en la supervivencia, el enraizamiento y la morfología de las estacas, pero no hubo efecto en sus interacciones (Cuadro 1).
Cuadro 1 Valores de probabilidad (p) en el análisis de varianza de las variables de enraizamiento en estacas de Cedrela odorata L.
| Fuente de variación | Superv. (%) | Estacas enraizadas (%) | Número de raíces | Longitud de raíces | ||
| Prim. | Secun. | Prim. | Secun. | |||
| Sustrato | 0.0001† | 0.0001† | 0.4196 | 0.4462 | 0.0012† | 0.0749 |
| Dosis de AIB | 0.0316† | 0.0454† | 0.0141† | 0.0079† | 0.0707 | 0.4370 |
| Tipo de estaca | 0.0072† | 0.0453† | 0.0327† | 0.0073† | 0.0037† | 0.0894 |
| Sustrato*Dosis | 0.9793 | 0.8502 | 0.8174 | 0.5978 | 0.0786 | 0.2898 |
| Sustrato*Estaca | 0.6807 | 0.6850 | 0.1459 | 0.0841 | 0.1377 | 0.0043† |
| Dosis*Estaca | 0.2164 | 0.0513 | 0.8958 | 0.9438 | 0.9267 | 0.6807 |
| Sustrato*Dosis*Estaca | 0.7858 | 0.7998 | 0.3844 | 0.4060 | 0.1985 | 0.3051 |
†Diferencias significativas (p≤0.05). Superv: supervivencia, Prim: raíces de primer orden, Secun: Raíces de segundo orden.
Efecto del sustrato
Los mayores porcentajes de supervivencia y de estacas enraizadas se obtuvieron con agrolita (Cuadro 2). La agrolita incrementó el enraizamiento y la supervivencia, respecto a la mezcla (turba-agrolitavermiculita 2:1:1), en proporción dos a uno. En la mezcla la longitud en las raíces de primer orden fue mayor que en la agrolita, en proporción dos a uno.
Cuadro 2 Valor promedio y error estándar para cada factor y tratamiento en el enraizamiento de estacas de Cedrela odorata L. a las 20 semanas.
| Factor | Superv. (%) | Estacas enraizadas (%) | Número de raíces | Longitud de raíces (mm) | ||
| Primarias | Secundarias | Primarias | Secundarias | |||
| Sustratos | ||||||
| Mezcla | 38.3±4.8 b | 32.5±4.7 b | 14.9±6.6 a | 11.8±1.7 | 684±66 a | 73±35 a |
| Agrol. | 82.5±3.9 a | 79.1±4.1 a | 12.1±2.0 a | 10.2±1.5 | 347±68 b | 59±28 a |
| CV (%) | 8.4 | 8.7 | 10.5 | 8.6 | 8.6 | 4.8 |
| Dosis de AIB (ppm) | ||||||
| 0 | 66.6±3.8 b | 63.3±4.9 b | 4.8±3.46 b | 5.2±1.7 b | 356±237 | 62±23 a |
| 1500 | 75.0±3.6 a | 73.3±3.6 a | 14.2±2.7 ab | 11.5±3.8 ab | 640±205 | 71±39 a |
| 5000 | 48.3±9.3 c | 43.3±.4.7 c | 21.2±3.4 a | 16.4± 1.4 a | 538±178 | 67±32 a |
| 10 000 | 41.6±4.6 c | 31.6±4.8 c | 11.1±2.5 ab | 9.5±1. ab | 292±202 | 53±28 a |
| CV(%) | 9.1 | 9.5 | 9.6 | 7.9 | 8.9 | 4.9 |
| Tipo de estaca | ||||||
| Apical | 68.7±4.5 a | 66.2±4.8 a | 21.3±5.5 a | 16.7±4.8 a | 705±149 a | 75±09 a |
| Inter. | 55.0±5.3 b | 52.5±8.1 b | 8.9±4.5 b | 7.1±3.3 b | 390±165 b | 60±08 a |
| Basal | 35.0±11.8 c | 32.5±4.8 c | 4.4±2.6 b | 5.3±2.8 b | 148±156 b | 50±16 a |
| CV (%) | 9.1 | 9.5 | 9.1 | 7.2 | 8.0 | 4.7 |
| Prom. | 56.7 | 52.7 | 12.5 | 10.4 | 4.55 | 0.63 |
Letras diferentes en una columna, para cada factor, indican diferencias estadísticamente significativas (p≤0.05). Superv: supervivencia, mezcla: turba-agrolita-vermiculita 2:1:1 v/v, Agrol: agrolita, ppm: partes por millón, CV: coeficiente de variación, Inter: intermedia, Prom: promedio general
El mejor enraizamiento en agrolita se debe a sus características físicas, como porosidad, aireación, drenaje y pH (Saranga y Cameron, 2007). Según Bonfil et al. (2007), un sustrato debe suministrar humedad continua, mantener temperatura moderada y permitir aireación adecuada. La agrolita muestra estas características, por lo que favorece el enraizamiento de Pinus leiophylla Schiede ex Schltdl. et Cham. (Cuevas et al., 2015), Pinus pinaster Ait. (Majada et al., 2010), Pinus elliottii Engelm. var. elliottii Little & Dorman, Pinus caribea var. hondurensis Barr. & Golf. (Hunt et al., 2011), Juniperus scopulorum Sarg. y Thuja occidentalis L. (Bielenin, 2003).
Efecto de la dosis de AIB
Mil quinientas ppm de AIB aumentaron 10 % el enraizamiento respecto al testigo; dosis mayores lo disminuyeron, probablemente por toxicidad. Según Mesén et al. (1997), algunas especies forestales no toleran concentraciones mayores a 2000 ppm; es el caso de Eucalyptus, Cupressus lusitánica Mill., Alnus acuminata Kunth y Tectona grandis L. (Husen y Pal, 2003). Con 1500 ppm de AIB el número de raíces de primero y segundo orden fue tres veces y el doble que en el testigo. La longitud de las raíces de primer y segundo orden no cambió (p≤0.05) con el AIB, respecto al testigo. Pero en la supervivencia de estacas hubo más de 30 % de diferencia (p≤0.05) con el mejor y peor tratamiento.
Con la dosis de 5000 ppm el número de raíces de primero y segundo orden fue cinco y tres veces mayor (p≤0.05) que en el testigo. Esto se debió a que después de que la estaca enraíza dosis crecientes de AIB aumentan el transporte de sustancias hacia la base de la estaca, con lo que aumenta el desarrollo de más raíces (Mesén et al., 1997).
Con 10 000 ppm se obtuvo el porcentaje menor de estacas enraizadas (Cuadro 2), atribuible a la intoxicación por auxinas. Las estacas sin AIB tuvieron dos veces más enraizamiento que con 10 000 ppm; esto indica que las estacas contienen auxinas endógenas en cantidad suficiente para promover el enraizamiento.
Las auxinas promueven el desarrollo de raíces adventicias en el enraizamiento de estacas (Bielenin, 2003; Castrillón et al., 2008; Kumar et al., 2011). Castillo et al. (2013) recomendaron el uso de AIB para especies en las que el enraizamiento se califica como difícil. En el porcentaje de estacas enraizadas hubo diferencias significativas (p≤0.05) entre las dosis de AIB. Las auxinas estimularon la actividad del cámbium y la movilización de reservas hacia el sitio de iniciación de raíces (Dhillon et al., 2011). El AIB mejoró el enraizamiento en Pinus jaliscana Pérez de la Rosa. (Aparicio et al., 2006), Viccinium meridionale Swartz (Castrillón et al., 2008), Taxus globosa Schltdl. (Muñoz et al., 2009), Olea europaea L. (Denaxa et al., 2012) y Gmelina arborea Roxb. (Ruíz et al., 2005).
Efecto de la posición de las miniestacas
Los porcentajes de supervivencia y enraizado fueron 20 y 50 % superiores en las estacas apicales que en las intermedias y las basales (Cuadro 2). Las estacas apicales favorecieron dos veces más cantidad de raíces de primero y segundo orden que las intermedias y cuatro veces más que las basales. Además, las apicales tuvieron longitud de raíces de primer orden dos veces mayor que las intermedias del brote, que fue la segunda mejor. En la longitud de raíces de segundo orden no hubo diferencias.
La estaca apical presentó el enraizamiento mejor, que podría deberse a alguna condición fisiológica o anatómica asociada con la presencia de yemas. De acuerdo con Hartmann et al. (2013) las estacas apicales enraizan mejor que las laterales, porque las yemas apicales son productoras de auxinas abundantes y estimulan el desarrollo radical (Moore, 1984). Ruíz et al. (2005) señalaron que el grado de juvenilidad, la lignificación del tejido, el contenido de reservas y en particular el nivel endógeno de fitohormonas son factores involucrados en la capacidad mayor de enraizado de estacas apicales de Gmelina arborea.
En algunos casos las estacas basales enraizan mejor, como en Populus con el doble, respecto a las estacas intermedias y apicales (Zalesny et al., 2003). Las estacas basales de P. leiophylla tienen 3.5 más probabilidad de éxito en el enraizamiento (Cuevas et al., 2015). La causa de esta variación entre especies se desconoce y se debe investigar. En nuestro estudio las estacas apicales tuvieron respuesta mejor de enraizamiento, pero no se descarta usar estacas basales para propagación de clones selectos de C. odorata. La experimentación debería continuar para aumentar los porcentajes de enraizamiento de estacas basales y se aprovecharía la mayoría del material para maximizar la producción de planta.
Efecto de la interacción entre factores
Sólo la interacción del sustrato por el tipo de estaca mostró efecto significativo en la longitud de raíces de segundo orden (Cuadro 1). La combinación de la estaca apical con el sustrato (turba-agrolita-vermiculita 2:1:1) propició la longitud mayor de raíces de segundo orden, 96±11 mm. La longitud de la raíz decreció gradualmente con el tipo de la estaca, mayor en las apicales a menor en las basales; la reducción fue mayor en la longitud de raíces en la mezcla (estacas de posición intermedia; Figura 1).
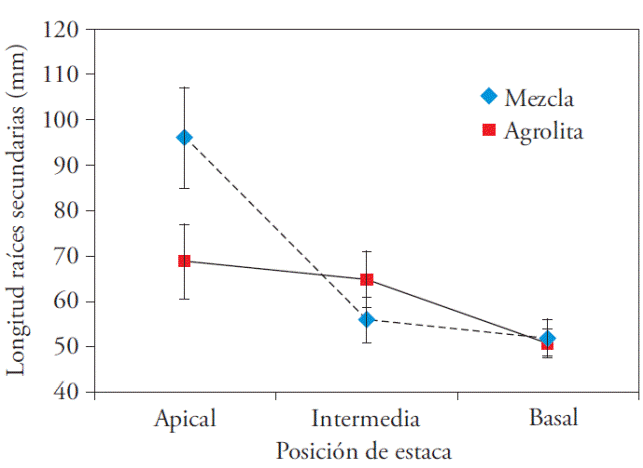
Figura 1 Interacción de la posición de la estaca y el sustrato en la longitud de raíces secundarias. Cada punto representa el promedio con la desviación estándar.
La longitud de las raíces inducidas en las estacas puede indicar la velocidad de enraizamiento, las más largas tuvieron menos tiempo para enraizar (Ruiz et al., 2005). En nuestro estudio, probablemente las raíces secundarias de las estacas apicales aparecieron primero en el sustrato de la mezcla que en la agrolita. Esto podría ser una ventaja porque disminuiría la estadía de las estacas en el módulo de propagación, los riesgos de contaminación y los costos de mantenimiento. Pero este sustrato todavía no se recomienda porque se desconoce si la longitud de raíces de las estacas establecidas en agrolita pueden alcanzar el tamaño que en la mezcla en una o dos semanas.
Estos resultados indican que la propagación vegetativa de C. odorata mediante el enraizamiento de estacas tiene gran potencial. Más investigación sería necesaria para determinar si el porcentaje de enraizamiento mayor se logra en menos tiempo. También podrían evaluarse otras concentraciones de AIB, alrededor de los 1500 ppm, otros clones de calidad superior y secciones apicales.
Conclusiones
La agrolita, como sustrato, favorece el enraizamiento y supervivencia de las estacas. Dosis de 1500 ppm de AIB incrementa la capacidad de enraizamiento de las estacas; pero, dosis mayores la inhiben. La posición apical de las estacas favorece la supervivencia y enraizado, con raíces de primero y segundo orden. Así, la combinación mejor para el enraizamiento es la posición apical y 1500 ppm de AIB.

