










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.49 no.2 Texcoco feb./mar. 2015
Recursos naturales renovables
Calidad del agua en la producción de Pinus cembroides Zucc. en vivero
Water quality for the forest nursery production of Pinus cembroides Zucc.
Jesús V. Gutiérrez-García, Dante A. Rodríguez-Trejo*, Antonio Villanueva-Morales, Silvia García-Díaz, José L. Romo-Lozano
División de Ciencias Forestales, Universidad Autónoma Chapingo. Chapingo, Estado de México, 56230. (valenton900@hotmail.com.mx) *Autor responsable (dantearturo@yahoo.com) (antoniov28@gmail.com) (edith65@gmail.com) (jlromo@aya.yale.edu).
Recibido: abril, 2014.
Aprobado: febrero, 2015.
Resumen
La calidad del agua es un factor relevante para la producción de especies forestales, pero no siempre se toma en cuenta en viveros de México. Uno de los parámetros de calidad de agua es su acidez. El objetivo de esta investigación fue estudiar la respuesta en la morfología y en la concentración de iones y cationes en el tejido foliar de Pinus cembroides Zucc., a diferentes niveles de pH de agua y concentraciones de compuestos en las soluciones de ferti-irrigación. Las plantas fueron cultivadas en vivero, en contenedores de 170 cm3, con sustrato a base de turba de musgo (40 %), vermiculita (30 %) y agrolita (30 %). El diseño experimental fue en bloques completamente al azar, con dos factores y dos niveles por factor: 1) pH del agua de riego (8 y 5.5); 2) fertilización (ppm): establecimiento (50-123-73 de N, P K para ambos niveles), crecimiento (100-22-83 y 150-33-125), y Analizador (15-41-109 y 30-82-218); estos niveles de fertilización se denominaron bajo y alto. El efecto de factores y su interacción fue determinado con un ANDEVA (p≤0.10). Las variables morfológicas e índices de calidad evaluados fueron: altura, diámetro, peso seco aéreo, peso seco de raíz, índice de calidad de Dickson (ICD), relación parte aérea/raíz, índice de lignificación, índice de esbeltez y concentración nutrimental foliar. No hubo respuesta en variables morfológicas ni en índices, pero sí en concentración nutrimental foliar. La mayor concentración foliar de K (p≤0.001) fue con la fertilización alta en ambos niveles de pH, y la de Mn con fertilización alta y sin ajuste de pH (8.0). Las mayores concentraciones de Ca (p≤0.092), Mg (p≤0.074) y Cu (p≤0.093) ocurrieron con fertilización alta y pH = 5.5, y la de Zn (p≤0.001) fue con fertilización baja y pH ajustado. Hubo interacciones entre fertilización y pH del agua para K (p≤0.005), Ca (p≤0.1) y Zn (p≤0.039). Las mayores concentraciones nutrimentales fueron para agua con pH = 5.5 y la fertilización alta.
Palabras clave: Agua, análisis nutrimental, calidad de planta, fertilización, pH del agua, Pinus cembroides.
Abstract
Water quality is a relevant factor for production of forest species, but it is not always taken into account in nurseries in Mexico. One of the parameters of water quality is its acidity. The objective of this work was to study the response in morphology and concentration of ions and cations in leaf tissue of Pinus cembroides Zucc., to different water pH and concentrations of compounds in fertigation solutions. Plants were cultivated in a nursery in 170 cm3 containers with a substrate made of peat moss (40 %), vermiculite (30 %) and agrolite (30 %). The experimental design was completely randomized blocks with two factors and two levels: 1) pH of irrigation water (8 and 5.5); 2) fertilization (ppm): establishment (50-123-73 of N, P, K for both levels), growth (100-22-83 and 150-33-125) and finalization (1541-109 and 30-82-218); these levels of fertilization were denominated low and high. The effect of factors and their interaction was determined with an ANOVA (p≤0.10). The morphological variables and quality indexes evaluated were height, diameter, shoot dry weight, root dry weight, Dickson quality index, shoot/root ratio, lignification index, slenderness index and leaf nutrient concentration. There was no response in terms of morphological variables or indexes. There was, however, a response in leaf nutrient concentration. The highest leaf concentration of K (p≤0.001) was obtained with high fertilization at both levels of pH, and the highest concentration of Mn with higher fertilization without adjusting pH (8.0). The highest concentrations of Ca (p≤0.092), Mg (p≤0.074), and Cu (p≤0.093) occurred with high fertilization and pH= 5.5, and that of Zn (p≤0.001) was with low fertilization and adjusted pH. There were interactions between fertilization and water pH for K (p≤0.005), Ca (p≤0.1) and Zn (p≤0.039). The highest nutrient concentrations were obtained with water pH= 5.5 and high fertilization.
Key words: Water, nutrient analysis, seedling quality, fertilization, water pH, Pinus cembroides.
INTRODUCCIÓN
Pinus cembroides Zucc. es el pino piñonero con más amplia distribución en 18 estados del norte y centro de México y en el sur de EE.UU. A pesar de su porte pequeño, proporciona leña y madera de cortas dimensiones para construcciones rurales. Su semilla es alimenticia y tiene alta demanda y valor comercial (Perry, 1991, Farjon et al., 1997). Por su relevancia ecológica se usa cada vez más en reforestaciones, en proyectos de restauración, debido a su resistencia a condiciones adversas de precipitación, suelo y heladas (Vázquez et al., 1999). Al fertilizar en el vivero forestal se debe cuidar que ningún nutrimento sea deficitario, pues la ley del mínimo de Liebig establece que el nutrimento en menor concentración limitará la producción. Pero al aumentar de manera gradual la fertilización, los incrementos en la producción se reducirán hasta no tener más respuesta en el cultivo, según la ley de los rendimientos decrecientes de Mitscherlich (García et al, 2009).
El agua es un factor primordial para el desarrollo de las plantas y en condiciones naturales es su principal factor limitativo. Casi todos los procesos fisiológicos de los vegetales están afectados en mayor o menor medida por el agua (Peñuelas y Ocaña, 1994). Un aspecto importante en el establecimiento y manejo de viveros forestales es la calidad del agua. Los fertilizantes y plaguicidas químicos contribuyen a aumentar el crecimiento de las plantas en los viveros, pero pueden contaminar las aguas superficiales y subterráneas cuando se liberan del medio de propagación (Landis et al., 1994).
En los viveros forestales, el agua de calidad tiene baja salinidad (salinidad total e iones tóxicos) y libre de hongos fitopatógenos, semillas de malezas, algas, y plaguicidas (Landis, 1989). Respecto a salinidad, los criterios para determinar la calidad del agua son: 1) salinidad, para evaluar el riesgo de que el agua ocasione altas concentraciones de sales, con el correspondiente efecto osmótico y disminución de rendimiento de cultivos; 2) sodicidad, para analizar el riesgo de inducir un elevado porcentaje de sodio intercambiable (PSI), con deterioro de la estructura del suelo; 3) toxicidad, para estudiar los problemas creados por algunos iones (Mendoza, 2009). Para viveros forestales en contenedores, el agua de riego no debe tener contaminantes ni fitopatógenos, y su pH debe ser 5 a 5.5 porque se debe a una intensiva aplicación de sales fertilizantes y el pH afecta la disponibilidad de los nutrimentos. En suelos orgánicos, el pH con mayor disponibilidad es 5 a 5.5 (Landis et al., 1994; Lambers et al., 1998). De acuerdo con Peñuelas y Ocaña (1994) y Ruano (2003), las principales técnicas para mejorar la calidad del agua son acidificación, ósmosis inversa, desionización, suavizantes de agua, cloración y filtración. En viveros forestales en contenedores, la acidificación involucra el uso de pequeñas dosis de ácido, principalmente ácido fosfórico (H3PO4).
El objetivo del presente estudio fue evaluar el efecto de dos niveles de pH de agua de riego y de dos niveles de fertilización, en la morfología y concentración nutrimental foliar de P. cembroides Zucc. La hipótesis fue que un nivel elevado de pH reduce la disponibilidad y, por ende, la concentración de nutrimentos en el follaje de los árboles y afecta su morfología.
MATERIALES Y MÉTODOS
Etapa de vivero
Este estudio se realizó en el vivero forestal de la División de Ciencias Forestales (DICIFO) de la Universidad Autónoma Chapingo, Estado de México, ubicado a 19° 29' 46'' N y 18° 00' 51" O, a una altitud de 2250 m. El clima es Cw, templado-frío con lluvias en verano, la temperatura media anual es 15 °C, con extremas promedio de 24.2 °C y 6.0 °C (García, 1978).
La semilla fue recolectada de árboles en una población de la especie en Querétaro, y se sembró en contenedores de 54 cavidades, con capacidad de 170 cm3 por cavidad. El sustrato fue a base de turba de musgo (peat moss) (40 %), vermiculita (30 %) y agrolita (30 %), más Trichoderma harzianum Rifai, cepa T-22 KRL-AG2 (PHC® T-22® ) (0.1 g L-1) y hongos ectomicorrízicos Pisolithus tinctorius (Pers.) Coker et Couch (PHC® Ecto-Rhyza® ) (200 g m-3), aplicados a la siembra. Un total de 864 árboles fueron producidos.
Ajuste del pH del agua de riego
Para acidificar el agua a los niveles de pH requeridos para el experimento, se preparó 1 L de solución de ácido fosfórico (H3PO4) al 1 % vertiendo 988.25 mL de agua destilada en una probeta y se agregaron 11.75 mL del ácido. Luego se colocó 1 L de agua de riego muestreada en un vaso de precipitado, se realizó la primera lectura con el potenciómetro (modelo HI 991002, Hanna instruments), se agregó 1 mL de la solución con ácido al 1 % y se obtuvo la siguiente lectura con el potenciómetro. Este procedimiento se repitió hasta obtener el pH de agua deseado. Así, para 1 L de agua de riego obtenida en diciembre de 2011, se redujo el pH original de 8 hasta 5.5, para lo cual se aplicaron 21 mL de solución al 1 % de H3PO4.
Fertilización
Ésta se realizó con el fertilizante comercial soluble Peters Professional®: iniciador (8-45-14), desarrollo (2010-20) y finalizador (4-25-35), y las dosis de fertilización fueron denominadas alta y baja. La dosis alta tuvo más N, P y K, en particular N (50 % más), que la dosis baja durante la etapa de desarrollo, y también una mayor concentración de los mismos nutrientes, en especial K, durante la fase finalizadora (100 % más) (Cuadro 1). Una vez establecido el experimento, se realizó ferti-irrigación con regadera y tres aplicaciones a capacidad de campo por semana, con uno a dos riegos más por semana. El fertilizante para la etapa de establecimiento se aplicó ocho semanas, el de la fase de crecimiento rápido 24 semanas, y el finalizador seis semanas.
Análisis morfológicos
Después de 9.5 meses los análisis morfológicos de calidad de planta en los brinzales fueron: diámetro del cuello de la raíz (con vernier digital, Trupper), altura de plántula (con regla graduada), biomasa aérea, biomasa de la raíz (las muestras fueron secadas en horno a 70 °C hasta obtener peso constante, y pesadas con una báscula electrónica GT41DD, Ohaus), y biomasa total (suma de pesos secos de la parte aérea y de la raíz).
Los índices morfológicos de calidad fueron: esbeltez (IE), la relación peso seco parte aérea/peso seco raíz (A/R), Dickson (ICD) (Dickson et al., 1960; citados por Duryea, 1985), y lignificación (IL) (Prieto et al., 2004) y se usaron los modelos 1 a 4.

donde A: altura del brinzal (cm) y D: diámetro del cuello de la raíz (mm).

donde BA: biomasa aérea (g) y BR: biomasa radical (g).
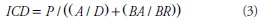
donde P: peso seco del brinzal (g).

donde Phum: peso del brinzal en húmedo (g).
Para el análisis nutrimental foliar se tomaron ocho muestras (plantas) por tratamiento, recolectadas al azar de cada unidad experimental, tomando toda la parte aérea de la planta y se separó el follaje. Las muestras foliares de cada árbol fueron mantenidas separadas, se secaron en la estufa del laboratorio de la DICIFO a 70 °C. En el Laboratorio Central de Suelos de la Universidad Autónoma Chapingo se realizó el análisis de: N (digestión con mezcla diácida y determinado por arrastre de vapor); P (digestión con mezcla diácida y determinado por fotocolimetría por reducción con molibdo-vanadato); K (digestión con mezcla diácida y determinado por espectrofotometría de emisión de flama); Ca, Mg, Cu, Zn, Mn (digestión con mezcla diácida y determinados por espectrofotometría de absorción atómica) y B (digestión con mezcla diácida y determinado por fotocolimetría con azometina-H).
Diseño experimental y modelo estadístico
El diseño experimental fue en bloques completos al azar, con dos factores: pH del agua de riego y fertilización. Cada factor tuvo dos niveles (pH de 5.5 y 8, y fertilización alta y baja; Cuadro 1), y hubo cuatro bloques. Cada unidad experimental consistió de 54 brinzales en arreglos de 6×9 cavidades y se asignaron aleatoriamente cuatro unidades experimentales por cada bloque.
El modelo estadístico de análisis fue (Infante y Zárate, 2000):
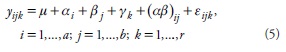
donde yijk: valor de respuesta del k-ésimo bloque, j-ésimo nivel del factor fertilización, e i-ésimo nivel del factor calidad de agua; μ: media general; αi: efecto del i-ésimo nivel del factor pH del agua; βj: efecto del j-ésimo nivel del factor nivel de fertilización; γk: efecto del k-ésimo bloque; (αβ)ij: efecto de la interacción entre el i-ésimo nivel del factor pH del agua, y el j-ésimo nivel del factor fertilización; εijk: error experimental.
Análisis estadístico
Con los datos se realizó un ANDEVA (SAS, 9.0), y las medias se compararon con la prueba de diferencia mínima significativa (p≤0.10). El análisis estadístico se efectuó para los tratamientos de fertilización usados en la etapa de finalización. También se usó SAS para cuatro pruebas de normalidad a los residuales del modelo estadístico ajustado a los datos de las variables e índices morfológicos evaluados: Shapiro-Wilk, Kolmogorov-Smirnov, Cramer-von Mises y Anderson-Darling (p≤0.05).
Los datos en el Cuadro 2 muestran que no hay evidencia sobre desviación de la hipótesis de normalidad de los datos en las variables morfológicas evaluadas. Los valores p de las cuatro pruebas son ≥0.15 para todas las variables, excepto peso seco parte raíz cuyos valores son de 0.0208 a 0.0662. En el Cuadro 3 todos los valores p son ≥ 0.15, esto es, no hay evidencia de una posible violación de la normalidad de los datos para los cuatro índices morfológicos evaluados.
Como complemento a los resultados obtenidos de las pruebas de normalidad se computaron los gráficos de probabilidad normal para los residuales de cada variable e índice morfológico evaluado (no mostrados). Los resultados en todos los casos muestran que no hay evidencia de que estos gráficos salgan del patrón de línea recta que se requiere para evidenciar normalidad.
Respecto a la posibilidad de varianza no constante (heterocedasticidad), los gráficos de residuales versus medias de tratamientos (resultados no mostrados), indican que no hay evidencia de que la dispersión (variabilidad) se desvíe de un patrón de varianza constante (homocedasticidad). En este caso, los tamaños de muestra fueron iguales para todos los tratamientos (experimento balanceado). Según la teoría, para experimentos balanceados el efecto sobre inferencia de una posible heterocedasticidad de los datos no es tan importante, sobre todo respecto a la prueba de F de igualdad de medias de tratamientos.
Finalmente, la independencia de las muestras quedó garantizada debido a la aleatorización de las unidades experimentales a los diferentes tratamientos y el orden aleatorio de la toma de observaciones.
RESULTADOS Y DISCUSIÓN
Las variaciones del pH del agua de riego, de diciembre de 2011 hasta noviembre de 2012, muestran una diferencia entre los valores máximo y mínimo (7.5 a 8.2) igual a 0.7 unidades, que representaron 9.3 % con respecto al valor mínimo y 8.5 % en relación al máximo. La media fue igual a 7.96. Durante la primavera los valores fueron elevados, pero aumentaron aún más durante la primera mitad del verano, para reducirse gradualmente en la segunda mitad. En el otoño e invierno hubo valores menores de pH, lo cual puede deberse a mayor precipitación en el verano y parte del otoño que aumentaría la infiltración de sales y, a su vez, aumentaría el pH alcalino del agua en el suelo.
Respecto a la evaluación de los indicadores e índices morfológicos, (Cuadros 4 y 5) no hubo diferencias significativas entre tratamientos para altura, diámetro, pesos secos de parte aérea y raíz, biomasa total, relación peso seco parte aérea entre peso seco raíz, índices de robustez, lignificación e índice de Dickson.
La relación peso seco parte aérea entre peso seco raíz, muestra un valor promedio de 1.17, lo cual es un buen balance entre parte aérea y subterránea de los brinzales (Duryea, 1985). Esta relación no debe exceder de 2.5, en especial cuando la planta se llevará a sitios con escasa precipitación (Thompson, 1985), como es el ambiente de la especie bajo estudio. El coeficiente de esbeltez promedio entre todos los tratamientos fue 4.08 (Cuadro 5), menor al valor límite máximo de 6 para que un brinzal esté equilibrado (Quiroz et al., 2009).
Las concentraciones foliares de nutrimentos en la especie estudiada fueron diferentes entre tratamientos (Cuadro 6). Una planta bien nutrida tendrá mejor vigor y supervivencia, comparada con una que presente deficiencias nutrimentales (Landis, 1989), pero el exceso de nutrimentos puede afectar el desarrollo de la planta y ser tóxico, por lo cual la concentración de nutrimentos es útil como estimador de calidad de planta. Un árbol puede tener una buena nutrición pero si está en un contenedor pequeño su sistema radical será deficiente ante ambientes con limitaciones de humedad o suelos pobres. Por lo tanto, este tipo de indicadores fisiológicos no debe considerarse sin tomar en cuenta indicadores morfológicos de calidad (Rodríguez, 2008).
En los viveros forestales que producen en contenedores, los sustratos como la turba de musgo, la vermiculita y la agrolita, son infértiles básicamente, lo cual es una propiedad deseable en ellos (Landis, 1990) y el viverista puede controlar la concentración nutrimental. Como referencia se puede usar el pH de los suelos que tienen cierto nivel de humedad, para comparar con el del agua de riego usado en los viveros en términos de disponibilidad de nutrientes y su concentración foliar. Esa será parte de la base para la discusión de las interacciones entre fertilización y agua de riego, para los nutrimentos usados en el presente experimento.
Lambers et al. (1998) ejemplifican la disponibilidad de nutrimentos a diferentes niveles de pH del suelo (Figura 1). La disponibilidad y, por ende, la concentración foliar de varios nutrientes a distintos niveles de pH del agua de riego, con frecuencia es semejante a las presentes en los niveles de pH del suelo. Sin embargo, en otros nutrimentos no es así pues las propiedades físico-químicas de los suelos no son las mismas que las de los sustratos preparados en el vivero para el cultivo de especies forestales (Landis et al., 1994).
Con base en los resultados mostrados en el Cuadro 6 y los niveles óptimos para coníferas de los EE. UU. usados como referencia, no hubo deficiencias nutrimentales en los tratamientos. Pero hubo diferencias significativas para la concentración foliar de K (p≤0.001) (Figura 2), porque los tratamientos con fertilización alta, independientemente del pH del agua de riego, tuvieron mayor concentración de K (Cuadro 6). La interacción pH en agua×fertilización fue significativa para K (p≤0.005) (Figura 2) y para ambos niveles de pH; la mayor fertilización aumentó la concentración foliar de K, en particular con agua cuyo pH fue 5.5. Según Van den Driessche (1984a), en los suelos muy ácidos (pH<4.5) no se retienen cationes de K, y la mayor disponibilidad de este nutrimento es con pH≥6 (Lambers et al., 1998), lo cual probablemente ocurrió en esta investigación. Los árboles que salen del vivero con mayores niveles de K foliar toleran mejor la sequía y las heladas porque influye en la apertura y cierre de estomas. Una vez que entra al sistema metabólico celular, el K forma sales con ácidos orgánicos e inorgánicos, las cuales contribuyen a regular el potencial osmótico y el contenido de agua celular (Rodríguez, 1999).
El Mg también mostró diferencias significativas (p≤0.074) y su concentración máxima en el tejido vegetal fue con pH en agua de 5.5 y fertilización alta. La concentración foliar de K aumentó y la de Mg se redujo en sustrato de P. sylvestris L. tratado con ceniza de madera y residuos de la producción de cemento para aumentar su pH (Klóšeico et al., 2014). Además, las coniferas tienen una menor capacidad que las latifoliadas para absorber cationes al aumentar el pH (Cregg, 2005), lo cual puede explicar, en parte, la menor concentración foliar de Mg con pH altos del agua de riego y del sustrato, observada en la presente investigación y por Klóšeico et al. (2014).
La mayor disponibilidad de K es otro factor que puede reducir la concentración foliar de Mg, por lo menos en el nivel alto de pH del agua en este experimento, pues en P. radiata D. Don las dosis mayores de K reducen la concentración foliar de Mg porque se abate su movilidad (Sun y Payn, 1999). Aquí puede estar involucrada la formación de un complejo con ácido málico y el Mg en la albura, y esto reduce la traslocación de Mg de raíces a parte aérea como se ha registrado para Fagus sylvatica L. El K puede tener una función en dicho enlace químico o en la producción de compuestos químicos que enlazan cationes (Schell, 1997). Aun usando agua con pH 5.5, el aporte mayor de Mg en la fertilización alta pudo contribuir a su concentración mayor en esa combinación de niveles de tratamientos.
La concentración promedio de Ca tuvo diferencias significativas (p≤0.092) y también fue significativa la interacción pH del agua y fertilización (Figura 2); así, el tratamiento con pH 5.5 y fertilización alta tuvo la concentración promedio mayor (Cuadro 5). Altland y Buamscha (2008) indican que el pH bajo no reduce la disponibilidad de Ca, como en el presente estudio donde no hubo niveles deficitarios de ese nutrimento, y que la disponibilidad de Ca está en función de otros factores y no sólo del pH. En algunos suelos minerales la disminución del pH (7) aumenta la disponibilidad de varios nutrimentos (Buamscha et al., 2007). Estos autores señalan que ácidos orgánicos liberados por la descomposición de la corteza de Pseudotsuga menziesii Mirb. (Franco) podrían reducir el pH y aumentar la disponibilidad de Ca, P, B y Fe.
Para el cobre hubo diferencias (p≤0.093) sólo entre los tratamientos extremos, T1 y T4, con la mayor concentración foliar a pH bajo y fertilización alta. Gil (1995) señala que al igual que otros cationes divalentes, este nutrimento se adsorbe fuertemente a las partículas de arcilla en forma intercambiable, pero también se halla en forma de complejos estables en las moléculas orgánicas, como ácidos fúlvicos y húmicos. El cobre reduce gradualmente su disponibilidad en los suelos alcalinos y solamente se encuentra en proporciones abundantes entre pH 5 a 7 (Lambers et al., 1998). Los resultados del presente estudio muestran que un pH alto en el agua de riego también reduce la disponibilidad del Cu, porque se proporcionó en abundancia pero las concentraciones foliares más altas ocurrieron al regar con agua de pH 5.5.
El zinc también tuvo diferencias significativas entre tratamientos (p≤0.001) al igual que la interacción entre pH del agua y la fertilización (p≤0.03), porque la concentración foliar de Zn se redujo con la mayor fertilización y agua con pH bajo, pero aumentó con la mayor fertilización y agua con pH alto (Figura 2). Según McBride (1994), el Zn tiene una regular movilidad en suelos con pH ácido, mantenida en formas intercambiables en arcillas y materia orgánica, mientras que en suelos con pH cerca de la neutralidad, la quimioabsorción en óxidos y aluminosilicatos, así como la complexación con el humus, reducen marcadamente la solubilidad del Zn. No obstante, aunque la movilidad de los cationes Zn2+ es pequeña en suelos ligeramente alcalinos, complejos orgánicos de Zn se pueden solubilizar y aumentar la disponibilidad de esos cationes. Ello puede ayudar a explicar por qué la mayor fertilización en este estudio aumenta la concentración foliar de Zn con pH= 8.
Para el fósforo se esperaría que al aplicar ácido fosfórico para reducir el pH del agua de riego, las concentraciones foliares mayores de P ocurrirían en las plantas con dicho tratamiento, pero no fue así. Van den Driessche (1984b) usó el mismo ácido para manejar el pH del agua y como fuente de P en la producción de Pseudotsuga menziesii var. menziesii, pero tampoco halló relación con la concentración foliar de este nutriente. En el presente estudio las concentraciones foliares de N, P y B no fueron estadísticamente diferentes entre tratamientos y el Mn sólo fue diferente entre T2 y T3. Las interacciones pH agua de riego y fertilización tampoco fueron significativas para las concentraciones foliares de N, P, Mg, Cu, Mn y B. Para N la disponibilidad cambia relativamente poco en el intervalo de pH estudiado, y el P muestra más variabilidad, pero en los niveles de pH estudiados su disponibilidad es similar (Lambers et al., 1998). Estos seis nutrimentos tuvieron concentraciones foliares superiores al mínimo recomendado (Cuadro 6). Manganeso y boro rebasaron el máximo pero aparentemente no llegaron a niveles tóxicos en la especie estudiada.
Al comparar resultados de la literatura consultada con los del presente estudio, la concentración de varios nutrimentos no mostró una tendencia típica. En parte esto puede ser porque los objetos de estudio fueron coníferas, cuya respuesta sería diferente a las latifoliadas o las especies de cultivos agrícolas, y en estas últimas se ha realizado más investigación sobre nutrición vegetal.
CONCLUSIONES
Los niveles de fertilización y pH del agua de riego estudiados no afectaron los indicadores morfológicos de calidad de Pinus cembroides, pero sí hubo efectos en las concentraciones foliares de K y Ca. Estos dos nutrimentos son importantes porque contribuyen a la resistencia a sequía y a bajas temperaturas, comunes en los ambientes donde la especie es plantada. Por lo tanto, en los viveros que produzcan Pinus cembroides el agua debe ser acidificada y usar la fertilización alta como en el presente estudio porque esta combinación causó las mayores concentraciones foliares de K y Ca.
Todos los tratamientos presentaron concentraciones nutrimentales dentro de un intervalo considerado óptimo, salvo el Mn, que tuvo niveles de consumo en exceso, pero no tóxicos.
AGRADECIMIENTOS
El primer autor agradece al posgrado de la DICIFO, UACH, por permitirle realizar estudios en su programa de Maestría en Ciencias en Ciencias Forestales. También agradece al CONACYT por la beca proporcionada, a la UACH, por el apoyo para la realización de los análisis nutrimentales, proyecto no. 137902003 (Las plantaciones en la restauración), al vivero y al Laboratorio de Semillas Forestales de la DICIFO, por el apoyo recibido durante la investigación.
LITERATURA CITADA
Altland, J. E., and M. G. Buamscha. 2008. Nutrient availability from Douglas fir bark in response to substrate pH. Hortscience 43(2): 478-483. [ Links ]
Buamscha, M. G., J. E. Altland, D. M. Sullivan, D. A. Horneck, and J. Cassidy. 2007. Chemical and physical properties of Douglas fir bark relevant to the production of container plants. Hortscience 42(5): 1281-1286. [ Links ]
Cregg, B. 2005. Conifer nutrition. The Michigan Landscape Sep.-Oct.: 42-45. [ Links ]
Dickson, A., A. L. Leaf, and J. F. Hosner. 1960. Quality appraisal of white spruce and white pine seedlings stock in nurseries. Forestry Chronicle 36: 10-13. [ Links ]
Duryea, M. L. 1985. Evaluating seedling quality: importance to reforestation. In: Duryea, M. L. (ed). Evaluating Seedling Quality: Principles, Procedures and Predictive Abilities of Major Tests. Forest Research Laboratory. Corvallis, USA. pp: 1-4. [ Links ]
Farjon, A., J. A. Pérez de la R., y B. T. Styles. 1997. Guía de Campo de los Pinos de México y América Central. The Royal Botanic Gardens, Universidad de Oxford. Bélgica. 151 p. [ Links ]
García, E. 1978. Modificaciones al Sistema de Clasificación Climática de Köppen. 2a. ed, Instituto de Geografía, Universidad Nacional Autónoma de México. México. 246 p. [ Links ]
García S. J., P., J. J. Lucena M., S. Ruano C., y M. Nogales G. 2009. Guía práctica de la fertilización racional de los cultivos agrícolas de España. Ministerio de Medio Ambiente y Medio Rural y Marino. España. 119 p. [ Links ]
Gil, M. F. 1995. Elementos de Fisiología Vegetal: Relaciones Hídricas, Nutrición Mineral, Transporte y Metabolismo. Mundi Prensa. México. 1147 p. [ Links ]
Infante G., S., y P. L. Zárate G. 2000. Métodos Estadísticos: Un Enfoque Interdisciplinario. Trillas. México, D. F. 684 p. [ Links ]
Klõšeico, J., T. Kuznetsova, M. Tilk, and M. Mandre. 2014. Short-term influence of Clinker dust and wood ash on macronutrients and growth in Norway spruce (Picea abies) and Scots pine (Pinus sylvestris) seedlings. Comm. Soil Sci. Plant Analysis 45(16): 2105-2117. [ Links ]
Lambers, H., F. S. Chapin III, and T. L. Pons. 1998. Plant Physiological Ecology. Springer. New York. 540 p. [ Links ]
Landis, T. D. 1985. Mineral nutrition as an index of seedling quality. In: Duryea, M. L. (ed). Evaluating Seedling Quality: Principles, Procedures, and Predictive Abilities of Major Tests. Oregon State University. Corvallis, USA. pp: 29-48. [ Links ]
Landis, R. D. 1989. Mineral nutrients and fertilization. In: Landis, T. D., R. W. Tinus, S. E. McDonald, and J. P. Barnett (eds). The Container Tree Nursery Manual. Vol. 4. Agriculture Handbook 674. USDA Forest Service. Washington, D. C. pp: 72-126. [ Links ]
Landis, R. D. 1990. Containers and growing media. In: Landis, T. D., R. W. Tinus, S. E. McDonald, and J. P. Barnett (eds). The Container Tree Nursery Manual. Vol. 2. Agriculture Handbook 674. USDA Forest Service. Washington, D. C. pp: 41-85. [ Links ]
Landis, T. D., R. W. Tinus, S. E. McDonald, and J. P. Barnett. 1994. Manual de Viveros para la Producción de Especies Forestales en Contenedor. Vol 1. Manual Agrícola 674. Departamento de Agricultura de los Estados Unidos de América, Servicio Forestal. México. 188 p. [ Links ]
McBride, M. B. 1994. Environmental Chemistry of Soils. Oxford University Press. New York. 406 p. [ Links ]
Mendoza, S. I. 2009. Calidad de las aguas residuales urbano-industriales que riegan el valle de mezquital, Hidalgo, México. Tesis de Doctorado. Colegio de Postgraduados. Montecillo, Texcoco, Estado de México. 195 p. [ Links ]
Peñuelas, R. J., y B. L. Ocaña. 1994. Cultivo de Plantas Forestales en Contenedor. 2ª ed. Mundi-Prensa. Madrid, España. 190 p. [ Links ]
Perry Jr., J. P. 1991. The Pines of Mexico and Central America. Timber Press. Portland, USA. 231 p. [ Links ]
Prieto R., J. A., E. H. Cornejo O., P. A. Domínguez C., J. de J. Návar Ch., J. G. Marmolejo M., y J. Jiménez P. 2004. Estrés hídrico en Pinus engelmannii Carr., producido en vivero. Investigación Agraria. Sistemas y Recursos Forestales 13(3): 443-451. [ Links ]
Quiroz, I., E. García, M. González, P. Chung, K. Casanova, y H. Soto. 2009. Calidad de planta y certificación. Centro tecnológico de la planta forestal. U. A. N. L. Nuevo León, México. 5 p. [ Links ]
Rodríguez S., F. 1999. Fertilizantes. Nutrición Vegetal. A. G. T. Ed. México, D. F. 157 p. [ Links ]
Rodríguez T., D. A. 2008. Indicadores de Calidad de Planta Forestal. Mundi-Prensa. México, D. F. 156 p. [ Links ]
Ruano M., R. 2003. Viveros Forestales. Mundi-Prensa. Madrid, España. 279 p. [ Links ]
Schell, J. 1997. Interdependence of pH, malate concentration, and calcium and magnesium concentrations in the xylem sap of beech roots. Tree Physiol. 17: 479-483. [ Links ]
Sun, O. J., and T. W. Payn. 1999. Magnesium nutrition and photosynthesis in Pinus radiata: clonal variation and influence of potassium. Tree Physiol. 19: 535-540. [ Links ]
Thompson, B. E. 1985. Seedling morphology: what you can tell by looking. In: Duryea, M. L. (ed). Evaluating Seedling Quality. Principles, Procedures, and Predictive Abilities of Major Tests. Corvallis, USA. pp: 59-71. [ Links ]
Van den Driessche, R. 1984a. Soil fertility in forest nurseries. In: Duryea, M. L., and T. D. Landis (eds). Forest Nursery Manual: Production of Bareroot Seedlings. Martinus Nijhoff/Dr. W. Junk Pubs. The Haghe. pp: 63-74. [ Links ]
Van den Driessche, R. 1984b. Response of Douglas fir seedlings to phosphorus fertilization and influence of temperature on this response. New Forests 50: 155-169. [ Links ]
Vázquez Y., C., A. I. Batis M., M. I. Alcocer S., M. Gual D., y C. Sánchez D. 1999. Árboles y arbustos potencialmente valiosos para la restauración ecológica y la reforestación. Reporte técnico del proyecto J084. CONABIO, Instituto de Ecología, UNAM. México. [ Links ]

