










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgrociencia
On-line version ISSN 2521-9766Print version ISSN 1405-3195
Agrociencia vol.49 n.5 Texcoco Jul./Aug. 2015
Recursos naturales renovables
Respuesta de árboles de duraznero [Prunus persica (L.) Batsch] compuesto de dos cultivares en un ambiente con incidencia de heladas: floración y amarre de fruto
Response of peach [Prunus persica (L.) Batsch] trees composed by two cultivars in an environment of frost incidence: bloom and fruit set
Horacio Santiago-Mejía1, José I. Cortés-Flores2*, Antonio Turrent-Fernández3, Manuel Livera-Muñoz4, Edmundo García-Moya5, Hilda A. Zavaleta-Mancera5, Víctor H. Volke-Haller2
1 Universidad Intercultural del Estado de México. División de Desarrollo Sustentable. 50640. Libramiento Francisco Villa S/N, Centro, San Felipe del Progreso, Estado de México.
2 Edafología. Campus Montecillo. Colegio de Postgraduados. 56230. Carretera México-Texcoco km. 36.5 Montecillo, Estado de México. *Autor responsable (jicortes@colpos.mx).
3 Campo Experimental Valle de México. INIFAP. 56250. Carretera los Reyes-Texcoco, Km.13.5. Coatlinchán, Texcoco, Estado de México.
4 Genética. Campus Montecillo. Colegio de Postgraduados. 56230. Carretera México-Texcoco km. 36.5 Montecillo, Estado de México.
5 Botánica. Campus Montecillo. Colegio de Postgraduados. 56230. Carretera México-Texcoco km. 36.5 Montecillo, Estado de México.
Recibido: noviembre, 2014.
Aprobado: mayo, 2015.
Resumen
En México, las heladas que coinciden con la floración y el amarre del fruto del duraznero [Prunus persica (L.) Batsch] afectan su productividad y el uso de cultivares precoces injertados sobre cultivares tardíos (composición bicultivar) puede atemperar el daño por heladas en flores y frutos a diferencia de árboles unicultivares precoces alternados con tardíos. Por ello se evaluaron dos cultivares precoces (CP-Precoz y Puebla) y dos tardíos (Supremo y Zacatecas) de duraznero a la composición cultivar del árbol y su fertilización (NPK y pollinaza) en la floración y amarre de fruto en ambiente con incidencia de heladas. El estudio se estableció en el Campo Experimental Valle de México en el 2005 y durante el invierno 2011-2012 fueron evaluados la floración y el amarre de fruto. La composición bicultivar, retrasó 8 días el periodo de floración de Supremo en comparación a la unicultivar y 12 días con la fertilización 90-30-90 g de N-P2O5-K2O por árbol por año. Supremo como interinjerto incrementó 15.3 % la apertura floral de los cultivares precoces, con un efecto mayor en CP-Precoz con 3 kg de pollinaza por árbol por año; en Puebla se retrasaron 16 y 22 días el inicio y el final de la floración, en CP-Precoz el final de la floración se retrasó 5 días. Con excepción de Zacatecas, los cultivares incrementaron y retrasaron la floración e incrementaron el amarre de fruto al ser injertados con dos cultivares, lo cual sugiere que esta práctica es útil para reducir el daño por heladas.
Palabras clave: Heladas, cultivar precoz, cultivar tardío, interinjerto, fertilización.
Abstract
In Mexico, the frosts matching with the blooming and fruit set of peach plants [Prunus persica (L.) Batsch] affect their productivity; nevertheless the use of early cultivars grafted on late ones (bi-cultivar composition) can reduce the frost damages on flowers and fruits, unlike early uni-cultivar trees alternated with late cultivars. Because of this, two early (CP-Precoz and Puebla) and two late (Supremo and Zacatecas) peach cultivars were evaluated regarding their tree cultivar composition and their fertilization (NPK and poultry manure) during the blooming and fruit set in an environments with recurrent frost. The study was carried out in the Campo Experimental Valle de México (Experimental Field in Mexico Valley) during 2005, and in the winters of 2011 and 2012 blooming and fruit set were evaluated again. When using the bi-cultivar tree blooming was delayed 8 d in Supremo, as compared with the uni-cultivar one. When using 90-30-90 g fertilization formula (N-P2O5-K2O) per tree per year blooming was delayed 12 d. Supremo as inter-stock increased 15.3 % anthesis of early varieties, with a higher effect in the CP-Precoz with 3 kg poultry manure per tree and per year. In Puebla the first and last bloom were delayed 16 to 22 d, and the end of the blooming period was further delayed 22 d. In CP-Precoz the last bloom was delayed 5 d. With the exception of the Zacatecas cultivar, all treatments increased and delayed blooming, and increased fruit-set when submitted to peaches of the bi-cultivar composition, suggesting that this practice is helpful in reducing frost damage.
Key words: Ground frost, early cultivar, late cultivar, inter-graft, fertilization.
INTRODUCCIÓN
En el centro de México las variedades de duraznero que se cultivan en el sistema agroforestal Milpa Intercalada con Árboles Frutales (MIAF), confieren rentabilidad económica a las unidades familiares de producción (Cortés et al., 2005), debido al valor en el mercado de los frutos frescos cuando la oferta es baja. La productividad del duraznero en este sistema se afecta principalmente por las heladas durante el invierno e inicio de la primavera, periodo en el cual ocurren la floración y el amarre de fruto y los dos procesos son sensibles a las temperaturas bajas. Los sistemas de calefacción o de riego, los ventiladores, las cubiertas y los métodos químicos para la protección contra las heladas son poco accesibles porque requieren de inversiones cuantiosas. Una alternativa para reducir los efectos de las heladas es el uso de cultivares tardíos cuya floración y amarre de fruto ocurre al inicio de la primavera. Los cultivares de durazno comerciales locales predominantes son precoces y semitardíos, florecen en el invierno y tienen el riesgo de dañarse por heladas durante la floración y el amarre de fruto, y en algunos años la pérdida de la cosecha es total.
La región subtropical del centro de México se caracteriza en el invierno por las noches frías y los días calurosos. Las temperaturas altas de los días invernales tienen un efecto negativo en la acumulación de unidades frío de la noche (Richardson et al., 1974; Miranda et al., 2013). En las noches existe pérdida de calor por radiación; se presentan inversiones térmicas a través del dosel del árbol, debido a que el aire frío más denso desciende a las zonas bajas y el aire cálido más ligero asciende (Barrales et al., 2002). En las heladas de radiación la temperatura del aire a una altura entre 1.25 a 2.0 m por encima del nivel del suelo, dentro de una garita meteorológica, es de 0 °C o menos, los estratos bajos del dosel pueden sufrir daños mayores por las temperaturas bajas (Lhomme et al., 2007).
La selección de cultivares y prácticas de manejo adecuadas pueden reducir el daño por heladas de radiación e incrementar la producción en regiones de producción de duraznero templadas y subtropicales (Pérez, 2004). Los portainjertos e interinjertos en el duraznero confieren ventajas en adaptación, vigor, nutrición, producción y calidad del fruto a las variedades comerciales (Giorgi et al., 2005; Gullo et al., 2014; Malcolm et al., 2014). La nutrición adecuada del duraznero incrementa la tolerancia a condiciones restrictivas de crecimiento (Nario et al., 2003).
El objetivo de esta investigación fue evaluar la respuesta de árboles de durazno compuestos de dos cultivares precoces y dos tardíos, fertilizados con NPK y pollinaza, y cultivados en un ambiente con heladas, en la floración y amarre de frutos. La hipótesis fue que durazneros compuestos con un cultivar precoz en el estrato medio superior, injertado en uno tardío, atempera más el efecto negativo de las heladas de radiación sobre la floración y el amarre de fruto, y este efecto es más notable que en durazneros precoces alternados con tardíos.
MATERIALES Y MÉTODOS
Material vegetal
Los portainjertos en todos los tratamientos fueron plantas germinadas de semilla criolla de durazno (Cortés-Flores[6]). Los cultivares evaluados fueron: CP Precoz y Puebla, de floración temprana y Supremo y Zacatecas de floración tardía. CP Precoz y Supremo se obtuvieron del programa de Fruticultura del Colegio de Postgraduados; Puebla y Zacatecas fueron obtenidos por los productores de los estados de Puebla y Zacatecas, respectivamente.
Sitio experimental
El experimento se estableció en el Campo Experimental Valle de México (CEVAMEX), Texcoco, Estado de México, en el año 2005. El sitio está localizado a 19° 29' 17" N y 98° 53' 41" O, a una altitud de 2280 m. El clima del sitio es templado subhúmedo, con temperatura media anual de 15.45 °C y precipitación media anual de 644 mm. El suelo es Fluvisol móllico (Govaerts et al., 2008).
Diseño experimental
Un experimento factorial 25 se estableció con una repetición, en un diseño de parcelas divididas. Los dos niveles de cada factor fueron: 1) composición cultivar del árbol (Cva), bicultivar y unicultivar, 2) cultivar tardío (Vt), Supremo y Zacatecas, 3) cultivar precoz (Vp), CP-Precoz y Puebla, 4) fertilizante N-P2O5-K2O (F), 60-15-60 y 90-30-90 g por árbol por año, y 5) pollinaza (P), cero y 3 kg por árbol por año. El efecto factorial Cva y la interacción VtVpFP se usaron como contrastes de definición (CD) para definir cuatro lotes (bloques) de ocho unidades. Así, la lista de 32 tratamientos quedó fraccionada en cuatro lotes de ocho tratamientos cada uno, de acuerdo con las combinaciones 0-0, 0-1, 1-0 y 1-1 de los contrastes de definición (Cuadro 1). El método de lotificación se realizó de acuerdo con la teoría de los campos de Galois (Kempthome, 1979).
Manejo del cultivo
Las plántulas de los portainjertos de dos meses de edad fueron trasplantadas en mayo de 2005 y fertilizadas con la fórmula 30-30-30 g de N-P2O5-K2O y pollinaza 1 kg por árbol. La evaluación del injerto en los cultivares en los cultivares temprano y tardío se realizó en abril de 2006.
Los durazneros unicultivares tardíos, Supremo o Zacatecas, se alternaron sobre la misma hilera con un cultivar precoz, CP Precoz o Puebla (Figura 1A). Los durazneros bicultivares fueron árboles compuestos: estrato medio superior de un cultivar precoz injertado sobre un cultivar tardío (estrato medio inferior) (Figura 1B). Para ambas composiciones del árbol, el injerto sobre el portainjerto criollo se ubicó a los 15 cm del nivel del suelo. En los árboles bicultivares, el injerto de cultivar precoz se estableció a la altura de 1.5 m del árbol, en marzo de 2007.
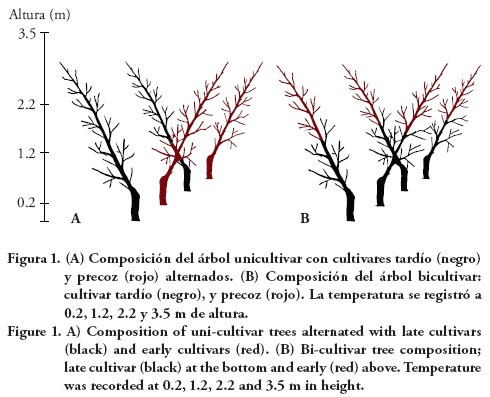
Los durazneros se plantaron en hileras con un espacio de 1 m entre ellas, con una orientación norte-sur. La conducción de los árboles fue con el sistema 'Tatura' modificado (Cortés et al., 2005), se dejó una rama principal con orientación perpendicular a la hilera e inclinación aproximada de 30° en relación al tronco del árbol. La rama del árbol contiguo se orientó en sentido opuesto al anterior y así sucesivamente (Figura 1). La altura promedio de los árboles a los 6 años de edad fue de 3 m. La unidad experimental fue de cuatro árboles y la unidad de observación incluyó los dos árboles centrales.
El fertilizante NPK y la pollinaza se aplicaron a partir del año 2006. Las fórmulas del fertilizante sintético fueron 20-520 o 30-10-30 g de N-P2O5-K2O por árbol, con incremento progresivo durante tres años, y desde el 2008 las fórmulas fueron 60-15-60 o 90-30-90 según el tratamiento. Las fuentes fueron urea, superfosfato triple y cloruro de potasio. Las dosis iniciales de pollinaza fueron cero y 1 kg por árbol con incremento progresivo hasta llegar a cero y 3 kg por árbol en el 2008. Las fechas de aplicación fueron abril (1/3; 1-1-0 de N, P, K, pollinaza) y agosto (2/3; 0-0-1 N, P, K, pollinaza) de cada año. El fertilizante de NPK y la pollinaza se aplicaron en las dos zonas de goteo de la hilera de árboles, a 20 cm de profundidad. Los durazneros fueron irrigados por surco sin restricción de humedad.
Con una microestación meteorológica WatchDog 1000 Series (Caectrum Technologies, Inc.; USA) instalada en el centro de la parcela experimental, equipada con termopares externos, se midió cada 15 min la temperatura del aire a 0.20, 1.20, 2.20 y 3.5 m de altura (Figura 1), del 17 de noviembre de 2011 al 31 de marzo de 2012. Con esta información se calcularon las UF de acuerdo con Vega et al. (1990) (Cuadro 2).

Variables evaluadas
Floración y amarre de fruto
Los análisis de floración y amarre de frutos del experimento se realizaron en tres secciones: 1) los efectos del estrato, cultivar tardío, cultivar precoz, fertilizante NPK y pollinaza fueron evaluados en los árboles unicultivares; 2) los efectos del interinjerto en el cultivar precoz, injerto en el cultivar tardío, fertilizante NPK y pollinaza, fueron evaluados en los árboles bicultivares; y 3) los efectos del cultivar tardío o precoz, fertilizante NPK y pollinaza, fueron evaluados en conjunto. Las variables respuesta en los árboles unicultivares se analizaron en el estrato medio inferior <1.5 m y medio superior >1.5 m, y en los bicultivares se analizaron el cultivar tardío y el precoz. Se evaluó: densidad de yemas (yemas m-1), apertura floral (%), densidad floral (flores m-1), inicio de la floración (días julianos al 10 % del total de flores abiertas), floración plena (días julianos al 80 % del total de flores abiertas), final de la floración (días julianos al 100 % de flores abiertas) y amarre de fruto (%). Para ello se marcaron al azar cinco ramillas mixtas en cada estrato (10 por árbol), de todos los tratamientos, a principios de noviembre del 2011 y se determinó el número de yemas florales, la longitud, las flores abiertas (cada cuatro días) y el número de frutos amarrados. El porcentaje de apertura floral fue calculado a partir del 100 % de yemas florales, y el de amarre de fruto a partir del 100 % de flores abiertas. Los días julianos fueron desde el 1 de enero de 2011 al 31 de marzo de 2012.
Análisis estadístico
Los datos de las variables de floración y amarre de fruto se analizaron mediante ANDEVA (p≤0.05 y p≤0.01) y análisis de regresión mediante el procedimiento de selección a pasos (Stepwise) (sle= 0.10 sls= 0.05) y variables mudas o dummy. En el estudio se hicieron ANDEVA del factorial 25 con una repetición y los grados de libertad del error se obtuvieron de las interacciones de orden mayor, asumiendo que estas son de menor importancia agronómica (Cuadro 3).

RESULTADOS Y DISCUSIÓN
La acumulación unidades frío (UF) a lo largo del estarto de los durazneros y las variables respuesta de floración y amarre de fruto se presentan en tres secciones: 1) en durazneros unicultivares, 2) en durazneros bicultivares, y 3) en el conjunto, esto es composición del cultivar tardío, cultivar precoz, fertilizante NPK y pollinaza. En el ANDEVA las fuentes de variación que tuvieron significancia en las variables respuesta corresponden a las incluidas en las ecuaciones de regresión que se reportan.
Unidades frío
A mayor altura en el perfil de árbol, la acumulación de frío fue mayor al final del invierno; 859, 1021, 1035, 1153 UF a 0.2, 1.2, 2.2 y 3.5 m de altura. La diferencia entre 0.2 y 3.5 m fue 294 UF. La temperatura del aire durante el día se incrementó con la disminución de la altura del árbol; la temperatura mayor se detectó a 0.2 m y la menor a 3.5 m. Las temperaturas altas durante el día tuvieron efecto negativo en la acumulación de frío; por esto los estratos más altos acumularon más UF que los bajos. Aunque hay evidencias del efecto negativo de las temperaturas altas en la acumulación de UF (Richardson et al., 1974; Miranda et al., 2013; Luedeling et al., 2013), no hay información suficiente de la acumulación de UF a través del perfil de los árboles frutales.
Las heladas más intensas se presentaron en el periodo en que aún no iniciaba la floración de las variedades precoces, por lo que no provocaron daños a las flores. Las yemas florales en letargo de los árboles aún no presentaban la conexión vascular entre el pedúnculo y el primordio floral (Santiago-Mejía, 2014), esta característica les confiere la capacidad de sobreenfriamiento y resistencia a las heladas (Ashworth, 1984).
Floración y amarre de fruto de durazneros unicultivares
Densidad de yemas
El número de yemas por metro de ramilla mixta de los durazneros unicultivares tardíos dependió de la interacción cultivar y estrato del árbol, mientras que en los precoces dependió del cultivar, del estrato del árbol y de la interacción fertilizante NPK y pollinaza. La densidad de yemas de Supremo y Zacatecas fue menor en el estrato medio inferior del árbol y se incrementó en el superior (Cuadro 4), con un efecto mayor en Supremo (Figura 2A). Los cultivares precoces, CP-Precoz y Puebla, también presentaron densidad mayor de yemas en el estrato medio superior que en el inferior, y Puebla presentó una densidad mayor de yemas que CP-Precoz (Cuadro 5). En ambos cultivares precoces, la fórmula baja de NPK (60-15-60 kg por árbol por año) y sin pollinaza aumentó la densidad de yemas comparado con la fórmula 90-30-90 kg por árbol por año; pero cuando se aplicaron 3 kg de pollinaza por árbol por año y la fórmula baja de NPK, la densidad de yemas se abatió y fue mejorada con la fórmula alta de NPK (Figura 2B). Esto podría indicar que con la dosis baja de NPK y sin pollinaza se obtendrían densidades de yemas similares que con la dosis alta de NPK y pollinaza.
El estrato medio superior al recibir mayor radiación fotosintéticamente activa y mejor calidad de luz incrementa la síntesis de fotosintetizados (Gullo et al., 2014; Marsal et al., 2014), citocininas (Bernier, 2005) y proteínas (Zeevaart, 2008) que estimulan la diferenciación floral. En cambio las hojas que reciben menor radiación solar tienen mayor concentración de giberelinas, que inhiben la diferenciación floral (González-Rossia et al., 2007; Sharp et al., 2010); esto pudo haber ocurrido en el estrato medio inferior de los árboles de duraznero. Aunque, la densidad floral se supone que depende principalmente del genotipo (Sánchez-Pérez et al., 2014), valores de 50 a 120 yemas por metro se han reportado, y está relacionada directamente con la producción final de fruto (Pérez, 2004). El incremento por efecto del estrato (en promedio 2.59 % más en el estrato medio superior que el inferior) tiene implicaciones agronómicas importantes en regiones de cultivo con presencia de heladas, ya que se reduce la exposición a las temperaturas bajas.
Apertura floral
En los durazneros unicultivares tardíos, el porcentaje de apertura floral de la ramilla mixta dependió de las interacciones cultivar y cultivar precoz alternado, y cultivar precoz alternado y fertilizante NPK; mientras que en los unicultivares precoces dependió del cultivar tardío alternado y de la interacción cultivar por pollinaza (Cuadro 4). La apertura floral de Supremo fue mayor (21.28 %) al alternarse con Puebla que con CP-Precoz con la fórmula baja de NPK, 60-15-60 g de N-P2O5-K2O por árbol por año; pero, con la fórmula alta de NPK, 90-30-90 g de N-P2O5-K2O por árbol por año se abatió 13.5 %. Mientras, Zacatecas al alternarse con Puebla tuvo efecto depresivo mayor en la apertura floral que con CP-Precoz y con ambas fórmulas de fertilización, sobre todo con la baja.
La apertura floral de CP-Precoz se incrementó con la aplicación de 3 kg de pollinaza por árbol por año, tanto al alternarse con Supremo como con Zacatecas (18.5 y 9.7 %). En Puebla incrementó 4 % al alternarse con Zacatecas y con la aplicación de 3 kg de pollinaza por árbol por año; sin embargo, abatió 6.7 % su apertura cuando se alternó con Supremo y con la dosis alta de pollinaza. Algunos cultivares pueden ser más eficientes en la utilización de los nutrientes debido a su vigor (Zarrouk et al., 2005); como CP-Precoz, que es menos vigoroso que Puebla. Además, Puebla tendría mayor competencia por nutrientes con Supremo que con Zacatecas (Marsal et al., 2014).
Densidad floral
El número de yemas florales por metro de ramilla mixta en los durazneros unicultives tardíos dependió del estrato, la variedad y de la interacción entre cultivar y cultivar precoz alternado; en cambio en los durazneros unicultivares precoces dependió únicamente del cultivar y el cultivar precoz alternado (Cuadro 4). No obstante que Supremo tuvo 16.5 flores m-1 más que Zacatecas, la densidad floral fue mayor en el estrato medio superior que en el medio inferior (3.5 flores m-1) en ambos cultivares; y se incrementó en Supremo al alternarse con Puebla que con CP-Precoz (6.34 flores m-1), pero se abatió en Zacatecas (4.5 flores m-1) al alternarse con Puebla que con CP-Precoz. Puebla tuvo 7.9 flores m-1 más que CP-Precoz, y la densidad floral se incrementó 2.5 flores m-1 en promedio para ambos cultivares cuando se alternaron con Zacatecas que con Supremo (Cuadro 6).
Cada cultivar tiene respuesta específica cuando se alterna con otro y dependerá también de la fertilización, y esta forma de cultivo genera competencia por nutrientes, humedad y radiación entre los árboles que demerita la apertura y la densidad floral (Smith, 1991; Elloumi et al., 2014). Los cultivares tardíos que requieren cantidad mayor de frío que los precoces, respondieron al efecto del estrato del árbol. El gradiente ascendente de frío acumulado, con la altura del dosel de los durazneros, estuvo directamente relacionado con el número de flores por metro de rama. El suministro adecuado de frío en duraznero, favorece el proceso normal de morfogénesis de los meristemos florales en cambio, la deficiencia de frío les provoca malformaciones (Nava et al., 2009), como ocurrió en Zacatecas, que presentó granos de polen abortados y malformados y deficiencias en la apertura floral (Bonhomme et al., 2005; Okie and Blackburn, 2011). Cuando hay daños por heladas a las flores, el interés agronómico se enfoca en incrementar el porcentaje de apertura floral y la densidad de flores; los incrementos mostrados en apertura y densidad floral para cada cultivar pueden significar un incremento sustancial en la producción final de fruto.
Periodo de floración
En los durazneros unicultives tardíos, los días al inicio de la floración, la floración plena y el final de la floración dependieron principalmente del cultivar, y el final de la floración recibió influencia de la interacción cultivar y cultivar precoz alternado; en contraste, estas variables en los cultivares precoces dependieron del estrato del árbol y del cultivar mismo. Zacatecas fue más tardío (en promedio 22 días) en todo el periodo de floración que Supremo (Cuadro 4), pero el final de la floración se adelantó 10 días cuando se alternó con Puebla que con CP-Precoz; quizá por una condición de agobio debido a una competencia entre los cultivar (Smith, 1991; Reig et al., 2013). En cambio, Supremo tuvo un retraso ligero (3 días) en el final de la floración, cuando se alternó con Puebla que con CP-Precoz. A pesar que Puebla fue más precoz que CP-Precoz (en promedio 20 días) en el periodo de floración, el inicio y final de la floración se retrasaron en el estrato medio superior respecto al bajo (2.6 y 3.3 días, respectivamente) en ambos cultivar (Cuadro 5). Esta respuesta pudo deberse al fenotipo, al patrón acrópeto de la floración (Pérez, 2004) y al rasgo poligénico de heredabilidad alta en la fecha de floración (Sánchez-Pérez et al., 2014).
Amarre de fruto
El porcentaje de amarre de fruto en los durazneros unicultivares tardíos dependió del estrato del árbol, del cultivar mismo, del cultivar precoz alternado y de la interacción entre estos dos últimos (Cuadro 4); en cambió los durazneros unicultivares precoces solo dependieron del estrato y el cultivar (Cuadro 5). Supremo tuvo porcentaje mayor de amarre de fruto que Zacatecas (en promedio 9.5), aun así, en ambos cultivares, el amarre de fruto se incrementó 5.5 % en el estrato medio superior con relación al medio inferior; pero en este disminuyó el mismo porcentaje cuando los cultivar tardíos se alternaron con Puebla que con CP-Precoz (Figura 3). Puebla y CP-Precoz tuvieron amarre mayor de fruto en el estrato medio superior del árbol que el medio inferior (4.3 %), y se incrementó aún más cuando se alternaron con Zacatecas que con Supremo (2.8 %).

El incremento del amarre de fruto en el estrato medio superior estuvo estrechamente relacionado con la densidad floral mayor. El amarre de fruto y la densidad floral son favorecidos por la calidad e intensidad lumínica (Okie y Blackburn, 2011), aunque este fue en detrimento por la competencia de las variedades alternadas.
Floración y amarre de fruto de durazneros bicultivares
En los árboles bicultivares los cultivares tardíos, Supremo y Zacatecas, no fueron afectados por los cultivares precoces que se les injertó, lo cual pudo deberse a periodos diferentes de diferenciación floral, floración y amarre de fruto y que evitó competencia entre el cultivar tardío y precoz en el mismo árbol. En contraste, en los cultivares precoces, CP-Precoz y Puebla, la densidad de yemas florales y el periodo de floración fueron afectados por los cultivares en los que estaban injertadas (Cuadro 6).
Densidad de yemas
En el estrato medio superior de los cultivares precoces de los durazneros bicultivares el número de yemas por metro lineal de ramilla mixta dependió del cultivar y de la interacción entre cultivar tardío como interinjerto (Cuadro 6). Puebla en promedio tuvo 27.1 yemas m-1 más que CP-Precoz. Ambos cultivares, sin pollinaza e injertados en Supremo tuvieron una densidad mayor de yemas (13.4 yemas más m-1) que cuando se injertaron en Zacatecas; y con 3 kg de pollinaza por árbol por año, la densidad de yemas se incrementó de 49.4 a 55.9 yemas m-1 cuando fueron injertados en Zacatecas que en Supremo, pero sin alcanzar las 61.4 yemas m-1 cuando fueron injertados con Supremo y sin pollinaza. Supremo como interinjerto al parecer favoreció una mejor distribución de la biomasa (Gullo et al., 2014), evitando la competencia entre el crecimiento vegetativo y la diferenciación floral en los cultivares precoces (Chalmers et al., 1981; Aloni et al., 2010). Este efecto puede ser mejorado con una fertilización adecuada para cada cultivar (Nario et al., 2003).
Periodo de floración
En los cultivares precoces de los durazneros bicultivares, la fecha de floración fue afectada por el cultivar y por la interacción entre cultivar tardío como interinjerto y cultivar. A pesar que Puebla fue más precoz que CP-Precoz (Cuadro 6), atrasó el inicio y la plena floración cuando se injertó en Supremo que en Zacatecas (15.7 y 9.7 días, respectivamente); en cambio CP-Precoz los atrasó en menor grado 3.8 y 7.6 días, respectivamente, cuando se injertó en Zacatecas que en Supremo (Figuras 4 A y B). La interacción de los cultivares comerciales con los interinjertos o injertos puede modificar el periodo de floración (Carrera y Gomez-Aparasi, 1998; Tomaz et al., 2010; Ghrab et al., 2014). El retraso del periodo de la floración de Puebla significó evadir cinco heladas no severas al inicio de la floración y tres en floración plena; en el periodo de retrasó para CP-Precoz no ocurrieron heladas. Aunque las heladas que ocurrieron en este periodo crítico no causaron daños, el retraso de la floración puede significar que la producción de fruto se salve con la ocurrencia de heladas más severas.
Efecto de la composición varietal en la floración y amarre de fruto de cultivares tardíos
Floración
En los cultivares tardíos no hubo diferencias en la densidad de yemas, apertura floral, densidad floral, inicio y plena floración por efecto de la composición varietal del árbol; pero sí en el final de la floración y amarre de fruto (Cuadro 7). A pesar que Supremo es menos tardío que Zacatecas, el final de la floración se retrasó en promedio 8 días en los durazneros bicultivares que en los unicultivares, con un efecto mayor de 12 días con la fórmula alta 90-30-90 g de N-P2O5-K2O por árbol por año. La respuesta de Zacatecas fue variable, con la fórmula baja de N-P2O5-K2O, 60-15-60, el final de la floración se adelantó 4 días en los árboles bicultivares con relación a los unicultivares, y con la alta se retrasó 8 días, pero solo superó un día a la fórmula baja de NPK (Figura 5). Aunque durante el periodo de retraso del final de la floración en Supremo y Zacatecas no se presentó alguna helada, en retrasos similares puede significar el logro de la producción (Pendergrass et al., 2000).

Amarre de fruto
A pesar que Supremo tuvo 8.3 % más amarre de fruto que Zacatecas, ambos cultivares lo incrementaron 1.8 % en promedio en los durazneros tardíos bicultivares que los unicultivares (Cuadro 7).
Efecto de la composición cultivar en la floración y amarre de fruto de cultivares precoces
Floración
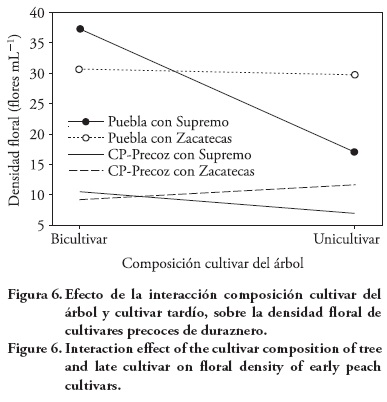
En las cultivares precoces no hubo diferencias en la densidad de yemas, pero sí en la apertura, densidad floral y periodo de floración por efecto de la composición cultivar del árbol (Cuadro 8). Con Supremo los cultivares precoces tuvieron en promedio 15.3 % más flores abiertas en los árboles bicultivares que en los unicultivares; mientras que con Zacatecas la apertura floral de los cultivares precoces fue 3.8 % mayor en los durazneros unicultivares. Además, CP-Precoz incrementó 10.9 % su apertura floral con 3 kg de pollinaza por árbol por año, pero en Puebla esta dosis de pollinaza tuvo efecto depresivo en la apertura floral, 3.8 % menos.
Puebla tuvo en promedio 9.6 flores m-1 más que CP-Precoz (Cuadro 8), injertado sobre Supremo (composición bicultivar) tuvo 20 flores m-1 más que en forma alternada (composición unicultivar); injertado sobre Zacatecas también tuvo 1 flor m-1 más que alternado, pero 6.6 flores m-1 menos que injertado en Supremo. CP-Precoz tuvo la mayor densidad floral (11.6 flores m-1) cuando se alternó a Zacatecas, injertado en Supremo la densidad floral fue mayor (3.6 flores m-1 más) que alternado a él (Figura 6).
En los durazneros bicultivares versus los unicultivares Puebla injertado sobre Supremo retrasó 16 días el inicio (Figura 7A) y 22 el final (Figura 7B) de la floración, e injertado en Zacatecas no modificó la fecha de inicio de floración, pero si retrasó 10 días el final de la floración, aunque 14 días menos que con Supremo. En cambio CP-Precoz alternado con Zacatecas el inicio de la floración fue atrasado 4.4 días en los árboles unicultivares respecto a los bicultivares (Figura 7A), pero esta respuesta se invirtió en la fecha del final de la floración, hubo un retraso de un día en los árboles bicultivares en relación a los unicultivares. El comportamiento del inicio y final de la floración fue similar cuando CP-Precoz se injertó en Supremo pero en menor grado (Figuras 7 A-B). En Puebla el retraso del inicio y final de la floración implicó la evasión de cinco y una heladas no severas, respectivamente.
Amarre de fruto
Puebla injertado sobre Supremo en la composición bicultivar duplicó el amarre de fruto que alternado en la unicultivar, 16.4 y 7.9 %; aunque fue en promedio 3.4 % mayor ya sea injertado o alternado con Zacatecas. CP-Precoz injertado sobre Supremo también incrementó 3.4 % el amarre de fruto que de forma alternada; sin embargo, cuando se injertó sobre Zacatecas se abatió de 14.2 a 0.8 %. Una combinación correcta de interinjerto y el cultivar induce a un equilibrio fisiológico que incrementa la producción de frutos debido a la distribución mejor de nutrientes y fotosintatos (Stevens y Weswood, 1984; Marra et al. 2013; Gullo et al., 2014); pero en algunos casos el portainjerto puede reducir la producción de frutos (Scarpare et al. 2000; Giorgi et al., 2005).
CONCLUSIONES
Durante el período de floración, cultivares precoces y tardíos de duraznero se sometieron a la composición cultivar del árbol y a la fertilización con NPK y pollinaza, y hubo un gradiente ascendente de acumulación de unidades frío del estrato medio inferior al medio superior del árbol, con apertura y densidad floral mayores en el estrato medio superior del árbol.
La composición bicultivar del duraznero comparado con el unicultivar, retrasó el final de la floración de Supremo, con un efecto mayor con la fórmula alta de NPK, e incrementó el amarre de fruto de Supremo y Zacatecas. Además, en los cultivares precoces injertados en Supremo, la apertura floral se incrementó con un efecto mayor en CP-Precoz al aplicar pollinaza; en Puebla retrasó el inicio de la floración; en Puebla y CP-Precoz retrasó el final de la floración, e incrementó el amarre de fruto en Puebla y CP-Precoz.
En durazneros compuestos de un cultivar precoz en el estrato medio superior injertado sobre un cultivar tardío (composición bicultivar) con una fertilización adecuada de NPK y pollinaza, se retrasaró la floración e incrementó el amarre de fruto, en comparación con los durazneros precoces (composición unicultivar) alternados con tardíos.
Aunque las heladas a las que se expusieron los durazneros no fueron severas, la composición bicultivar se muestra como una estrategia prometedora para evitar o disminuir los daños por heladas de radiación.
LITERATURA CITADA
Aloni, B., R. Cohen, L. Karni, H. Aktas, and M. Edelstein. 2010. Hormonal signaling in rootstock—scion interactions. Sci. Hort. 127: 119-126. [ Links ]
Ashworth, E. N. 1984. Xylem development in Prunus flower buds and the relationship to deep supercoiling. Plant Physiol. 74: 862-865. [ Links ]
Barrales, D. J. S., M. Livera M., V. González H., C. B. Peña V., J. Kohashi S. y F. Castillo G. 2002. Relaciones térmicas en el sistema suelo-planta-atmósfera durante la incidencia del fenómeno de enfriamiento o helada. Rev. Fitotec. Mex. 25: 289-297. [ Links ]
Bernier, G. 2005. The florigen quest: classical developments. Flower Newslett 40: 4-11. [ Links ]
Bonhomme, M., R. Rageau, A. Lacointe and M. Gendraud. 2005. Influences of cold deprivation during dormancy on carbohydrate contents of vegetative and floral promordia and nearby structures of peach buds (Prunus persica L. Batch). Sci. Hort. 105: 223-240. [ Links ]
Carrera, M. and J. Gomez-Aparasi. 1998. Rootstock influence on the performance of the peach variety 'Catherine'. Acta Hortic. 465: 573-578. [ Links ]
Chalmers, D. J., P. D. Mitchell, and L. Van Heek. 1981. Control of peach tree growth and productivity by regulated water supply, tree density, and summer pruning. J. Am. Soc. Hort. Sci. 106: 307-312. [ Links ]
Cortés, J. I. F., A. Turrent F., P. Díaz V., R. Hernández E., R. Mendoza R., y E. Aceves. 2005. Manual para el establecimiento y manejo del sistema milpa intercalada con árboles frutales (MIAF) en laderas. SAGARPA-Colegio de Postgraduados. pp: 1-11. [ Links ]
Elloumi, O., M. Ghrab, and M. B. Mimoun. 2014. Effects of flower buds removal on seasonal starch storage and mobilization in fruiting and non-fruiting branches of pistachio trees cv. Mateur under dry and warm climate. Sci. Hort. 172: 19-25. [ Links ]
Giorgi, M., F. Capocasa, J. Scalzo, G. Murri, M. Battino, and B. Mezzetti. 2005. The rootstock effects on plant adaptability, production, fruit quality, and nutrition in the peach (cv. 'Suncrest'). Sci. Hort. 107: 36-42. [ Links ]
González-Rossia, D., C. Reig, M. Juan, and M. Agustí. 2007. Horticultural factors regulating effectiveness of GA3 inhibiting flowering in peaches and nectarines (Prunus persica L. Batsch). Sci. Hort. 111: 352-357. [ Links ]
Govaerts, B., M. G. Barrera-Franco, A. Limón-Ortega, P. Muñoz-Jiménez, D.K. Sayre, y J. Deckers. 2008. Clasificación y evaluación edafológica de tres sitios experimentales del altiplano central de México. Tropicultura 26: 2-9. [ Links ]
Ghrab, M., M. Ben M., M. Moncef M., and N. Ben M. 2014. Chilling trends in a warm production area and their impact on flowering and fruiting of peach trees. Sci. Hort. 178: 87-94. [ Links ]
Gullo, G., A. Motisi, R. Zappia, A. Dattola, J. Diamanti, and B. Mezzetti. 2014. Rootstock and fruit canopy position affect peach [Prunus persica (L.) Batsch] (cv. Rich May) plant productivity and fruit sensorial and nutritional quality. Food Chem. 153: 234-242. [ Links ]
Kempthome, O. 1979. The design and analysis of experiments. John Wiley & Sons. New York , N. Y., U.S.A., 631 p. [ Links ]
Lhomme, J.P., J.J. Vacher, and A. Rocheteau. 2007. Estimating downward long-wave radiation on the Andean Altiplano. Agricult. Forest Meterol. 145: 139-148. [ Links ]
Luedeling, E., L. Guo, J. Dai, C. Leslie, and M. M. Blanke. 2013. Differential responses of trees to temperature variation during the chilling and forcing phases. Agricult. Forest Meterol. 181: 33-42. [ Links ]
Malcolm, P. J., P. Holford, I. Barchia, and W. M. McGlasson. 2014. High and low root zone temperatures at bud-break reduce growth and influence dry matter partitioning in peach rootstocks. Sci. Hort. 171: 83-90. [ Links ]
Marra, F. P., R. Lo Bianco, M. La Mantia, and T. Caruso. 2013. Growth, yield and fruit quality of 'Tropic Snow' peach on size-controlling rootstocks under dry Mediterranean climates. Sci. Hort. 160: 274-282. [ Links ]
Marsal, J., S. Johnson, J. Casadesus, G. Lopez, J. Girona, and C. Stöckle. 2014. Fraction of canopy intercepted radiation relates differently with crop coefficient depending on the season and the fruit tree species. Agricult. Forest Meterol. 184: 1-11. [ Links ]
Miranda, C., L. G. Santesteban, and J. B. Royo, 2013. Evaluation and fitting of models for determining peach phenological stages at a regional scale. Agricult. Forest Meterol. 178-179: 129-139. [ Links ]
Nava, G., G. Dalmago, H. Bergamaschi, R. Paniz, R. dos Santos, and G. Marodin. 2009. Effect of high temperatures in preblooming and blooming periods on ovulo formation, pollen grains and yield of 'Granada' peach. Sci. Hort. 122: 37-44. [ Links ]
Nario, A., I. Pino, F. Zapata, M. P. Albornoz, and P. Baherle. 2003. Nitrogen (15N) fertiliser use efficiency in peach (Prunus persica L.) cv. Goldencrest trees in Chile. Sci. Hort. 97: 279-287. [ Links ]
Okie, W. R., and B. Bryan. 2011. Interactive effects of light and chilling on peach flower and leaf budbreak. HortScience 46: 1056-1062. [ Links ]
Pendergrass, R., R. K. Roberts, D. E. Deyton, and C. E. Sams. 2000. Economics of using soybean oil to reduce peach freeze damage and thin fruit. HortTechnology 10: 211-217. [ Links ]
Pérez, S. 2004. Yield stability of peach germplasm differing in dormancy and blooming season in the Mexican subtropics. Sci. Hort. 100: 15-21. [ Links ]
Reig, G., S. Alegre, F. Gatius, and I. Iglesias. 2013. Agronomical performance under Mediterranean climatic conditions among peach [Prunuspersica L. (Batsch)] cultivars originated from different breeding programmes. Sci. Hort. 150: 267-277. [ Links ]
Richardson, E. A., S. D. Seeley, and D.R. Walker. 1974. A model for estimating the completion of rest for 'Redhaven' and 'Elberta' peach trees. HortScience 9: 331-332. [ Links ]
Santiago-Mejía H. 2014. El sistema agroforestal Milpa Intercalada con Árboles Frutales (MIAF): composición varietal del árbol y fertilización de durazno en condiciones de heladas. Tesis doctoral. Colegio de Postgraduados, Montecillo, México. 75 p. [ Links ]
Sánchez-Pérez, R., J. del Cueto, F. Dicenta and P. Martínez-Gómez. 2014. Recent advancements to study flowering time in almond and other Prunus species. Front Plant Sci. Lausanne, Switzerland. 334 p.
Scarpare, F. J. A., R. A. Kluge, F. V. Filho, J. T.Neto, e A. P. Jacomino. 2000. Comportamento de duas cultivares de pesseguerio com interenxerto da maeixeira 'Junuária'. Pesq. Agropec. Bras. 35: 757-765. [ Links ]
Sharp, R. G., M. A. Else, W. J. Davies, and R. W. Cameron. 2010. Gibberellin-mediated suppression of floral initiation in the long-day plant Rhododendron cv. Hatsugiri. Sci. Hort. 124: 231-238. [ Links ]
Smith, M.W. 1991. Influence of tree spacing on performance of 'Garnet Beauty' peach. Sci. Hort. 48: 52-60. [ Links ]
Stevens, G., and M. N. Weswood. 1984. Fruit set and cytokinin-like activity in the xylem sap of sweet cherry (Prunus avium) as affected by rootstock. Physiol. Plant. 61: 464-468. [ Links ]
Tomaz, Z. F. P., C. S. Madruga L., M. Aldrighi G., L. Rufato, and A. De Rossi R. 2010. Crescimento vegetativo, floração e frutificação efetiva do pessegueiro 'Jubileu' submetido a diferentes comprimentos de interenxertos. Pesq. Agropec. Bras. 45: 973-979. [ Links ]
Vega, N. R., L. A. Aceves N., J. Trujillo A., y R. Arriaga R. 1990. Generación y aplicación de modelos agroclimáticos a la fenología de la palomilla de la manzana Cydia pomonella L. (Lepidoptera: Tortricidae), en Canatlán, Durango. Agrociencia 24: 89-100. [ Links ]
Zarrouk, O., Y. Gogorcena, J. Gómez-Aparisi, J.A. Betrán, and M.A. Moreno. 2005. Influence of almondXpeach hybrids rootstocks on flower and leaf mineral concentration, yield and vigour of two peach cultivars. Sci. Hort. 106: 502-514. [ Links ]
Zeevaart, J. AD. 2008. Leaf-produced floral signals. Curr. Opin. Plant Biol. 11:541-547. [ Links ]
6 José Isabel Cortés Flores, profesor investigador titular del Programa en Edafología del Colegio de Postgraduados, Campus Montecillo (correo electrónico: jicortes@colpos.mx).

