










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgrociencia
versão On-line ISSN 2521-9766versão impressa ISSN 1405-3195
Agrociencia vol.48 no.4 Texcoco Jun. 2014
Fitociencia
Líneas homocigóticas de maíz de alto rendimiento como progenitoras de híbridos de cruza simple
High yielding inbred lines of maize as progenitors of single cross hybrids
Francisco J. Pérez-López1, Ricardo Lobato-Ortiz1* , José de J. García-Zavala1, José D. Molina-Galán1, José de J. López-Reynoso2, Tarcicio Cervantes-Santana (†)
1 Postgrado en Recursos Genéticos y Productividad-Genética, Colegio de Postgraduados, Campus Montecillo. 56230. Montecillo, Texcoco, Estado de México. *Autor responsable
2 Departamento de Fitotecnia, Universidad Autónoma Chapingo, Km. 38.5 Carr. México-Texcoco. 56230. Chapingo, Estado de México. (rlobato@colpos.mx)
Recibido: enero, 2014.
Aprobado: mayo, 2014.
Resumen
En México, la producción de semilla de maíz (Zea mays L.) de híbridos de cruzas simples no es rentable debido a que las líneas usadas en ellos como progenitor femenino son de bajo rendimiento. El objetivo de este estudio fue evaluar la aptitud combinatoria general y específica de 10 líneas de maíz de alto rendimiento derivadas de poblaciones de maíces exóticos adaptados para emplearlas como progenitores de híbridos de cruzas simples. Las 45 cruzas dialélicas entre las 10 líneas, más los progenitores (Método II de Griffing) y cuatro híbridos comerciales usados como testigos, se evaluaron por tres años en Montecillo, Estado de México. Las líneas se derivaron de un sistema de siembra en altas densidades e irradiación gama durante ocho ciclos de selección. Las variables evaluadas fueron el rendimiento de mazorca por planta, sus componentes y los días a floración femenina. Las líneas con mayor y menor rendimiento per se rindieron 7.34 y 5.45 t ha-1, respectivamente. La Línea 3 presentó los efectos más altos de ACG, buen comportamiento per se y participó en tres de las cinco cruzas simples de mayor rendimiento. En cambio, la Línea 5 que estuvo presente en tres de las cinco cruzas de menor rendimiento, presentó menor rendimiento per se, baja ACG y baja ACE en combinaciones específicas. Las cruzas L3XL4 y L4XL5 igualaron el rendimiento de los mejores híbridos testigos, y fueron más precoces. Las líneas más productivas podrían representar una ventaja en la producción de semilla híbrida de cruzas simples con alto potencial productivo y de menor costo.
Palabras clave: Zea mays L., aptitud combinatoria, cruzas dialélicas, híbridos de cruza simple.
Abstract
In Mexico, maize (Zea mays L.) seed production of single cross hybrids is not profitable because the inbred lines used in these as the female parent are of low yield. The objective of this study was to assess the general and specific combining ability of 10 high yielding inbred lines of maize derived from adapted exotic maize populations to be used as parents for single cross hybrids. The 45 diallelic crosses between the 10 lines, the progenitors (Griffing's Method II), and four commercial hybrids used as controls, were assessed for three years in Montecillo, Estado de México. The lines were derived from a high-density cultivation system and gamma irradiation during eight cycles of selection. The traits evaluated were ear yield per plant, yield components, and days to female flowering. The lines with the greatest and lowest per se yield produced 7.34 and 5.45 t ha-1, respectively. Line 3 showed the highest GCA effects along with a good behavior per se and also participated in three of the five single crosses with the highest yield. In contrast, Line 5, which was present in three of the five crosses with the lowest yield, showed a low yield per se, low GCA and low SCA in specific combinations. The crosses L3XL4 and L4XL5 matched the yield of the best check hybrids and were more precocious. The most productive lines could represent an advantage for the production of seed from single cross hybrids of high productive potential and at lower costs.
Key words: Zea mays L., combining ability, diallelic crosses, single cross hybrids.
INTRODUCCIÓN
La producción comercial de híbridos de maíz (Zea mays L.) de cruzas simples se inició en EE.UU. en I960 porque ya existían líneas de alto rendimiento factibles de usarse como progenitores hembra (Crow, 1998). En México todavía no existen líneas altamente productivas, por lo cual se usan híbridos dobles o trilineales; además, el costo elevado de la semilla de híbridos de cruza simple ha limitado su producción comercial (Luna et al., 2012). Espinosa et al. (2002) afirman que la producción comercial de semilla de híbridos de maíz de cruzas simples es rentable cuando las líneas progenitoras hembra tienen un rendimiento mínimo de 3000 kg ha-1. Otros factores que limitan la producción de semilla de estos híbridos son: la condición homogénea de las líneas que ocasiona poco amortiguamiento ante los cambios en los ambientes de producción, la falta de mejoramiento en las poblaciones usadas como fuente de líneas y el riesgo de perder el control de los progenitores (Preciado y Terrón, 1996).
Para obtener líneas de maíz con alto potencial de rendimiento es necesario disponer de poblaciones base de tamaño grande, variables, y de alto rendimiento, y usar sistemas de endogamia moderado (Márquez, 1988) y, según Hallauer et al. (2010), su rendimiento puede aumentar mediante selección y mejoramiento de las líneas existentes. Cervantes et al. (2001) obtuvieron líneas con baja endogamia y alto rendimiento mediante irradiación gama y selección en altas densidades. Las líneas con bajos niveles de endogamia son vigorosas (Astete y Branco, 2002), por lo cual son más rendidoras y su uso como progenitoras de híbridos de cruza simple aumentaría la producción de semilla del progenitor hembra; los híbridos obtenidos tendrían un rendimiento igual o superior y una mayor homogeneidad genética, respecto a los híbridos trilineales y dobles.
La introducción de material exótico de maíz a los programas de mejoramiento permite aumentar la diversidad genética. Al identificar germoplasma exótico con alelos favorables carentes en las poblaciones locales, es posible transferir y aprovechar estos nuevos alelos en líneas de maíz y en los híbridos generados (Dudley, 1984). Las cruzas dialélicas permiten estimar los componentes de la variación genética, identificar las combinaciones superiores y seleccionar los mejores progenitores para el diseño de métodos de mejoramiento eficientes (Kempthorne y Curnow, 1961; Gutiérrez et al, 2002).
Sprague y Tatum (1942) definieron la aptitud combinatoria general (ACG) como el comportamiento promedio de una línea en sus combinaciones híbridas, y la aptitud combinatoria específica (ACE) como los casos en los cuales ciertas combinaciones híbridas específicas se expresan favorablemente o no con respecto al comportamiento promedio de sus progenitores. Griffing (1956) desarrolló cuatro métodos para evaluar ACG y ACE considerando: 1) los progenitores y sus cruzas F1 directas y recíprocas, 2) los progenitores y sus cruzas directas F1, 3) las cruzas F1 directas y recíprocas, 4) la evaluación de las cruzas F1 directas únicamente.
Mendes et al. (2003), Reyes et al. (2004) y Escorcia et al. (2010) señalan que una cruza simple es de alto rendimiento cuando las dos líneas progenitoras son de alta ACG, o bien, al menos una línea es de alta ACG, pero presenta efectos positivos de ACE. Las líneas con efectos altos de ACG pueden emplearse para desarrollar variedades sintéticas, mientras que combinaciones específicas de alto rendimiento se pueden usar en la hibridación (Sprague y Eberhart, 1977). Los efectos de ACG se relacionan con efectos aditivos que se pueden usar por selección y luego por hibridación, mientras que los efectos de ACE se deben a efectos de dominancia y epistasis (Inram et al., 2004) y son utilizados por la hibridación.
Con base en lo anterior, el objetivo de este estudio fue evaluar las aptitudes combinatorias general y específica de 10 líneas altamente homocigóticas y de alta capacidad de rendimiento, derivadas de poblaciones de maíces exóticos adaptados a clima templado, para emplearlas como progenitoras de híbridos de cruza simple. La hipótesis fue que los híbridos de cruza simple formados tendrán alta capacidad de rendimiento y características agronómicas similares o superiores a los híbridos comerciales de la zona.
MATERIALES Y MÉTODOS
En este estudio se evaluaron las 45 cruzas posibles entre 10 líneas de maíz altamente homocigóticas de alto potencial de rendimiento, derivadas de germoplasma exótico, las 10 líneas progenitoras y cuatro híbridos comerciales como testigos; es decir, se evaluaron 59 genotipos. Las líneas fueron generadas mediante la siembra de poblaciones de maíces tropicales adaptadas a valles altos en densidades de 200 000 plantas ha-1 y estas poblaciones se formaron previamente mediante cruzas intervarietales y por compuestos de razas tropicales de maíz. Antes de la floración, se seleccionaron y autofecundaron las plantas más vigorosas y precoces, bajo el supuesto de que éstas tenían una alta frecuencia de alelos favorables dominantes, hasta terminar el ciclo de cultivo con densidades de 80 000 plantas ha-1. A la cosecha se formaba un compuesto balanceado con igual número de semillas por mazorca de las plantas seleccionadas. Para obtener líneas altamente homocigóticas y con un alto coeficiente de endogamia (0.996), esta metodología se repitió por ocho ciclos; además se aplicó irradiación gama de Cobalto 60 entre cada ciclo de selección (Cruz et al., 2010). La genealogía de las líneas se encuentra en el Cuadro 1. Los testigos fueron los híbridos comerciales: HS-2, AMCCG01, AMCCG02 y PROMESA.

Los 59 genotipos se sembraron en los ciclos P-V 2009, 2010 y 2011 en Montecillo, Estado de México, de clima tipo Cw (i) B/2 (a') templado con lluvias en verano; precipitación media anual de 655.6 mm; temperatura media anual de 16.3 °C. Cada ciclo de cultivo se sembró en mayo y se aplicaron riegos de auxilio suficientes hasta el establecimiento de las lluvias. La fertilización fue 120 kg N y 80 kg P2O5 ha-1; todo el P2O5 y la mitad del N se aplicó a la siembra y el resto en la escarda (30 d después de la siembra, dds). La maleza se controló con el herbicida Gesaprim Calibre 90 GDA®, 3 L ha-1 aplicado, 15 dds.
El diseño experimental fue de bloques completos al azar y los tratamientos tuvieron cuatro repeticiones. La parcela experimental fue dos surcos, 6 m de largo, con 80 cm entre surcos y 40 cm entre plantas, y dos plantas por mata (62 500 plantas ha-1).
Catorce caracteres cuantitativos se midieron y con los datos de los tres años de evaluación se aplicó un análisis de correlación para identificar las cinco variables con mayor correlación positiva con el rendimiento de mazorca por planta. Estas variables fueron: número de mazorcas por planta (NMZ, número total de mazorcas por parcela experimental entre número de plantas); peso de mazorca (PMZ, g, peso seco por mazorca, promedio de cinco mazorcas seleccionadas al azar por parcela); peso seco de grano por mazorca (PGMZ, g, peso promedio de las cinco mazorcas); porcentaje de desgrane [(PDD = (PMZ/PGMZX100)]; y rendimiento de mazorca por planta (RMZP, g, peso de las mazorcas secas por parcela entre el número de plantas por parcela.
El análisis de varianza fue combinado para los tres años de evaluación para cada variable. La suma de cuadrados de cruzas se dividió en suma de cuadrados de ACG y suma de cuadrados de ACE. Además se estimaron los efectos de ACG (ĝi) de las 10 líneas y los efectos de ACE (ŝij) de las 45 cruzas posibles.
En el análisis genético se usó el método II del modelo del diseño dialélico de Griffing (1956):
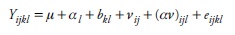
se consideró que:

donde Yijkl es el valor observado; µ es la media general; bkl es el efecto de bloque o repetición dentro de ambientes; αl es el efecto ambiental; Vij es el efecto de cruza; (αv)ijl es la interacción entre ambientes y cruzas; eijkl es el efecto residual o error; gi es el efecto de ACG para el progenitor i; gj es el efecto de ACG para el progenitor j; Sijes el efecto de ACE de la cruza ij; (αg)il es el efecto de interacción entre el efecto gi y el ambiente l; (αg)jl es el efecto de interacción entre el efecto gj y el ambiente l; (αs)ijl es el efecto de la interacción entre el efecto sij y el ambiente l.
La estructura genética para el rendimiento (RMZP) de las 45 cruzas se construyó mediante el modelo del diseño dialélico de Griffing (1956):

donde Yij es el rendimiento observado de la cruza ij en promedio de repeticiones y ambientes; û es la media general estimada; ĝi, ĝj y ŝij, estimadores de los efectos gi, gj y Sij, respectivamente.
Las medias en cada carácter se compararon con la prueba de Tukey (p≤0.05). Además se hizo una comparación especial del RMZP de los 10 mejores híbridos incluyendo a los testigos, en los que se agregó la variable días a floración femenina (DFF), en promedio de los tres ambientes.
RESULTADOS Y DISCUSIÓN
Análisis de varianza general
Los cuadrados medios del análisis de varianza combinado fueron significativos (p≤0.01) entre ambientes para todos los caracteres (Cuadro 2), lo cual significa que las condiciones y efectos ambientales fueron diferentes en los tres años de evaluación. Los efectos de ACG y ACE fueron significativos (p≤0.01) para todas las variables. En la suma de cuadrados (datos no presentados) de las cruzas, la variación correspondiente a efectos de ACG para las cinco variables fue: 24 % (RMZP), 30 % (NMZ), 31 % (PMZ), 31 % (PGMZ), y 15 % (PDD); en cambio en los efectos de ACE los resultados fueron: 76 % (RMZP), 70 % (NMZ), 69 % (PMZ), 69 % (PGMZ) y 85 % (PDD). Esto implica que las cinco variables evaluadas están controladas principalmente por efectos no aditivos. González et al. (1997) y de la Rosa et al. (2006) también encontraron mayor contribución de los efectos no aditivos en el rendimiento y sus componentes al evaluar cruzas dialélicas de maíz, mientras que Vasal et al. (1992) señalan que los efectos de ACE son mayores en cruzas de maíz cuyas líneas progenitoras son derivadas de distintas poblaciones y razas.
La interacción cruzas por ambiente fue altamente significativa para todas las variables (Cuadro 2). Las cruzas fueron evaluadas en la misma localidad durante distintos años, pero el comportamiento de algunas cruzas simples cambió drásticamente de un año a otro. Al respecto, Rowe y Andrew (1964) y Hallauer et al. (2010) mencionan que las cruzas simples presentan mayor interacción con el ambiente al ser poblaciones de condición más homogénea en comparación a los híbridos trilineales y dobles. La suma de cuadrados de la interacción cruzas por ambiente se dividió en ACG y ACE por ambientes, donde todas las variables mostraron diferencias significativas (p≤0.01), excepto PGMZ y PDD que presentaron diferencias significativas (p≤0.05) en la interacción ACG por ambientes. Los coeficientes de variación fluctuaron entre 2 y 15 %.
Comportamiento per se de las líneas
La Línea 2 mostró el promedio más alto para rendimiento de mazorca por planta respecto sólo a las líneas 10 y 7, donde la 7 tuvo el menor RMZP (Cuadro 3). La diferencia entre el promedio de RMZP de la línea 2 (117.6 g) con respecto a la 7 (87.2 g) es 25.8 %. Destaca que en la Línea 2 los promedios de PMZ, PGMZ y PDD no fueron diferentes (p>0.05) a los de la línea 7, y hubo diferencia significativa sólo en el número de mazorcas por parcela (línea 2 con 51; línea 7 con 46 mazorcas).
El rendimiento potencial de las líneas con base en una densidad de siembra de 62 500 plantas ha-1, indicó que las 10 líneas son altamente productivas, con 7.34 y 5.45 t ha-1 para la línea con mayor y menor rendimiento, respectivamente. Los rendimientos son superiores a 3000 kg ha-1, y esto es el mínimo que una línea de maíz debe producir para ser utilizable como progenitor femenino en híbridos de cruza simple (Espinosa et al., 2002).
Aptitud combinatoria general
Los mayores efectos positivos de ACG para rendimiento de mazorca por planta se observaron en las líneas 3 y 2, mientras que las líneas 7 y 5 presentaron los efectos negativos más bajos (Cuadro 4). Los efectos de aptitud combinatoria para NMZ, PMZ y PGMZ fueron positivos para las líneas 3 y 2; en cambio, en todos los caracteres fueron negativos para las líneas 7 y 5.
En general, los efectos de ACG de las líneas no mostraron valores positivos altos, lo cual podría deberse a que las líneas fueron seleccionadas por su comportamiento per se en campo y no con base en una prueba temprana de ACG. De acuerdo con los resultados, las líneas 3 y 2 podrían ser buenos progenitores para híbridos de cruza simple porque presentaron efectos positivos para ACG y mostraron buen comportamiento per se para rendimiento y sus componentes.
Aptitud combinatoria especifica (ACE)
De las 45 cruzas simples posibles sólo se muestran dos grupos de 10 cruzas (Cuadro 5): las cinco con los mayores y las cinco con los menores efectos de ACE para rendimiento de mazorca por planta. Para el primer grupo, los valores de ACE para los caracteres NMZ, PMZ y PGMZ fueron negativos (L3XL4) en los híbridos formados entre una línea de baja ACG con una línea de alta ACG, mientras que los valores de ACE para todos los caracteres fueron positivos (L8XL9) en los híbridos donde ambas líneas fueron de baja ACG. En el segundo grupo de cruzas con efectos de ACE negativos, se identificaron híbridos entre líneas de alta y baja ACE (L2XL6 y L3XL8), y también híbridos donde ambas líneas fueron de baja ACE (L7XL10, L5XL9, y L1XL5). Al respecto, De la Cruz et al. (2005) indican que en cruzas con altos y bajos efectos de ACE participan líneas con baja y alta ACG.
Estructura genética de las cruzas
Respecto a la estructura genética de las cinco cruzas simples con mayor y menor rendimiento de mazorca por planta, en las cruzas de mayor rendimiento participó al menos una línea con efectos positivos de alta ACG, además de que los efectos de ACE fueron positivos y altos, mientras que en las cruzas de menor rendimiento el efecto de ACG fue negativo en al menos una línea, y el efecto de ACE fue negativo en todos los casos (Cuadro 6). Estos resultados coinciden con los obtenidos por Mendes et al. (2003), Reyes et al. (2004) y Escorcia et al. (2010).
Las cruzas simples serán de alto rendimiento si sus dos líneas progenitoras son de alta ACG o si su efecto de ACE es alto y al menos uno de los progenitores es de alta ACG (Reyes, 2004), pero se busca que los efectos aditivos y no aditivos sean altos. En este estudio, en cuatro de las cinco cruzas simples de mayor rendimiento los efectos ACE fueron mayores a la suma de efectos de ACG de sus progenitores; así es posible que al combinar estas líneas de alto rendimiento con otro grupo de líneas de alta ACG el rendimiento de las cruzas simples pueda aumentar. La línea 3 estuvo presente en tres de las cinco cruzas simples con mayor rendimiento y presentó rendimiento alto per se, alta ACG y alta ACE en combinaciones específicas. En cambio, la línea 5 estuvo presente en tres de las cinco cruzas de menor rendimiento, pero tuvo rendimiento per se menor, ACG baja y ACE baja en combinaciones específicas.
Comparación de las cruzas simples con respecto a los testigos
En el Cuadro 7 se muestran los diez híbridos que tuvieron el mayor rendimiento de mazorca por planta, 2 testigos y 8 cruzas simples. Los híbridos comerciales HS-2 y AMCCG01 tuvieron los promedios más altos, pero dos cruzas simples (L3XL4 y L4XL5) fueron similares a los testigos (p>0.05). Los días a floración femenina tuvieron un valor promedio de 74 d para las ocho cruzas simples de mayor RMZP y fueron más precoces, pues los híbridos comerciales de mayor rendimiento presentaron valores para DFF entre los 79 y 83 d.

En el presente estudio se identificaron dos cruzas simples que estadísticamente fueron similares (p>0.05) a los híbridos comerciales de mayor rendimiento, y por lo menos ocho cruzas simples superaron a los híbridos testigos PROMESA (121.6 g) y AMCCG02 (127.1 g). En relación al número de días a floración femenina, las cruzas simples de mayor rendimiento en general fueron más precoces que los cuatro híbridos comerciales (Cuadro 7). La precocidad de las cruzas simples puede deberse a que para obtener las líneas se eliminaron las plantas más tardías y menos altas, las cuales no soportaron la competencia de las plantas precoces que crecieron más rápido y las sombrearon. Esto permitió que las plantas seleccionadas resultaran cuando menos de igual precocidad y altura que la variedad original, pero con mayor capacidad de rendimiento.
Las cruzas simples L3XL4 y L4XL5 podrían usarse como híbridos comerciales porque las líneas progenitoras de ambas cruzas mostraron buen rendimiento de semilla, y rendimientos similares al de los mejores híbridos testigos. Además, los híbridos HS-2 y AMCCG-01 son cruzas trilineales que requieren más de un ciclo agrícola y más trabajo para obtener y mantener su semilla; ésta es la ventaja de las cruzas simples de alto rendimiento identificadas en este estudio, porque podrían reducir el costo de la semilla híbrida para el productor.
CONCLUSIONES
El rendimiento de semilla de las 10 líneas en estudio fue superior a 3000 kg ha-1, por lo cual son económicamente rentables para utilizarlas como progenitoras hembras en híbridos de cruza simple. En las cinco cruzas simples de mayor rendimiento, al menos una línea progenitora mostró efectos positivos de ACG y efectos de ACE positivos y altos. La línea 3 se identificó como un progenitor potencial para híbridos de cruzas simple, porque presentó efectos positivos de ACG, efectos altos de ACE en combinaciones específicas y rendimiento per se bueno; además, esta línea participó en tres de las cinco cruzas simples con mayores rendimientos. Dos híbridos de cruza simple mostraron rendimientos similares a los mejores híbridos testigo, pero fueron más precoces. Los resultados no permiten aceptar la hipótesis planteada para todos los caracteres agronómicos, pero las cruzas simples más productivas tuvieron RMZP similar y menor número de días a floración femenina respecto a los testigos. Las líneas más productivas podrían tener una ventaja en la producción de semilla al generar híbridos de cruzas simples con un costo menor.
LITERATURA CITADA
Astete M., F., and J. Branco de M. 2002. Inbreding depression in maize populations of reduced size. Sci. Agric. 59: 335-340. [ Links ]
Cervantes S., T., M. Oropeza R., y D. Reyes L. 2001. Selección para rendimiento y heterosis de líneas endogámicas de maíz irradiado. Agrociencia 34: 421-431. [ Links ]
Crow J., F. 1998. 90 years ago: the beginning of hybrid maize. Genetics 148: 923-928. [ Links ]
Cruz I., S., A. Muratalla L., y A.T. Kato Y. 2010 (Comp). Cruzas Dialélicas de Líneas de Maíz Exóticas Adaptadas de Alto Rendimiento. La Investigación al Servicio del Campo Mexicano. Postgrado en Recursos Genéticos y Productividad-Genética. Colegio de Postgraduados, Campus Montecillo. Montecillo, Texcoco, Estado de México. pp: 10-12. [ Links ]
De la Cruz-Lázaro, E., S. Rodríguez-Herrera, M. Estrada-Botello, J. Mendoza-Palacio, y N. Brito-Manzano. 2005. Análisis dialélico de líneas de maíz QPM para características forrajeras. Universidad y Ciencia 21: 19-26. [ Links ]
De la Rosa L., A., H. De León C., F. Rincón S., y G. Martínez Z. 2006. Efectos genéticos, heterosis y diversidad genética entre híbridos comerciales de maíz adaptados al Bajío Mexicano. Rev. Fitotec. Mex. 29: 247-254. [ Links ]
Dudley J., W. 1984. A method for identifying populations containing favorable alleles not present in elite germplasm. Crop Sci. 24: 1053-1054. [ Links ]
Escorcia G., N., J. Molina G., F. Castillo G., y J. Mejía C. 2010. Rendimiento, heterosis y depresión endogámica de cruzas simples de maíz. Rev. Fitotec. Mex. 33: 271-279. [ Links ]
Espinosa C., A., M. Sierra M., y N. Gómez M. 2002. Producción y tecnología de semillas mejoradas de maíz por el INIFAP en el escenario sin la PRONASE. Agron. Mesoam. 14: 117-121. [ Links ]
González S., H. Córdova, S. Rodríguez, H. De León, y V. Serrato. 1997. Determinación de un patrón heterótico a partir de la evaluación de un dialélico de diez líneas de maíz Subtropical. Agron. Mesoam. 8: 01-07. [ Links ]
Griffing, B. 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Austr. J. Biol. Sci. 9: 463-493. [ Links ]
Gutiérrez D., E., A. Palomo G., A. Espinoza B., y E. De la Cruz L. 2002. Aptitud combinatoria y heterósis para rendimiento de líneas de maíz en la Comarca Lagunera, México. Rev. Fitotec. Mex. 25: 271-277. [ Links ]
Hallauer A., R., M. Carena, and J. B. Miranda F. 2010. Quantitative Genetics in Maize Breeding. Springer-Verlag New York Inc. 664 p. [ Links ]
Inram M., S., H. Nawaz M., H. Mahmood M., and M. Munir. 2004. General and specific combining ability studies in maize diallel crosses. Int. J. Agric. Biol. 6: 856-859. [ Links ]
Kempthorne O., and R. Curnow N. 1961. The partial diallel cross. Biometrics 17: 229-250. [ Links ]
Luna M., A. Hinojosa R., O. Ayala G., F. Castillo G., y J. Mejía C. 2012. Perspectivas de desarrollo de la industria semillera de maíz en México. Rev. Fitotec. Mex. 35: 1-7. [ Links ]
Márquez S., F. 1988. Genotecnia Vegetal. Tomo II (métodos, teoría y resultados). AGT EDITOR, S.A. México. pp: 472-473. [ Links ]
Mendes A., A., L. Carlini-Garcia., A. Resende S., M. Figueireido S., A. Franco G., and C. Lopes S. 2003. Combining ability of inbred lines of maize and stability of their respective single-crosses. Sci. Agric. 60: 83-89. [ Links ]
Preciado O., R., y A. Terrón I. 1996. Efectos heteróticos entre y dentro de fuentes de germoplasma de maíz a través de cruzas simples emparentadas. Agric. Tec. Mex. 22: 219-230. [ Links ]
Reyes L., D., J. Molina G., M. Oropeza R., y E. Moreno P. 2004. Cruzas dialélicas entre líneas autofecundadas de maíz derivadas de la raza Tuxpeño. Rev. Fitotec. Mex. 27: 49-56. [ Links ]
Rowe, P., R., and R. Andrew. 1964. Phenotipic stability for a systematic series of corn genotypes. Crop Sci. 4: 563-567. [ Links ]
Sprague G., F., and S. Eberhart A. 1977. Corn and Corn Improvement. American Society of Agronomy. Madison, Wisconsin. pp: 305-363. [ Links ]
Sprague G., F., and A. Tatum L. 1942. General vs. specific combining ability in single crosses of corn. J. Am. Soc. Agron. 34: 923-32. [ Links ]
Vasal S., R., G. Sirinivasan, and C. Han G. 1992. Heterotic patterns of eightyeight white subtropical CIMMYT maize lines. Maydica 37: 319-327. [ Links ]

