










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgrociencia
On-line version ISSN 2521-9766Print version ISSN 1405-3195
Agrociencia vol.48 n.8 Texcoco Nov./Dec. 2014
Ciencia animal
Efecto del nivel de concentrado sobre el perfil de ácidos grasos de la leche de vacas Holstein en pastoreo
Effect of level of concentrate on milk fatty acid profile from grazing Holstein cows
Horacio Castro-Hernández1, F. Filiberto González-Martínez1, I. Arturo Domínguez-Vara1, J. Manuel Pinos-Rodríguez2, Ernesto Morales-Almaráz1*, Rodolfo Vieyra-Alberto1
1 Facultad de Medicina Veterinaria y Zootecnia. Universidad Autónoma del Estado de México. Instituto Literario 100 Ote. Centro. Toluca, México. * Autor responsable (maernesto@hotmail.com).
2 Centro de Biociencias, Universidad Autónoma de San Luis Potosí, México.
Recibido: mayo, 2014.
Aprobado: noviembre, 2014.
Resumen
El contenido de ácido linoléico conjugado (CLA c9 t11) en leche depende principalmente de la producción en el rumen de ácido vaccénico, el cual está influenciado por el aporte de los ácidos linoléico y linolénico en la dieta. El objetivo de este estudio fue evaluar el efecto del nivel de concentrado en la producción, composición y perfil de ácidos grasos de la leche de nueve vacas Holstein en praderas asociadas de gramíneas y leguminosas. El diseño experimental fue un cuadro latino 3x3 y los tratamientos fueron: 1) 8 kg concentrado y 8 h en la pradera (8c); 2) 5 kg concentrado y 12 h en la pradera (5c), y 3) 3 kg de concentrado y 12 h en la pradera (3c). Los datos se analizaron con el procedimiento MIXTO de SAS y las medias de los tratamientos se compararon con la prueba de Tukey (p≤0.05). La producción de leche y el contenido de grasa, proteína y lactosa fueron mayores (p≤0.05) en el tratamiento 8c. La concentración de los ácidos grasos (AG) láurico, mirístico y palmítico se incrementó en la leche (p≤0.05) al aumentar el nivel de concentrado. El contenido de los AG de cadena larga (≥C18) fue mayor (p≤0.05) con 3 kg de concentrado, excepto para C18:2 c9c12. La reducción del concentrado suministrado a vacas Holstein en praderas asociadas disminuye el contenido de AG saturados, pero aumenta el contenido de los insaturados, especialmente de los ácidos oleico, linolénico, vaccénico y CLA c9 t11.
Palabras clave: bovinos, ácido linoléico conjugado, leche, pastoreo.
Abstract
The content of conjugated linoleic acid (c9 t11 CLA) in milk depends mainly on the production in rumen of vaccenic acid, which is influenced by the supply of linoleic and linolenic acids in the diet. The aim of this study was to evaluate the effect of the level of concentrate on the production, composition and fatty acid profile of milk from nine Holstein cows in prairies with grass and legumes. The experimental design was a 3x3 Latin square and the treatments were: 1) 8 kg concentrate and 8 h on the prairie (8c); 2) 5 kg concentrate and 12 h on the prairie (5c), and 3) 3 kg of concentrate and 12 h on the prairie (3c). The data were analyzed with the SAS MIXED procedure and treatment means were compared with the Tukey test (p≤0.05). Milk yield and fat, protein and lactose content were higher (p≤0.05) in 8c treatment. The concentration of lauric, myristic and palmitic fatty acids (FA) increased in milk (p≤0.05) with the increase of the concentrate level. The content of long-chain FAs (≥C18) was higher (p≤0.05) with 3 kg of concentrate, except for C18:2 c9c12. The reduction of the concentrate supplied to Holstein cows in prairies decreased the content of saturated FA, but increased the content of the unsaturated, particularly oleic, linolenic, vaccenic and c9 t11 CLA acids.
Keywords: cattle, conjugated linoleic acid, milk, grazing.
INTRODUCCIÓN
La grasa de la leche contiene 70 a 75 % de ácidos grasos (AG) saturados (AGS) (Lock y Shingfield, 2004), los cuales están relacionados con efectos hipercolesteromiantes y enfermedades cardiacas, en especial los ácidos láurico, mirístico y palmítico (Williams, 2000); sin embargo la grasa de la leche también contiene 5 % de AG poliinsaturados (AGPI) (Lock y Shingfield, 2004) como el ácido linoleico con acción hipocolesteromiante en seres humanos (Grundy, 1994) y el isómero C18:2 c9 t11 (ruménico) del ácido linoleico conjugado, con propiedades anticarcinogénicas en modelos animales (Ip et al., 1999; Corl et al., 2003) y posiblemente en humanos (Belury, 2002). Por lo tanto, es importante aumentar la concentración de AGPI y disminuir la de AGS en la leche, productos lácteos y otros alimentos para consumo humano.
El perfil lipídico de la leche es afectado por la dieta (Shingfield et al., 2005), la raza animal (Carroll et al., 2006), la época del año (Lock y Garnsworthy, 2003), o la etapa de lactancia (Auldist et al., 1998). Además, la concentración de AGPI aumenta con el pastoreo (Kelly et al., 1998), el tipo de ensilado (Dewhurst et al., 2003), la cantidad de concentrado en la dieta (Bargo et al., 2006), la fuente de granos (Wijesundera et al., 2003) y el suministro de aceites de oleaginosas (Khanal et al., 2002) o de aceites de pescado (Abu-Ghazaleh et al., 2003).
Según Albarrán-Portillo (1999) y Peña-Carmona (1999), citados por Arriaga-Jordán et al. (2001), el desempeño productivo es mejor en vacas lecheras alimentadas con forrajes y cantidades limitadas de concentrado (3.0 kg vaca-1 d-1), comparado con la alimentación tradicional de cantidad baja de forraje y alta de concentrado (de 5.0 a 9.0 kg vaca-1 d-1). De acuerdo con Arriaga-Jordán et al. (2001), la limitación del aporte de concentrado complementario al pastoreo en el ganado lechero puede cubrir los requerimientos nutricionales de las vacas.
En el altiplano de México, la evaluación de estrategias de alimentación en ganado lechero está dirigida hacia el estudio del desempeño económico y productivo principalmente, y está basada en el pastoreo de praderas mejoradas (Lolium perenne y Trifolium repens) y un suplemento de concentrado comercial (Arriaga-Jordán et al., 2001; 2002) y de ensilados (Garduño Castro et al., 2007; Hernández-Ortega et al., 2011). El contenido de AGPI en la leche de vaca aumenta al incluir forraje fresco en la dieta (Kelly et al., 1998; Dewhurst et al., 2006) y con más tiempo de pastoreo (Morales-Almaráz et al., 2010). En la literatura revisada hay poca información respecto a la modificación mediante estrategias de alimentación del contenido de AG en la leche en México.
La reducción de la cantidad de concentrado a expensas de asignar mayor tiempo de acceso a la pradera y suministrar ensilado de maíz, podría proporcionar un mayor aporte de AGPI, principalmente C18:3 desde el pasto y C18:2 desde el ensilado de maíz, para mejorar el perfil lipídico de la leche. Por lo tanto, el objetivo de la presente investigación fue evaluar el efecto de tres niveles de concentrado y el tiempo de acceso a la pradera sobre la producción, composición y perfil de AG de la leche de vacas Holstein en el Valle de Toluca, México.
MATERIALES Y MÉTODOS
Área de estudio
El experimento se realizó durante mayo y junio de 2011, en la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Autónoma del Estado de México, ubicada a 19° 24' 48'' O, 99° 40' 45'' N y una altitud de 2632 m (INEGI, 2003). El clima es templado subhúmedo con lluvias en verano, clasificado como Cb (W2) (W2) (García, 1987).
Animales, dieta y tratamientos
Nueve vacas Holstein multíparas (22.6±4.0 kg leche d-1 y 99.0 ±42.0 d en lactancia) fueron usadas en el estudio con tres periodos experimentales. La alimentación de las vacas estuvo basada en un sistema de producción mixto con tiempos de acceso al pastoreo más un suplemento de ensilado de maíz y concentrado en el establo. El pastoreo fue rotacional en praderas polífitas de 2.5 ha. Las especies predominantes fueron Lolium perenne, Trifolium repens, Festuca arundinacea, Dactylis glomerata y Pennisetum clandestinum. Las praderas fueron fertilizadas con 50 kg urea ha-1. El concentrado contenía (BS) 18 % de proteína cruda (PC) y 2.0 Mcal kg-1 MS de energía neta (EN) para lactancia (ENL), y fue elaborado con (g kg-1 MS) sorgo (486.7), pasta de soya (200.4), canola (147.5), salvado de trigo (146.7) más una premezcla de vitaminas y minerales (18.7). El concentrado (kg MS vaca- 1 d- 1) se proporcionó asociado al tiempo de acceso a la pradera y fue: 1) 7.2 kg de concentrado (8c) con acceso a la pradera de 07:00 a 15:00 h; 2) 4.5 kg de concentrado (5c) con acceso a la pradera de 7:00 a 15:00h y de 16:00 a 20:00 h; 3) 2.7 kg de concentrado (3c) con acceso a la pradera igual a 5c. El concentrado se ofreció a las 16:00 y 20:00 h para el tratamiento 1, y a las 20 y 24 h en los tratamientos 2 y 3. El ensilado de maíz se ofreció a libre acceso en los mismos tiempos. Las vacas tuvieron libre acceso al agua de bebida en la pradera y en el establo.
Desarrollo experimental
El estudio comprendió tres periodos experimentales, cada uno con 14 d de adaptación y 5 d de muestreo. Los ordeños se realizaron a las 06:00 y 15:00 h. La asignación de forraje en pastoreo fue 25 kg de MS vaca-1 d-1, determinada mediante medición de la biomasa antes del pastoreo, para lo cual el forraje se cortó en ocho cuadrantes (0.25 m2 cuadrante-1) a ras de suelo, se pesó y homogenizó el material fresco, se tomó la muestra y se secó a 60 °C en estufa de aire forzado.
La superficie de pastoreo fue ajustada cada día durante la etapa de medición. El manejo fue el siguiente: finalizado el ordeño matutino (07:00 h), las nueve vacas fueron llevadas a la pradera, donde permanecieron juntas hasta iniciar ordeño vespertino (15:00 h), y al término (16:00 h), las vacas del grupo 8c fueron estabuladas y recibieron el concentrado y el ensilado de maíz; mientras que las vacas de los grupos 5c y 3c se llevaron de nuevo a la pradera donde permanecieron hasta las 20:00 h; después fueron estabuladas y recibieron el concentrado y el ensilado de maíz. Para los tres grupos, la alimentación en la estabulación fue a las 20:00 y 24:00 h en comederos individuales, y para el grupo 8c una comida previa a las 16:00 h y en la comida de las 24:00 h solo se suministró ensilado de maíz.
Cada día se midió el consumo de alimento (ensilado de maíz y concentrado) en estabulación por diferencia de la oferta y el rechazo. El consumo de pasto se determinó mediante el método descrito por Macoon et al. (2003), para lo cual se calculó los requerimientos de ENL total del ganado lechero según las ecuaciones del NRC (2001), y el aporte de energía neta de los alimentos consumidos en el establo. El contenido de EN del pasto, ensilado de maíz y concentrado fue calculado con las ecuaciones descritas por Menke y Steingass (1988), a partir del contenido de fibra ácido detergente. La producción de leche se midió cada día, la leche fue muestreada individualmente en ambos ordeños, y se obtuvo una alícuota (50 mL) la cual estuvo congelada (-20 °C) hasta su análisis.
Análisis de laboratorio
En los alimentos se determinó el contenido de MS; materia orgánica (MO) y (PC) (AOAC, 2012); fracciones de fibra: neutro detergente (FND), ácido detergente (FAD) y lignina ácido detergente (LAD), de acuerdo con Van Soest et al. (1991). El aporte de EN de los alimentos se estimó según las ecuaciones descritas por Menke y Steingass (1988); el contenido de los AG de los alimentos (Cuadro 1) se determinó mediante la técnica descrita por Sukhija y Palmquist (1988) con modificaciones de Palmquist y Jenkins (2003) usando ácido clorhídrico metanólico al 10 % y hexano como solvente orgánico.
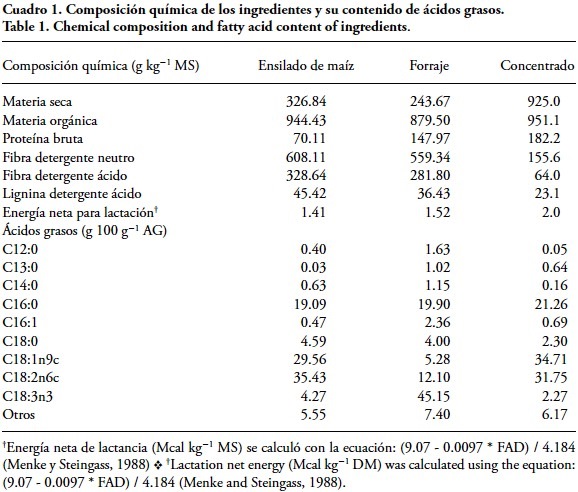
Para medir el contenido de proteína, grasa y lactosa en la leche se utilizó un analizador Lactoscan SL60. El perfil de AG de la leche se determinó previa extracción de la grasa de acuerdo con la técnica descrita por Feng et al. (2004) La metilación de la muestra se usando la metodología reportada por Christie (1982) con modificaciones de Chouinard et al. (1999). Los ésteres metílicos de los AG de la leche y los alimentos fueron separados y cuantificados por cromatografía de gases (Perkin Elmer Clarus 500) con una columna capilar de 100 m x 0.25 mm x 0.2 μm (SUPELCO TM-2560) utilizando nitrógeno como gas acarreador. El detector e inyector se mantuvieron a 260 °C, la temperatura inicial del horno fue 140 °C por 5 min, aumentando 4 °C por minuto hasta llegar a 240 °C. Cada pico fue identificado de acuerdo con los tiempos de retención de estándares de esteres metílicos (Supelco 37, FAME MIX analytical; trans-11-octadecenoic methyl ester; linoleic acid conjugated methyl ester SIGMA USA).
Análisis estadístico
El diseño experimental fue un cuadro latino 3x3 (n=3) y los datos se analizaron con el procedimiento MIXTO de SAS (2002). El modelo estadístico fue el siguiente:
Yijk = μ + Pi + Gj + Ck + Ez
donde Yijkz es la variable dependiente, μ es la media general, Pi es el efecto del periodo (1, 2, 3), Gj es el efecto del grupo (1, 2, 3), Ck es el efecto del nivel de concentrado (3, 5, y 8 kg MF), Ez es el error residual.
Las medias de los tratamientos se compararon con la prueba de Tukey (p≤0.05) (Steel et al., 1997).
RESULTADOS Y DISCUSIÓN
Variables productivas
Los datos de consumo, producción y composición de la leche se presentan en el Cuadro 2. El tratamiento 8c causó la mayor ingestión (p≤0.05) de ensilado de maíz y el consumo total en estabulación con respecto a los otros tratamientos. El consumo de pasto en la pradera fue 1.7 y 2.7 veces más (p≤0.05) para las vacas en los tratamientos 5c y 3c, respectivamente, respecto a las vacas en el tratamiento 8c. De acuerdo con el NRC (2001), vacas lecheras con un peso vivo promedio de 680 kg, con 90 d promedio de lactancia y una producción de 25 kg de leche d-1 necesitan consumir 19.6 kg MS, mientras que las vacas de 450 kg de PV, con producción de 20 kg de leche d-1, requieren consumir 16.5 kg de MS. En el presente estudio, la producción láctea no rebasó los 20.0 kg d-1 por lo cual se esperaría consumos de MS menores a 16 kg d-1 (NRC, 2001), y las vacas de los tratamientos 5c y 8c tuvieron un consumo similar, pero estadísticamente inferior al de las vacas del tratamiento 3c (Cuadro 2).

Al aumentar el concentrado, la producción de leche aumentó 10 y 23 % en 5c y 8c respecto a 3c (Cuadro 2) pero sin afectar (p>0.05) el contenido de grasa (36.63±8.86 g kg-1), de proteína (31.25±5.55 g kg-1) y de lactosa (43.83 ±3.77 g kg-1) en la leche.
El rendimiento de grasa, proteína y lactosa fue superior en 8c (p≤0.05), lo cual se debe al mayor aporte de energía en la dieta al aumentar el nivel de concentrado, lo cual se reflejó en un aumento de la producción de leche, sin afectar los componentes de la leche. Bargo et al. (2002) observaron respuestas similares en el rendimiento de los componentes en la leche de vacas que recibieron un concentrado (1 kg concentrado 4 kg-1 leche) versus sin concentrado.
Perfil de ácidos grasos
El nivel de concentrado afectó (p≤0.05) a la mayoría de los AG de la leche (Cuadro 3). Los AG de cadena corta (C4-C11) tuvieron mayor concentración en la leche de 8c, excepto C4 y C6. Al aumentar el nivel de concentrado de 3 a 8 kg, los AG láurico (C12), mirístico (C14) y palmítico (C16) elevaron (p≤0.05) su concentración en 24.4, 8.5 y 4.8 %, respectivamente, lo cual contribuyó al mayor contenido total de AG en la leche (68.00 vs 69.76 g 100 g-1 AG, respectivamente para 3c y 8c). En contraste, Nielsen et al. (2006) no observaron efecto del nivel alto o bajo de concentrado, a base de grano de cebada y pasta de colza, en la concentración de los AG C12:0, C14:0 y C16:0 en leche de vacas alimentadas con ensilado de maíz y pasto como única fuente de forraje. En la presente estudio, el aumento en el contenido de AG de cadena corta y media para 8c fue similar al observado por Stockdale et al. (2003) en vacas en pastoreo más 7 y 9 kg d-1 de concentrado: aumentaron los AG de cadena corta C6:0 a C10:0 y de cadena media C12:0 a C16:0 con respecto a vacas sin suplemento. Wijesundera et al. (2003) proporcionaron 4.5 kg d-1 de granos de cereales a vacas en pastoreo y la concentración de C10:0 a C14:0 aumentó en la grasa de la leche. Según Bargo et al. (2006), al dar 1 kg concentrado 4 kg-1 leche a vacas en pastoreo, la concentración de C12:0 y C14:0 fue mayor en comparación con vacas sin concentrado (2.50 vs 1.80 y 9.31 vs 7.59 g 100 g-1 AG).

En los AG de cadena media, no se observaron diferencias (p>0.05) en la concentración de ácido palmitoleico (C16:1 c9), C15 y C15:1 excretados en la leche.
Con excepción del ácido linoléico (C18:2 c9 c12), el contenido de AG de cadena larga en leche (>C17) fue mayor (p≤0.05) con el nivel más bajo de concentrado. El contenido de ácido vaccénico (C18:1 t11) en la leche fue superior (p≤0.05) en 3c: 9.3 % mayor que 5c y 12.3 % mayor que 8c. De forma similar, el contenido de ácido linoleico conjugado (ALC) c9 t11 fue 9.8 % mayor en 3c comparado con los otros dos tratamientos. Nielsen et al. (2006) observaron que el contenido de ALC c9 t11 (1.61 vs 1.17 g 100 g-1 AG) y ácido vaccénico (2.80 vs 1.55 g 100 g-1 AG) fue mayor en la leche de vacas alimentadas un nivel bajo de concentrado, pero no hubo diferencias en la concentración de ácido esteárico, oleico y linolénico por efecto del nivel de concentrado. Para 3c el contenido de ácido oleico (C18:1 c9) y C18 fue mayor (p≤0.05) en 7.8 y 22.4 %, respectivamente, respecto a 8c.
El incremento de C18:2 c9 c12 en la grasa de la leche en vacas del tratamiento 8c, comparado con los otros dos tratamientos, se debe al consumo mayor de ácido linoleico porque hubo más consumo de ensilado de maíz, el cual es una importante fuente de C18:2 c9 c12, y de los ácidos palmítico y oleico. Los AG del ensilado de maíz tienen una concentración alta de ácidos linoleico (Nielsen et al., 2006) y oleico, pero baja de ácido linolénico (Dewhurst et al., 2006), similar a lo reportado en el Cuadro 1.
El aumento en el consumo de MS de forrajes frescos puede tener efectos importantes sobre la composición de los AG de la grasa de la leche (Kelly et al., 1998; Bargo et al., 2006; Morales-Almaráz et al., 2010). La leche de las vacas que recibieron 2.7 kg concentrado (3c) mostró altas (p≤0.05) concentraciones de los ácidos esteárico, oleico y vaccénico (Cuadro 3) comparado con las vacas en 8c, y en especial con 5c. Dado que el tiempo de acceso a la pradera fue el mismo, el suministro de 2 kg más de concentrado a las vacas pudo cambiar el metabolismo ruminal de los lípidos, porque un suplemento alto en concentrado afecta la biohidrogenación debido al bajo pH ruminal (Lathamn et al., 1972; Bargo et al., 2006), lo cual cambia la producción de AG en el rumen y en la glándula mamaria (Enjalbert et al., 2008). Además, el efecto en el contenido en leche de los ácidos C18, incluido el CLA c9 t11 o ácido ruménico, podría estar relacionado con la variación individual en la eficiencia en la síntesis de novo en la glándula mamaria, porque la dieta tiene una función determinante en el contenido de ácido ruménico en la grasa de la leche de vacas, pero hay un efecto significativo de las diferencias entre individuos (Peterson et al., 2002), lo cual radica en la eficiencia de desaturación para sintetizar de novo CLA c9 t11 desde ácido vaccénico, principal ruta de síntesis del CLA c9 t11 excretado en la leche (Griinari et al., 2000).
Bargo et al. (2006) observaron que el suplemento con concentrado tuvo un efecto negativo en los AG de cadena larga, oleico y linolénico, cuya concentración fue mayor en vacas sin suplemento (30.89 vs 27.18 y 1.16 vs 0.79 g 100 g-1 AG, respectivamente); un efecto similar fue observado en el presente estudio al disminuir el nivel de concentrado. Asimismo, las concentraciones de los ácidos vaccénico y ALC c9 t11 (1.18 vs 1.36 y 2.79 vs 3.48 g 100 g-1 AG) disminuyeron en la leche de vacas que recibieron concentrado, respecto a vacas sin concentrado (Bargo et al., 2006), lo cual es similar a lo observado en la presente investigación.
La concentración total de AGS en la leche aumentó (p≤0.05) y la de AGI disminuyó debido al nivel de concentrado, resultado similar a lo reportado por Bargo et al. (2006). El suministro de 3.0 kg de concentrado podría mejorar la calidad de la leche debido a su menor concentración de AGS y su implicación como factores de riego en enfermedades cardiovasculares (Williams, 2000). Además hay una concentración mayor de ácidos oleico, linolénico, vaccénico y ALC c9 t11, benéficos por su acción hipocolesteromiante (Grundy, 1994).
CONCLUSIONES
La reducción de la cantidad de concentrado de 8.0 a 3.0 kg d-1 a vacas en pastoreo y el aumento de tiempo de acceso a la pradera, de 8 a 12 h, disminuyó la producción de leche. Sin embargo, el aporte de ácido linoléico y linolénico a las vacas contribuyó a mejorar el perfil de AG al aumentar el contenido de los ácidos oleico, linolénico, vaccénico y CLA c9 t11, lo cual resulta en un producto más saludable para el consumidor.
LITERATURA CITADA
AbuGhazaleh, A. A., D. J. Schingoethe, A. R. Hippen, and E. K. Kalscheur. 2003. Milk conjugated linoleic acid response to fish oil supplementation of diets differing in fatty acids profile. J. Dairy Sci. 86: 944-953. [ Links ]
Association of Official Analytical Chemists. 2012. Official Methods of Analysis. 19th ed. AOAC. Arlington. VA, USA. pp: 34-36. [ Links ]
Arriaga-Jordán, C. M., F. J. Flores-Gallegos, G. Peña-Carmona, B. Albarrán-Portillo, A. García-Martínez, A. Espinoza-Ortega, C. E. González-Esquivel, and O. A. Castelán-Ortega. 2001. Participatory on-farm evaluation of the response to concentrate supplementation by cows in early lactation in smallholder peasant (campesino) dairy production systems in the highlands of central Mexico. J. Agric. Sci. 137: 97-103. [ Links ]
Arriaga-Jordán, C. M., B. Albarrán-Portillo, A. Espinoza-Ortega, A. García-Martínez, and O. A. Castelán-Ortega. 2002. On-farm comparison of feeding strategies based on forages for small-scale Dairy production systems in the highlands of Central Mexico. Exp. Agric. 38: 375-388. [ Links ]
Auldist, M. J., B. J. Walsh, and N. A. Thomson. 1998. Seasonal and lactational influences on bovine milk composition in New Zeland. J. Dairy Res. 65: 401-411. [ Links ]
Bargo, F., J. E. Delahoy, G. F. Schroeder, and L. D. Muller. 2006. Milk fatty acid composition of dairy cows grazing at two pastures allowance and supplemented with different levels and sources of concentrate. Anim. Feed Sci. Technol. 125: 17-31. [ Links ]
Bargo F., L. D. Muller, J. E. Delahoy, and T. W. Cassidy. 2002. Milk response to concentrate supplementation of high producing dairy cows at two pasture allowances. J. Dairy Sci. 85: 1777-1792. [ Links ]
Belury, M. A. 2002. Dietary conjugated linoleic acid in health: physiological effects and mechanisms of action. Annu. Rev. Nutr. 22: 505-531. [ Links ]
Carroll, S. M., E. J. DePeters, S. J. Taylor, H. Rosenberg, H. Perez-Monti, and V. A. Capps. 2006. Milk composition of Holstein, Jersey, and Brown Swiss cows in response to increasing levels of dietary fat. Anim. Feed Sci. Technol. 131: 451-473. [ Links ]
Chouinard, P. Y., L. Corneau, D. M. Barbano, L. E. Metzger, and D. E. Bauman. 1999. Conjugated linoleic acids alter milk fatty acid composition and inhibit milk fat secretion in dairy cows. J. Nutr. 129: 1579-1584. [ Links ]
Christie, W. W. 1982. A simple procedure for rapid transme-thylation of glycerolipids and cholesterol esters. J. Lipid Res. 23: 1072-1075. [ Links ]
Corl, B. A., D. M. Barbano, D. E. Bauman, and C. Ip. 2003. Cis-9, trans-11 CLA derived endogenously from trans-11 18:1 reduces cancer risk in rats. J. Nutr. 133: 2893-2900. [ Links ]
Dewhurst R. J., K. F. Shingfield, M. R. F. Lee, and N. D. Scollan. 2006. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems. Anim. Feed Sci. Technol. 131: 168-206. [ Links ]
Dewhurst, R. J., N. D. Scollan, M. R. F. Lee, H. J. Ougham, and M. O. Humphreys. 2003. Forage dreading and management to increase the beneficial fatty acid content of ruminant products. Proc. Nutr. Soc. 62: 329-336. [ Links ]
Enjalbert, F., Y. Videau, M. C. Nicot, and A. Troegeler-Meynadier. 2008. Effects of induced subacute ruminal acidosis on milk fat content and fatty acid profile. J. Anim. Physiol. Anim. Nutr. 92(3): 284-291. [ Links ]
Feng, S., A. L. Lock, and P. C. Garnsworthy. 2004. Technical note: A rapid lipid separation method for determining fatty acid composition of milk. J. Dairy Sci. 87: 3785-3788. [ Links ]
García, E. 1987. Modificaciones al Sistema de Clasificación Climática de Kóppen. 4a. Ed., México, D. F. 217 p. [ Links ]
Garduño-Castro, Y., A. Espinoza-Ortega, C. E. González-Esquivel, B. Mateo-Salazar, and C. M. Arriaga-Jordán. 2007. Intercropped oats (Avena sativa) - common vetch (Vicia sativa) silage in the dry season for small-scale dairy systems in the Highlands of Central Mexico. Trop. Anim. Health Prod. 41: 827-834. [ Links ]
Griinari J. M., B. A. Corl, S. H. Lacy, P. Y. Chouniard, K. V. V. Nurmela, and D. E. Bauman. 2000. Conjugated linoleic acid is synthesized endogenously in lactating cows by delta 9-desaturase. J. Nutr. 130: 2285-2291. [ Links ]
Grundy, S. M. 1994. Influence of stearic acid on cholesterol metabolism relative to other long-chain fatty acids. Am. J. Clin. Nutr. 60: 986-1110. [ Links ]
Hernández-Ortega, M., D. Heredia-Nava, A. Espinoza-Ortega, E. Sánchez-Vera, and C. M. Arriaga-Jordán. 2011. Effect of silage from ryegrass intercropped with winter or common vetch for grazing dairy cows in small-scale dairy systems in Mexico. Trop. Anim. Health Prod. 43: 947-954. [ Links ]
INEGI. 2003. Anuario Estadístico. México. Gobierno del Estado de México. México. 230 p. [ Links ]
Ip, C., S. Banni, E. Angioni, G. Carta, J. McGinley, H. J. Thompson, B. Barbano, and D. E. Bauman. 1999. Conjugated linoleic acid-enriched butterfat alters mammary gland morphogenesis and reduces cancer risk in rats. J. Nutr. 129: 2135-2142. [ Links ]
Kelly, M. L., E. S. Kolver, D. E. Bauman, M. E. Van Amburgh, and L. D. Muller. 1998. Effect of intake of pasture on concentrations of conjugated linoleic acid in milk of lactating cows. J. Dairy Sci. 81: 1630-1636. [ Links ]
Khanal, R. C., T. R. Dhiman, D. J. McMahon, and R. L. Boman. 2002. Influence of diet on conjugated linoleic acid content of milk, cheese and blood serum. J. Dairy Sci. 85 (Suppl. 1): 356 (Abstr. [ Links ]).
Lathamn, M. J., J. E. Stony, and M. E. Sharpe. 1972. Effect of low-roughage diets on the rumen and lipid metabolism in the rumen. Appl. Microbiol. 24 (6): 871-877. [ Links ]
Lock, A. L., and K. J. Shingfield. 2004. Optimizing milk composition. In: Dairying -Using Science to Meet Consumer's Needs. Kebreab, E., J. Mills, and D. E. Beever. (eds). Loughborough, UK. Occ. Pub. No. 29, Brit. Soc. Anim. Sci. Nottingham University Press. pp: 107-188. [ Links ]
Lock, A. L., and P. C. Garnsworthy. 2003. Seasonal variation in milk conjugated linoleic acid and delta (9)- desaturase activity in dairy cows. Livest. Prod. Sci. 79: 47-59. [ Links ]
Macoon, B., E. Sollenberger, E. Moore, R. Staples, H. Fike, and M. Portier. 2003. Comparison of three techniques for estimating the forage intake of lactating dairy cows on pasture. J. Anim. Sci. 81: 2357-2366. [ Links ]
Menke, K. H., and H. Steingass. 1988. Estimation of the energetic feed value from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 28: 7-55. [ Links ]
Morales-Almaráz, E., A. Soldado, A. González, A. Martínez-Fernández, I. Domínguez-Vara. B. de la Roza-Delgado, and F. Vicente. 2010. Improving the fatty acid profile of dairy cow milk by combining grazing with feeding on total mixed ration. J. Dairy Res. 77 (2): 225-230. [ Links ]
NRC. 2001. Nutrient Requirements of Dairy Cattle. 7th rev. The National Academy Press, Washington, DC. USA. 292 p. [ Links ]
Nielsen T. S., E. M. Straarup, M. Vestergaard, and K. Sejrsen. 2006. Effect of silage type and concentrate level on conjugated linoleic acids, trans-C18:1 isomers and fat content in milk from dairy cows. Repr. Nutr. Dev. 46: 699-712. [ Links ]
Palmquist, D. L., and T. C. Jenkins. 2003. Challenges with fats and fatty acid methods. J. Anim. Sci. 81: 3250-3254. [ Links ]
Peterson, D. G., J. A. Kelsey, and D. E. Bauman. 2002. Analysis of variation in cis9, trans11 conjugated linoleic acid (CLA) in milk fat of dairy cows. J. Dairy Sci. 85: 2164-2172. [ Links ]
Statistical Analysis System (SAS). 2002. Software (V.9.0). User's Guide. N. C., USA. 315 p. [ Links ]
Shingfield, K., C. Reynolds, B. Lupoli, V. Tolvonet, M. Yurawecz, P. Delmonte, J. Griinari, A. Grandison, and D. Beever. 2005. Effect of forage type and proportion of concentrate in the diet on milk fatty acid composition in cow given sunflower oil and fish oil. Anim Sci. 80: 225-238. [ Links ]
Steel, R. G. D., J. H. Torrie, and D. A. Dickey. 1997. Principles and Procedures of Statistics: A Biometrical Approach. 3rd ed. McGraw-Hill Series in Probability and Statistics. USA. 622 p. [ Links ]
Stockdale, C. R., G. P. Waler, W. J. Wales, D. E. Dalley, A. Birkett, Z. Shen, and P. T. Doley. 2003. Influence of pasture and concentrates in the diet of grazing dairy cows on the fatty acid composition of milk. J. Dairy Res. 70: 267-276. [ Links ]
Sukhija, P. S., and D. L. Palmquist. 1988. Rapid method for determination of total fatty acid content and composition of feedstuff's and feces. J. Agr. Food Chem. 36: 1202-1206. [ Links ]
Van Soest, P. J., J. B. Robertson, and B. A. Lewis. 1991. Methods of dietary, neutral detergent fiber and non starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74: 3583-3597. [ Links ]
Wijesundera, C., Z. Shen, W. J. Wales, and D. E. Dalley. 2003. Effect of cereal grain and fibers supplements on the fatty acid composition of milk fat of grazing dairy cows in early lactation. J. Dairy Res. 70: 257-265. [ Links ]
Williams, S. M. 2000. Dietary fatty acids and human health. Ann. Zootech. 49: 165-180. [ Links ]

