











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Hasta ahora no se conoce una función metabólica específica del yodo en las plantas terrestres (Kabata-Pendias, 2011). Sin embargo, el yodo induce en las plantas una mayor presencia de antioxidantes y aporta un aumento en la tolerancia a algunos factores adversos (Blasco et al., 2008; Blasco et al., 2011; Gupta et al., 2015). En especies marinas como Laminaria digitata (kelp) el proceso de inducción de tolerancia está relativamente bien entendido, y la acumulación celular o volatilización hacia la atmósfera de este elemento está relacionado con el nivel de estrés oxidativo. Bajo condiciones de producción alta de especies reactivas del oxígeno (ROS) el yodo es volatilizado, mientras que con baja concentración de ROS el yodo es almacenado para estar disponible si aumenta el estrés (Kupper et al., 2008). Venturi (2011) propuso la hipótesis de que el yodo fue uno de los primeros antioxidantes usados por los organismos fotosintéticos. Según La Barre et al. (2010), las algas marinas usan yodo como un antioxidante durante estrés oxidativo, lo cual pudiera ser similar en las plantas terrestres. Sin embargo, el uso del yodo ha sido poco investigado en la práctica agrícola.
Los efectos positivos de la aplicación de yodo en las plantas terrestres se mantienen al usarlo en bajas concentraciones, pero ocurre toxicidad al aplicar en las plantas más allá de ciertas concentraciones (Caffaggni et al., 2011; Landini et al., 2011). Estos umbrales de beneficio y de toxicidad son variables y dependen de la especie (Kabata-Pendias, 2011).
En México, el pimiento morrón (Capsicum annuum L.) es el cuarto cultivo con mayor relevancia (SIAP, 2010). La aplicación de yodo en este cultivo pudiera tener ventajas como aumento en la concentración de yodo en las plantas, para propósitos de biofortificación y mejora en su capacidad de tolerancia frente al estrés por el aumento en la capacidad antioxidante. Sin embargo, no hay información publicada acerca de las respuestas del pimiento a las aplicaciones de este elemento. Por lo tanto, se requieren estudios para conocer las respuestas de crecimiento y calidad nutricional de la planta a la aplicación de yodo.
El objetivo de este estudio fue determinar la factibilidad de uso del yodo por vía foliar para promover el crecimiento de las plántulas de pimiento morrón.
La hipótesis fue que la aplicación de este elemento aumentará la tasa de crecimiento y el contenido de antioxidantes en las plántulas.
Material vegetal y condiciones de crecimiento
El experimento se realizó en el campus Montecillo del Colegio de Postgraduados, Montecillo, Estado de México. Los almácigos para la germinación de la semilla y el crecimiento de las plántulas se establecieron en cámara de ambiente controlado con temperatura de 23 a 25 °C y humedad relativa de 50 %, con periodo de luz de 14 h (pares fluorescentes) y 10 h de obscuridad. Para los almácigos se emplearon charolas de plástico de 50 cavidades. Cada charola se llenó con el sustrato peat-moss Sungro®, se colocó una semilla por cavidad y se cubrió con una fina capa de vermiculita. La variedad de pimiento morrón sembrada fue Sidenca de la compañía Enza Zaden. Los riegos se realizaron diario usando una regadera manual para mantener la humedad del sustrato. La nutrición se realizó con solución de Steiner al 25 % [0.147 mM Ca(NO3)2, 0.034 mM KH2PO4, 0.076 mM K2SO4, 0.176 mM de KNO3, 0.068 mM MgSO47 H2O, micros, pH 5.5]; inició al presentarse las primeras hojas verdaderas; 7 d después la concentración aumentó a 50 % y 7 d después a 75 % hasta el trasplante.
Aplicación foliar de yodo
Cuando la plántulas presentaron las primeras hojas verdaderas se realizaron tres aplicaciones foliares de yodo, con intervalos de 14 d. Los tratamientos fueron 10, 15, 30, 45 y 50 μM KI L-1 y un testigo (0 yodo), con tres repeticiones. Las aspersiones foliares se hicieron con un aspersor de ultra bajo volumen fuera de la cámara de crecimiento para evitar contaminación entre tratamientos. La unidad experimental fue una charola con 50 cavidades, y el diseño experimental fue completamente al azar.
Medición de variables durante el desarrollo del cultivo
Los muestreos se hicieron 7 d después de cada aplicación foliar del yodo, recolectando 10 plántulas por unidad experimental para medir: altura de planta (cm), desde el punto cero sobre la superficie del sustrato hasta el ápice; diámetro de tallo (mm) con un vernier digital marca Truper; área foliar (cm2) por el método de planimetría (escaneo y análisis de imágenes y estimación de la longitud y anchura de la hoja), por el método reportado en Melgarejo et al. (2011). Las plántulas se secaron 72 h en un horno de circulación continua a 70 °C y se pesaron para obtener peso seco en g.
Determinación de índices de crecimiento
Los datos de materia seca (MS) y área foliar se usó para determinar de los índices de crecimiento del pimiento: tasa de asimilación neta (TAN= LnA2-LnA1/t2-t1); tasa de crecimiento absoluto (TCA= P2-P1/t2-t1); tasa de crecimiento relativo (TCR= LnP2-LnP1/t2-t1); área foliar específica (AFE= AF/ biomasa de hoja). Ln A= logaritmo natural de área foliar (cm2); t= tiempo (d); P= peso seco total (g); Ln P= logaritmo natural de MS total (g) (Amaro et al., 2004).
Determinación de la actividad antioxidante total (método del DPPH)
La actividad de antioxidantes totales se determinó por el método del DPPH (Scherer y Texeira 2009). La reacción se basa en una disminución de color que ocurre cuando un electrón de un átomo de nitrógeno en DPPH es reducido por recibir un átomo de hidrogeno de los compuestos antioxidantes. En 10 repeticiones por tratamiento se maceró el material vegetal, ya homogenizado se pesaron 100 mg y se colocaron en frascos de 2 mL. Se agregaron 1.5 mL de etanol al 60 % y reposaron 24 h, las muestras se centrifugaron 15 min a 9 000 rpm, se tomaron 400 mL del sobrenadante de cada muestra y se colocaron en un frasco de 2 mL para agregar 600 mL de metanol al 80 % y 1 mL de la solución de DPPH. Las muestras reposaron 15 min y la lectura se hizo en el espectrofotómetro a 517nm. La lectura de cada muestra se repitió a los 30 y 60 min. Los resultados se expresan como % del DPPH inhibido.
Contenido de yodo y minerales en plántulas
El contenido de yodo se midió por la técnica de cenizas alcalinas (Fisher et al. 1986). Las 10 plántulas de pimiento muestreadas por tratamiento se colocaron 72 h en horno de circulación continua a 70 °C, se molieron y pesaron 0.5 g del tejido vegetal seco. Las muestras se colocaron en crisoles de porcelana, se agregóKOHyKNO3,y se pusieron 3h en una mufla a 580°C,se enfriaron a temperatura ambiente y las cenizas se pasaron a tubos cónicos para ser extraídas con KOH. Las cenizas de los tubos se centrifugaron 15 min a 7 200 rpm. Luego, 1 mL del sobrenadante se decantó y aforó a 10 mL con KOH 2M y se hicieron las lecturas para cuantificar de yodo en el equipo de espectroscopia de emisión atómica de inducción por plasma (ICP-OES, Varian 725-OES, Australia).
Elcontenidode P, K, Ca, Mg, S, B, Cu ,Fe, Mn y Zn en la parte aérea, se analizó mediante la técnica de digestión húmeda y se leyó en el ICP, y el N se determinó con el método Kjeldahl (Bremner, 1965).
Resultados y discusión
Medición del desarrollo del cultivo
Las plántulas de pimiento morrón mostraron una respuesta significativa (p≤0.05) a las aplicaciones foliares de yodo en altura de planta, diámetro de tallo, área foliar y MS para cada tratamiento. La altura aumentó con la aplicación foliar de yodo (10 μM L-1) y fue diferente los otros tratamientos (Cuadro 1; p≤0.05). Los resultados muestran que al aumentar la concentración de yodo disminuyó la altura de la plántula y en las concentraciones más altas se observaron los bordes de las hojas con síntomas de toxicidad. La respuesta positiva en la altura de la planta podría ser por un efecto antioxidante del yodo, lo cual permitiría a la planta una mayor adaptabilidad al entorno de crecimiento. Este efecto antioxidante fue mencionado por Venturi (2011), pero depende de la concentración de yodo en el medio de crecimiento ya que en concentración alta es tóxico (Caffaggni et al., 2011; Landini, et al., 2011). La misma respuesta se encontró en cebolla (Allium cepa L.), zanahoria (Daucus carota L.), apio (Apium graveolens L.) y espinaca (Spinacea oleracea L.), al aplicar más de 5 μM L-1 de yodo, y con 40 μM L-1 o más las plantas fueron más pequeñas y con síntomas de toxicidad (Jiu et al., 2004).
Cuadro 1 Efecto de seis dosis de KI aplicadas al follaje de plántulas de pimiento morrón sobre la altura de plántula, diámetro de tallo, área foliar y producción de materia seca.
| Aplicación foliar | Altura de plántula (cm) | Diámetro de tallo (mm) | Área foliar (cm 2 ) | Materia seca (g) |
|---|---|---|---|---|
| Testigo 0 μM | 12.81 c | 2.319 c | 68.43 c | 0.92 d |
| Yodo 10 μM | 17.63 a | 3.056 a | 93.06 a | 1.86 a |
| Yodo 15 μM | 14.06 b | 2.675 b | 74.79 b | 1.23 c |
| Yodo 30 μM | 14.03 b | 2.713 b | 76.82 b | 1.25 c |
| Yodo 45 μM | 14.55 b | 2.790 b | 78.61 b | 1.28 c |
| Yodo 50 μM | 13.02 c | 2.073 d | 55.59 d | 1.52 b |
| R2 | 0.8275 | 0.8486 | 0.9474 | 0.9492 |
| C.V. | 5.2535 | 5.5623 | 5.2077 | 5.4078 |
| DMS | 0.6627 | 0.2175 | 6.242 | 0.0992 |
Medias con diferente letra son estadísticamente diferentes (p≤0.05). ALP: Altura de plántulas; DIAT: Diámetro de tallo de pántulas; AF: Área foliar total de plántulas; MS: Acumulación de materia seca en plántulas; R2: Coeficiente de determinación; C.V.: Coeficiente de variación; DMS: Diferencia mínima significativa
El diámetro de tallo aumentó hasta 32 % respecto al testigo con la aplicación foliar de 10 μM L-1 (Cuadro 1) y 20 % con 45 μM L-1. El tallo más delgado se presentó en las plántulas con la dosis más alta de yodo y con quemado en algunos bordes de las hojas.
El área foliar aumentó en las plántulas con aplicaciones de yodo en concentraciones medias (10 a 45 μM L-1) (Cuadro 1). La aplicación de 50 μM L-1 produjo plantas con hojas más pequeñas y algunos síntomas de toxicidad. La misma tendencia se reporta en la biofortificación de lechuga (Lactuca sativa) con siete concentraciones (10, 20, 30, 40, 80, 160, y 240 μM L-1 KI) y disminuy el crecimiento de la planta y producción de biomasa al aplicar más de 20 μM L-1 de KI y la reducción es más notoria con 40 μM L-1 KI. El efecto tóxico del yodo en alta concentración está descrito para varias especies (Caffaggni et al., 2011; Landini, et al., 2011; Zhu y Liu, 2003), pero no está bien entendido. Es probable que el exceso de yodo interfiera con el metabolismo y señalización celular de las ROS, causando respuestas negativas en el crecimiento y otros procesos.
La MS fue favorecida al aplicar a las hojas10 μM L-1 de yodo; las plántulas testigo presentaron el peso seco más bajo (Cuadro 1) y con la dosis intermedias los valores fueron más bajos. Jiang et al. (2001) reportan resultados similares así como un efecto fitotóxico sobre el crecimiento de las plantas, y puede deberse a una acumulación excesiva del yodo en los tejidos vegetales, o que ya absorbido puede darse una oxidación intracelular a I2 que inhibe el proceso fotosintético y se refleja en el desarrollo de la planta.
Índices de crecimiento
Andrews et al. (2001) y Raven et al. (2005) indican que los niveles de radiación, fotoperiodo, agua y nutrientes influyen en la acumulación de la MS y el área foliar, las cuales son factores determinantes en los índices de crecimiento. Y en las hojas se sintetizan los carbohidratos que se distribuyen a los órganos de la planta (Amaro et al., 2004).
La aplicación de yodo modificó la dinámica de la acumulación de MS, cambiando los valores de los índices de crecimiento (Cuadro 2). Los datos (Cuadro 1) señalan que el tratamiento de yodo 10 μM causó el mayor peso seco final, pero los datos en el Cuadro 2 indican que el comportamiento dinámico de la acumulación fue distinta en cada caso. Según estos resultados, la aplicación de yodo en diferentes concentraciones modificó la estrategia de reparto de la biomasa, su eficiencia de producción y como se utiliza para formar estructuras fotosintéticas.
Cuadro 2 Índices de crecimiento en plántulas de pimiento morrón con aplicación foliar de seis dosis de KI.
| Aplicación foliar | TAN g cm 2 d -1 | TCA g d -1 | TCR g g -1 d -1 | AFE Cm 2 g -1 |
|---|---|---|---|---|
| Testigo 0 μM | 0.00011 d | 0.0385 c | 0.0101 a | 32.037 b |
| Yodo 10 μM | 0.00055 d | 0.0378 c | 0.0471 a | 19.441 a |
| Yodo 15 μM | 0.0359 c | 0.0427 b | 0.0424 f | 19.111 c |
| Yodo 30 μM | 0.0419 b | 0.0603 a | 0.0485 d | 24.861 c |
| Yodo 45 μM | 0.0514 a | 0.0616 a | 0.0086 e | 25.178 c |
| Yodo 50 μM | 0.00081 d | 0.0182 d | 0.0073 c | 14.906 d |
| R2 | 0.9991 | 0.9843 | 0.9988 | 0.9731 |
| C. V. | 3.0605 | 4.5303 | 2.4295 | 3.7328 |
| DMS | 0.0009 | 0.0026 | 0.0009 | 1.0337 |
Medias con diferente letra son estadísticamente diferentes (p≤0.05); TAN: Tasa de asimilación neta; TCA: Tasa de crecimiento absoluto; TCR: Tasa de creci miento relativo; AFE: Área foliar específica; R2: Coeficiente de determinación; C.V.: Coeficiente de variación; DMS: Diferencia mínima significativa
Los valores de TAN en el Cuadro 2 indican para cada tratamiento el peso seco producido por unidad de área foliar. Los tratamientos de 10 y 15 μM produjeron menos biomasa por unidad de área foliar, pero tienen mayor área foliar (Cuadro 1) y acumularon más biomasa seca. Los tratamientos con yodo de 30, 45 y 50 μM fueron más efectivos que el testigo para acumular biomasa por unidad de área foliar y acumularon más MS que este último. Los cambios en TAN indican modificaciones morfológicas, fisiológicas y bioquímicas asociadas con el metabolismo del carbono (Shipley, 2002). En las plantas marinas el yodo ejerce un efecto significativo sobre el metabolismo fotosintético (Kupper et al., 2008), pero no hay información sobre plantas terrestres.
Los resultados de TAC y TCR (Cuadro 2) muestran un comportamiento complejo para las concentraciones de yodo. Los valores altos de TCA en los tratamientos de 30 y 45 μM, que no tuvieron la mayor biomasa seca final (Cuadro 1), muestran que la plántulas no crecieron, sino que tuvieron una combinación de períodos de lento y rápido crecimiento, lo que pudiera indicar conflictos entre aporte y demanda. No obstante, presentaron mayor biomasa que el testigo. La TCR fue mayor para los tratamientos de 10, 15 y 30 m M, esto es, la mayor eficiencia en la acumulación de biomasa fue con las concentraciones intermedias de yodo.
Los datos del AFE (Cuadro 2) señalan los cambios inducidos por la concentración de yodo en la estrategia de la planta para repartir la biomasa seca. El AFE es un indicador de cómo la planta acumula biomasa seca en las estructuras foliares. Los valores bajos del AFE en los tratamientos de 10 y 15 μM muestran que las plantas tuvieron una alta densidad de estructuras fotosinténticas, cuya evidencia es la gran cantidad de masa por unidad de área foliar, comparado con el testigo y con las aplicaciones de 30 y 45 μM KI. Potencialmente esta característica se asociaría con mayor eficiencia de absorción de radiación PAR por unidad de área foliar. (Yao et al., 2016).
Es difícil explicar con mayor amplitud los resultados anteriores sin tener más información bioquímica, anatómica y genómico. Pero, los resultados muestran una dirección para investigaciones acerca del efecto del yodo en las plantas de cultivo.
Actividad antioxidante total
Las aplicaciones de yodo, como yoduro de potasio, aumentan la capacidad antioxidante de los tejidos de las plantas. En la Figura 1 se muestra como todas las concentraciones de KI aplicadas aumentaron hasta 12 % la inhibición del DPPH, incluso con las dosis más altas de KI, las cuales redujeron la altura de las plantas algunos síntomas de toxicidad. Esta respuesta es similar a la observada en hojas de lechugas tratadas con dosis de KI menores a 40 μM (Leyva et al., 2011). En otro experimento con lechugas, aplicaciones de 10 a 40 μM causaron un claro incremento en la actividad antioxidante total, debido principalmente a un aumento en los niveles de ácido ascórbico en su forma reducida y esta molécula tiene una capacidad alta para donar electrones y neutralizar directamente radicales libres en reacciones enzimáticas y no enzimáticas (Blasco, 2010). La respuesta antioxidante obtenida con estos tratamientos es relevante porque eleva la tolerancia de las plantas ante condiciones de estrés (Stevens et al., 2008).
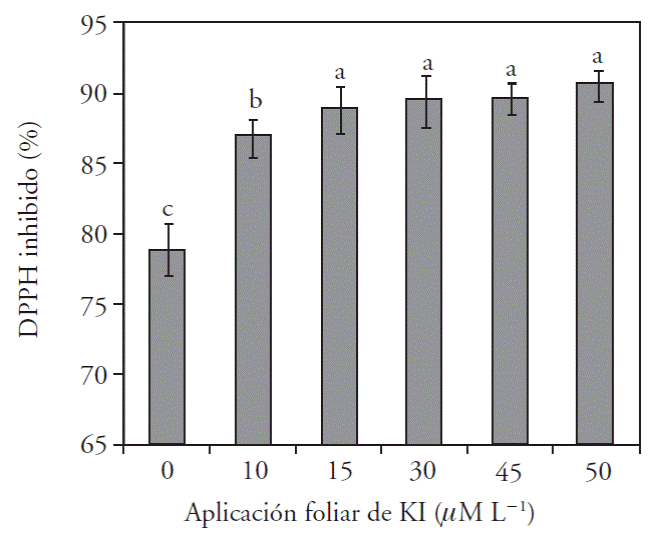
Contenido de yodo y minerales en la parte aérea de las plántulas
La concentración de yodo en las plántulas de pimiento morrón fue directamente proporcional a la dosis aplicada vía foliar (Figura 2). Hubo un aumento notable en la concentración de yodo en el tratamiento de 45 μM L-1 y de tres veces más con 50 μM L-1 respecto a los otros tratamientos.

Figura 2 Concentración de yodo en tejido vegetal de plántulas de pimiento morrón con aplicación foliar de seis dosis de KI.
Los resultados de nuestro estudio muestran como la absorción de yodo por las plántulas de pimiento morrón es directamente proporcional con las concentraciones aplicadas y hay una tendencia similar entre la concentración de yodo en parte aérea y antioxidantes totales. Estos resultados son similares a los reportados por Ujowondu et al. (2010) al biofortificar con yodo especies de hortalizas nativas de Nigeria, y hubo una mayor concentración de yodo en los tejidos vegetales al aumentar las dosis de yoduro y yodato de potasio en sus tratamientos.
Jiu et al. (2006) señalan que la concentración de yodo aumenta en las hojas y en la raíz en espinaca al aumentar las dosis de yoduro y yodato de potasio. Además, Zhu et al. (2003) reportan que la acumulación de yodo en tejido foliar de espinaca es directamente proporcional a las dosis de yoduro.
En la parte aérea, los resultados muestran que la aplicación de yodo incrementó en 5 % la concentración de nitrógeno, excepto con 45 μM L-1 la cual redujo el contenido de N, posiblemente por efecto de toxicidad (Figura 3).
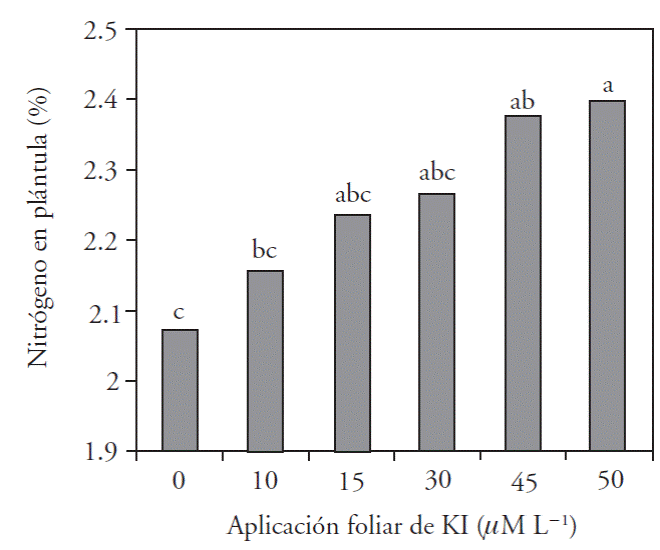
Figura 3 Concentración de nitrógeno total en tejido vegetal de plántulas de pimiento morrón con aplicación foliar de seis dosis de KI.
La concentración más baja de P, K, Ca, Mg, S se presentó en las plantas testigo, pero las dosis bajas y medias de yodo incrementan las concentraciones de los macro y minerales (Cuadro 3). Blasco et al. (2011) indican que la aplicación de yodo en la lechuga a dosis no mayores de 40 μM L-1, favorece la calidad nutricional, sin un efecto fitotóxico, por lo que consideran al yodo como un elemento beneficioso para las plantas y puede utilizarse en esas dosis. Jiu et al. (2004), al estudiar el efecto residual de yodo en el suelo, mencionan que concentraciones bajas de este elemento promueve la absorción y aprovechamiento de elementos esenciales en varias especies de plantas.
Cuadro 3 Concentración de minerales en parte aérea de plántulas de pimiento morrón con aplicación foliar de seis dosis de KI.
| Aplicación foliar | P | K | Ca mg kg -1 | Mg | S |
|---|---|---|---|---|---|
| Testigo 0 μM L-1 | 517 a | 3593 a | 5715 b | 5874 | 1860 |
| 10 μM L-1 | 1288 a | 4371 a | 12973 ab | 9648 | 2347 |
| 15 μM L-1 | 1163 a | 4599 a | 16599 a | 9525 | 2210 |
| 30 μM L-1 | 1134 a | 4072 a | 12630 | 8882 | 2101 |
| 45 μM L-1 | 990 a | 4473 a | 12184 | 8549 | 2085 |
| 50 μM L-1 | 846 a | 3902 a | 12030 | 8005 | 1897 |
| R2 | 0.4932 | 0.1426 | 0.6358 | 0.3890 | 0.6227 |
| C.V. | 20.053 | 11.6432 | 19.8691 | 13.870 | 15.541 |
| LSD | 484.14 | 1199.9 | 6507.20 | 2952.5 | 747.48 |
Los resultados (Cuadro 2) indican que una dosis óptima de aplicación foliar de yodo está entre 10 a 15 μM L-1, ya que favorece la absorción de los minerales. Además el incremento de área foliar por aplicación de yodo, muestra que el yodo favorece el desarrollo y absorción de minerales en plántulas de pimiento morrón.
Conclusiones
La aplicación foliar de yodo, como KI, en plántulas de pimiento morrón mejora la absorción de minerales, la cantidad y actividad de antioxidantes y el vigor de la plántula. Por lo tanto, la inclusión de yodo en la producción de almácigos permite obtener material vegetal de mejor calidad al trasplante.

