










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgrociencia
versão On-line ISSN 2521-9766versão impressa ISSN 1405-3195
Agrociencia vol.47 no.7 Texcoco Out./Nov. 2013
Fitociencia
Study of heterosis in Bangladeshi Chilli (Capsicum annuum L.) Landraces
Estudio de la heterosis en variedades nativas de chile (Capsicum annuum L.) de Bangladesh
M. Hasanuzzaman1, M. A. Hakim2,3*, M. M. Hanafi2, Abdul Shukor-Juraimi4, M. M. Islam5, A. K. M. Shamsuddin6
1 Department of Genetics and Plant Breeding.
2 Department of Land Management. *Author for correspondence. (ahakimupm@gmail.com).
3 Department of Agricultural Chemistry, HSTU, Dinajpur 5200, Bangladesh.
4 Department of Crop Science.
5 Department of Forest Management, Universiti Putra Malaysia, 43400 UPM Serdang Selangor, Malaysia.
6 Department of Genetics and Plant Breeding, BAU, Mymensingh, Bangladesh.
Received: September, 2012.
Approved: August, 2013.
Abstract
Chilli is an important cash crop in Bangladesh but average yield is very low (0.89 t ha-1) and genetic potentiality of Bangladeshi chilli landraces for hybrid variety development has not been evaluated. The objective of this study was to find out heterotic behavior following Gardner and Eberhart model (1966) II. Six different homozygous divergent parents CCA 2, CCA 5, BARI Morich 1, CCA 11, CCA 15 and CCA 19 were used to estimate heterosis. A significant amount of heterosis was present in yield and yield contributing traits. Estimate of variety heterosis for yield per plant was significantly positive in CCA 5 and BARI Morich 1. In BARI Morich 1, the significant and positive variety heterosis for yield per plant was associated with significant and positive estimates of heterosis for number of fruits per plant and number of seeds per fruit, suggesting that these yield traits contributed to the final heterosis manifested through yield. Indigenous x exotic crosses showed significant amount of heterosis. It is possible to emphasize indigenous X exotic crosses for good fruit yield, particularly to be used as commercial hybrids. Hybrids of BARI Morich 1 x CCA 19 and CCA 5 x BARI Morich 1 showed better performance.
Keywords: Capsicum annuum L., indigenous X exotic cross, yield.
Resumen
El chile es un cultivo comercial importante en Bangladesh pero su rendimiento promedio es muy bajo (0.89 t ha 1) y la potencialidad genética de las variedades nativas de chile de Bangladesh para desarrollar variedades híbridas no se ha evaluado. El objetivo de este estudio fue conocer el comportamiento heterótico siguiendo el modelo de Gardner y Eberhart (1966) II. Seis diferentes padres divergentes homocigóticos CCA 2, CCA 5, BARI Morich 1, CCA 11, CCA 15 y CCA 19 se usaron para estimar la heterosis. Hubo una cantidad significativa de heterosis en el rendimiento y las características que contribuyeron al rendimiento. La estimación de la heterosis varietal para rendimiento por planta fue significativamente positiva en CCA 5 y BARI Morich 1. En BARI Morich 1, la heterosis varietal significativa y positiva para el rendimiento por planta se asoció con las estimaciones significativas y positivas de heterosis para el número de frutos por planta y el número de semillas por fruto, sugiriendo que estos rasgos de rendimiento contribuyeron a la heterosis final manifiesta a través del rendimiento. Las cruzas nativas X exóticas mostraron una cantidad significativa de heterosis. Se puede enfatizar las cruzas nativas X exóticas para un buen rendimiento del fruto, particularmente si se usan como híbridos comerciales. Los híbridos de BARI Morich 1XCCA 19 and CCA 5 X BARI Morich 1 mostraron un mejor rendimiento.
Palabras clave: Capsicum annuum L., cruza nativo X exótico, rendimiento.
INTRODUCTION
Heterosis is an important avenue for increasing yield and other economic traits in capsicum peppers (Joshi, 1986). Chilli (Capsicum annuum L.) is a diploid (2n = 24) species and genetically self-pollinated and chasmogamous crop whose flowers open only after pollination (Lemma, 1998), and 2 to 96 % out-crossing was observed under open pollination (Tanksley, 1984; Pickersgill, 1997; AVRDC, 2000). Cross-pollination is caused by bees, by thrips and ants to a limited scale and rarely by wind (AVRDC, 2000).
Landrace varieties as parents show high heterosis and the fruit quality is higher than those of the landrace parents; besides, they show a rather good shape, smooth skin and large fruits (Milerue et al., 2000). Expression of heterosis in F1 hybrids of Capsicum species depends upon parents, which may be selected based on plant vigor, fruit size and yield (Greenleaf, 1947). According to Melchinger (1999), starting with genetically divergent populations as heterotic groups has advantage of maximum exploitation of heterosis and hybrid performance. Higher efficiency of inter-population improvement leads to superior hybrids by using divergent rather than genetically similar heterotic groups (Melchinger and Gumber 1998; Reif et al., 2007). Genetic diversity between parents, recognized as a requirement for maximal expression of heterosis, has probably arisen through diversity in origin (geographical separation), in ancestral relationships, in gene frequencies or in morphological traits (Virmani, 1996).
The chilli landraces of Bangladesh are heterogeneous, serve as a reservoir of genetic variability for plant breeder and have survived hundreds of years through human and natural selection. Landrace crops are grown for long periods of time within a given farming system, have high capacity to tolerate local biotic and abiotic stresses and show high yield stability and intermediate yield levels under low-input agricultural systems (Zeven, 1998). Chilli landraces are selected by farmers due to their important agronomic and horticultural traits (heat level, fruit size and color, early maturity) and as a result of natural selection, are well adapted to the specific environment. Landraces of Bangladesh are grown in different confined areas and genetically distinct from each other. The local chilli landraces are City morich, Bindu, Balijuri, Diapara, Jagri, Kolabari, Abhaguz, Bogra local, Hathazari local, Halda, Comilla local, Chittagong, Naga, Bain, Kantai, Mota-morich, Lomba-morich, Ausa marich, Akashi, Matal, Kala morich, Ausadhebra, Shamali, Shikharpuri, Dhani morich and Surjamukhi (Khaleque, 1992; Rashid, 1999), which are cultivated throughout the year.
Average green chilli yield was 1.69 t ha-1 during 2010-2011 (BBS, 2011), which is lower than the 1.59 t ha-1 (dried chilli) obtained in India during 20112012 (MoA, 2012). Developing hybrid varieties with desirable qualities could reverse the low productivity trend, but heterosis in chilli has not been reported yet in Bangladesh. Therefore, the objective of this study was to estimate heterosis among six parents of chilli (Capsicum annuum L.).
MATERIALS AND METHODS
Experimental site and soil characteristics
This experiment was carried out at the Research and Development Farm of Lal Teer Seed Limited (23.9763° N and 090.3539° E), Gazipur, Bangladesh, from October 2007 to July 2009. The soil was clay loam, slightly acidic (soil pH 6.0 to 6.5) and low in nitrogen content.
Plant materials
Chilli genotypes CCA 2 (P1), CCA 5 (P2), CCA 15 (P5) and CCA 19 (P6) were Bangladeshi landraces, BARI Morich 1(P3) released by BARI, Bangladesh (center of origin Sri Lanka) and CCA 11 (P4) (collected from Thailand), were crossed in a diallel mating system excluding the reciprocals in winter season 20072008. The six genotypes were supplied by Lal Teer Seed Limited, Bangladesh. In the next winter season (2008-2009), all the six parents and their 15 F1's were studied.
Seed sowing, cultivation and crossing
In 2007-2008, the parental materials were sown on September 26, 2007 in seedling trays and transplanted into the field on November 3, 2007. Crossing was done in a 6X6 diallel design excluding reciprocals between December 2007 and April 2008. The mature F1 fruits were harvested and dried and seeds were threshed and stored in a gene bank. In 2008-2009, the seeds of six parents and their 15 F1's were sown in seedling trays on September 10, 2008. Seedlings age of 4 to 5 leaves were transplanted to the field on October 16, 2008. Raised beds 1.5 m width were prepared for transplanting; plant to plant distance was 50 cm, row to row distance 70 cm and bed to bed distance 1.0 m which was used as drain. The experimental design was randomized complete block with three replications and the experimental unit was 3.5 mX1.5 m.
Data analysis
Components of heterosis in a diallel cross were estimated according to the Gardner and Eberhart (1966) model II. The analysis was performed on individual environments using the DIALLEL-SAS05 program (Zhang et al., 2005). The linear model (Zhang et al., 2005) was:
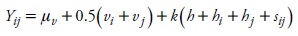
where Yij=mean of the cross between the ith and jth parental populations, μv=mean of all parental populations, vi=varietal effects of parental population I, vj= varietal effects of parental population j, h= average heterosis contributed by all parental populations used in crosses, hi= average contributions of individual parental population i to the expression of heterosis, hj= average contributions of individual parental population j to the expression of heterosis, sij= specific heterosis (specific combining ability, SCA) that occurs when parental population i is crossed to parental population j, k= 0 when i= j and k= 1 when i≠j.
RESULTS AND DISCUSSION
Analysis of variance due to heterosis
The variances due to heterosis were significant for days to 50 % flowering, fruit length, fruit weight, number of seeds per fruit, number of fruits per plant and yield per plant (Table 1). These results suggested the presence of non-additive gene effects for these characters. Variation due to heterosis (overall heterosis) was partitioned into average, variety and specific heterosis.
Significant average heterosis (parents vs. F1) of days to 50 % flowering, fruit length, fruit weight, number of fruits per plant, and yield per plant, showed heterosis. Variety heterosis was estimated to judge overall contribution of a variety to its array heterosis. The variance due to variety heterosis was significant for days to 50 % flowering, fruit length, fruit weight, number of seeds per fruit, number of fruits per plant and yield per plant. This result indicated the differences among the parental arrays for the heterosis of these characters (Table 1).
Importance of heterosis among crosses was assessed by variances due to specific heterosis which were significant for days to 50 % flowering, fruit length, fruit weight, number of seeds per fruit, number of fruits per plant and yield per plant. The results related to variance of different components of heterosis indicated that each significantly contributed to the total heterosis of the crosses. Souza and Maluf (2003) observed significant mean heterosis, varietal heterosis and specific heterosis components obtained from the Gardner-Eberhart (1966) analysis in number of seeds per fruit; however, mean heterosis was not significant in total fruit yield in Capsicum chinense. The varietal heterosis component of total fruit yield was not significant either, indicating that there were no significant differences among hybrid arrays of the different parental lines. The significance of the specific heterosis component in yield indicated that there are specific hybrid combinations with a high degree of heterosis. Therefore, in spite of the significance of the parental effects, the per se parental performance is not a good indicator of the hybrid total yield. The sum of the squares of all heterosis components contributed significantly to the treatment sum of the squares, showing the importance of non-additive genetic effects (dominance or epistasis) for the expression of number of seeds per fruit.
Variety heterosis of the parents
Among the six parents, only CCA 5 and BARI Morich 1 had significant positive estimates of yield per plant (Table 2) and significant positive estimates of heterosis for number of fruits per plant was expressed in BARI Morich 1, suggesting that this yield contributing trait contributed to the final heterosis observed through yield. Fruit weight played a significant role in parent CCA 5 by contributing significant variety heterosis in yield per plant. A significant positive estimate of number of seeds per fruits and plant height also was observed in BARI Morich 1. Souza and Maluf (2003) reported a significants positive and negative variety heterosis in seeds number per fruit, which is similar to the present result, but they found no parent significance for total fruit yield.
Specific heterosis of the crosses
Crosses P2XP3 and P3XP6 showed significant heterosis in fruit yield per plant and yield per plant, but in cross P3XP6 only number of seeds per fruit was significant (Table 3). These results are consistent with those obtained by Souza and Maluf (2003) who reported that 7 out 10 crosses were significant for specific heterosis in total fruit yield.
Top two yields per plant were obtained in cross P3XP6 (898.87g) and P2XP3 (833.63 g) (Table 4). In these two crosses parent BARI Morich 1 (Parent 3) was common, CCA 5 (Parent 2) and CCA 19 (Parent 6) were indigenous (Bangladesh) and BARI Morich 1 (Parent 3) was exotic (Sri Lanka). Indigenous X exotic crosses showed significant heterosis and they could be used for good fruit yield, as commercial hybrids, as well as for developing hybrid varieties. In hybrids among Asian lines there is high heterosis, indicating genetic diversity and potential for improving hot pepper using genotypes from different regions, along with elite inbred lines from local cultivars (Marame et al., 2009).
The high heterosis observed in this study suggests the usefulness of using genetically diverse germplasm in a breeding program. This is in agreement with the observations of Gill et al. (1973) and Shifriss and Sacks (1980) who reported that genetic divergence of the parents in Capsicum species is highly related to the heterosis of the F1. Very closely or distantly related parents showed low heterosis, but crosses between parents of intermediate divergence classes tended to show higher heterosis effect for fruit yield, fruit length and fruit weight (Geleta et al., 2004) and large heterosis.
CONCLUSIONS
Bangladeshi land races have genetic potential for developing hybrid. This potentiality can be explored by crossing it with exotic Asian genotypes.
ACKNOWLEDGEMENTS
We are thankful to the Ministry of Higher Education Malaysia for financial support through the Research University Grant Scheme to Universiti Putra Malaysia.
LITERATURE CITED
AVRDC (Asian Vegetable Research and Development Centre). 2000. Multiplying seed of pepper lines. International cooperators' guide. Asian Vegetable Research and Development Center, Taiwan. pp: 1. [ Links ]
BBS (Bangladesh Bureau of Statistics). 2011. 2011 Year Book of Agricultural Statistics of Bangladesh. Bangladesh Bureau of Statistics, Ministry of Planning, GOB. Dhaka, Bangladesh. 38 p. [ Links ]
Gardner, C. O., and S. A. Eberhart. 1966. Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22: 439-452. [ Links ]
Geleta, L. F., M. T. Labuschagne, and C. D, Viljoen. 2004. Relationship between heterosis and genetic distance based on morphological traits and AFLP markers in pepper. Plant Breed. 123: 467-473. [ Links ]
Gill, H. S., P. S. Thakur, and T. C. Thakur .1973. Combining ability in sweet pepper (Capsicum annuum L.) var. Grossum Scudt. Indian J. Agric. Sci. 43: 918-921. [ Links ]
Greenleaf, W. H. 1947. Line breeding as a method of improving the pimento peppers. Proc. Am. Soc. Hortic. Sci. 49: 224. [ Links ]
Joshi, S. 1986. Results of heterosis breeding on sweet pepper (Capsicum annuum L.). Capsicum Eggplant Newsletter 6: 49-50. [ Links ]
Khaleque, M. A. 1992. Principal component analysis in determining the taxonomic status of chili in Bangladesh. Bangladesh J. Bot. 21(2): 181-193. [ Links ]
Lemma, D. 1998. Seed production guideline for tomatoes, onion and hot pepper. IAR, Addis Abeba. pp: 11-27. [ Links ]
Marame, F., L. Dessalegne, C. Fininsa, and R. Sigvald. 2009. Heterosis and heritability in crosses among Asian and Ethiopian parents of hot pepper genotypes. Euphytica 168: 235-247. [ Links ]
Melchinger, A. E. 1999. Genetic diversity and heterosis. In: Coors, J. G., and S. Pandey (eds). The Genetics and Exploitation of Heterosis in Crops. Proc. Int. Symp. CIMMYT, Mexico City, Mexico, 17-22 August 1997. Crop Science Society of America, Madison, Wisconsin, USA. pp: 99-118. [ Links ]
Melchinger, A. E., and R. K. Gumber.1998. Overview of heterosis and heterotic groups in agronomic crops. In: Lamkey, K. R., and J. E. Staub (eds). Concepts and Breeding of Heterosis in Crop Plants. CSSA Special Publication No. 25. Crop Science Society of America, Madison, WI. pp: 29-44. [ Links ]
Milerue, N., M, Nikornpun, M. Nogluck, and N. Maneechat. 2000. Studies on heterosis of chilli (Capsicum annuum L.). Kasetsart J. Natural Sci. 34(2): 190-196. [ Links ]
MoA (Ministry of Agriculture). 2012. Agricultural Statistics at a Glance 2012. Directorate of Economics and Statistics, Department of Agriculture and Cooperation, Ministry of Agriculture, Government of India, New Delhi. 176 p. [ Links ]
Pickersgill, B. 1997. Genetic resources and breeding of Capsicum spp. Euphytica 96: 129-133. [ Links ]
Rashid, M. M. 1999. Shabjee Bigghan. Rashid Publishing House, 94, old DOHS, Dhaka-1206. pp: 201-205. [ Links ]
Reif, J. C., F. M. Gumpert, S. Fischer, and A. E. Melchinger. 2007. Impact of inter population divergence on additive and dominance variance in hybrid populations. Genetics 176: 1931-1934. [ Links ]
Shifriss, C., and J. M. Sacks.1980. The effect of distance between parents on the yield of sweet pepperXhot pepper hybrids, Capsicum annuum in a single harvest. Theor. Appl. Genet. 58(6): 253-256. [ Links ]
Souza, J. A. D., and W. R. Maluf. 2003. Diallel analyses and estimation of genetic parameters of hot pepper (Capsicum chinense Jacq.). Scientia Agricola 60(1): 105-113. [ Links ]
Tanksley, S. D. 1984. High rates of cross-pollination in chile pepper. Hort Sci. 19: 580-582. [ Links ]
Virmani, S. S. 1996. Hybrid in rice. Adv Agron. 57: 377-462. [ Links ]
Zeven, A. C. 1998. Landraces: a review of definitions and classifications. Euphytica 104(2): 127-139. [ Links ]
Zhang, Y., M. S. Kang, K. R. Lamkey. 2005. DIALLEL-SAS05: A comprehensive program for Griffing's and Gardner Eberhart analyses. Agron. J. 97: 1097-1106. [ Links ]

