










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.47 no.2 Texcoco feb./mar. 2013
Recursos naturales renovables
Almacenes de carbono en bosques montanos de niebla de la Sierra Norte de Oaxaca, México
Carbon storage in montane cloud forests in Sierra Norte of Oaxaca, México
Gustavo Álvarez–Arteaga1*, Norma E. García Calderón2, Pavel Krasilnikov3, Felipe García–Oliva4
1 Facultad de Ciencias. Universidad Nacional Autónoma de México. México D. F. *Autor responsable: galvareza68@gmail.com.
2 UMDI, Facultad de Ciencias. Campus UNAM–Juriquilla. Querétaro, México.
3 Institute of Biology, Karelian Research Centre RAS 185610, Petrozavodsk, Russia.
4 Centro de Investigaciones en Ecosistemas, Universidad Nacional Autónoma de México, Campus Morelia, Michoacán, México.
Recibido: abril, 2012.
Aprobado: enero, 2013.
Resumen
Una de las principales incertidumbres sobre la función de los bosques tropicales y subtropicales dentro del ciclo global del carbono proviene de la escasa información sobre la capacidad de estos ecosistemas para acumular carbono (C). En el caso de los bosques montanos de niebla, la amplia heterogeneidad ambiental a la que están sujetos determina cambios importantes sobre su composición estructural y florística, que pudieran incidir sobre los contenidos de C almacenado en sus diferentes componentes. El objetivo de este estudio fue evaluar el contenido de C en los almacenes aéreos y subterráneos para cinco comunidades de bosque montano de niebla en la Sierra Norte de Oaxaca, situadas entre1500 y 2500 m de altitud. El diseño fue de muestreo jerárquico anidado y las variables fueron C en la biomasa arbórea, estrato herbáceo y arbustivo, material leñoso muerto, mantillo, raíces finas y suelo mineral para cada comunidad vegetal. La suma de los distintos almacenes indicó que el C acumulado fue mayor a 1500 y 2500 m (440 y 420 Mg ha–1), y entre 1950 y 2400 m fue 354 Mg ha–1. En promedio, la reserva de carbono en biomasa y suelo para los bosques montanos de niebla fue 384.16±40.13 Mg ha–1. La conformación de estos almacenes está regulada por las variaciones biofísicas dentro del intervalo altitudinal, traducidas en cambios florísticos y estructurales de la vegetación, en la producción y preservación de residuos orgánicos poco transformados en la superficie del suelo, así como por la capacidad de éste último para formar asociaciones órgano minerales que estabilicen el carbono orgánico en el suelo.
Palabras clave: gradiente altitudinal, carbono aéreo, carbono subterráneo, condiciones físicas y bióticas.
Abstract
One of the principal uncertainties regarding the function of the tropical and subtropical forests within the global carbon cycle is due to the lack of information of the capacity of these ecosystems to accumulate carbon (C). In the case of the montane cloud forests, the wide environmental heterogeneity to which they are subjected determines important changes in their structural and floristic composition, which could affect the C contents stored in their different components. The objective of this study was to evaluate the C content in the above and below ground storage for five communities of montane cloud forest in the Sierra Norte of Oaxaca, situated between 1500 and 2500 m altitude. The design was hierarchic nested sampling and the variables were C in the arboreal biomass, herbaceous and shrub stratum, dead woody material, humus, fine roots and mineral soil for each plant community. The sum of the different stores indicated that the accumulated C was higher than 1500 and 2500 m (440 and 420 Mg ha–1), and between 1950 and 2400 m it was 354 Mg ha–1. On the average, the carbon reserve in biomass and soil for the montane cloud forests was 384.16±40.13 Mg ha–1. The conformation of these stores is regulated by the biophysical variations within the altitudinal interval, translated in floristic and structural changes of the vegetation, in the production and preservation of scantly transformed organic residue on the soil surface, as well as its capacity to form organo mineral associations that estabilize organic carbon in the soil.
Key words: altitudinal gradient, aboveground carbon, underground carbon, physical and biotic conditions.
INTRODUCCIÓN
Pese a que los bosques montanos de niebla (BMN) representan sólo 0.26 % de la superficie terrestre (Bubb et al., 2004), la participación de estos ecosistemas dentro del ciclo global del carbono (C) es relevante debido a su creciente deterioro, el cual implica la emisión de cantidades importantes de dióxido de carbono (CO2) a la atmósfera. Para entender la magnitud de este fenómeno es necesario cuantificar los almacenes de C terrestre en estos sistemas de la manera más precisa y completa posible, partiendo del conocimiento de su historia y de la gama de sus interacciones bióticas y abióticas; luego, con base en su estudio, debatir su capacidad para proveer servicios ambientales como la captura de C.
Los BMN tienen capacidad para almacenar cantidades altas de C en la biomasa y en el suelo mineral, comparados con otros ecosistemas forestales (Kitayama y Aiba, 2002). En la literatura revisada se encontraron pocos estudios acerca del C almacenado en estos ecosistemas en México (de Jong et al., 1999; Acosta, 2002 y Rodríguez et al, 2006). Los datos obtenidos confirman que el BMN está dentro de los ecosistemas de mayor acumulación de C por unidad de superficie en el país. Pero hay poca investigación acerca de la influencia que ejerce la heterogeneidad ambiental (distribución de especies, cambios en temperatura, humedad y propiedades edáficas) sobre la acumulación de C en los diferentes almacenes y que se magnifica dentro de estos sistemas a través del amplio intervalo altitudinal en el cual se establecen. Considerando esta influencia se propone que los cambios físicos y bióticos afectan la capacidad de acumular C en los diferentes componentes del sistema. Por tanto, el objetivo de este estudio fue evaluar los almacenes de C en la biomasa y el suelo mineral de cinco comunidades conservadas de BMN, a lo largo de un gradiente altitudinal y determinar su posible relación con algunas variables físicas y bióticas.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se realizó en los bosques de la comunidad indígena de Santa Cruz Tepetotutla, (Figura 1) Sierra Norte de Oaxaca, México (17° 38'–17° 40' N y 96° 32'–96° 33' O), caracterizada por su relieve montañoso abrupto, humedad relativa alta y una gran biodiversidad (Martin, 1993). Los bosques se ubican entre 1500 y 2500 m, dentro de la ladera norte del río Perfume, tributario de la cuenca del río Papaloapan. El material geológico está compuesto por esquistos de mica y clorita con inclusiones menores de cuarcita (Centeno–García, 2004). El clima de la zona es C (m)(w) templado húmedo con lluvias abundantes en verano (García, 2004). La precipitación media anual a una altitud de 1450 m es 5800 mm y la temperatura media anual es 16.5 °C. (Rzedowski y Palacios–Chávez, 1977).
Los suelos dominantes son Podzoles fólico estágnicos y Stagnosoles fólicos entre 2050 y 2500 m; 2050 y 1500 m son Cambisoles fólicos. Las características comunes a estos suelos son acidez extrema, fuerte lavado de bases y acumulación importante de restos orgánicos en la superficie del suelo (Álvarez et al., 2008).
Selección de sitios
Se estudiaron cinco comunidades vegetales conservadas de BMN ubicadas a 1500, 1940, 2050, 2400 y 2500 msnm. Las comunidades entre 1500 y 2050 m son bosques montanos bajos (Holdridge, 1967) y tienen gran afinidad florística con los bosques premontanos húmedos subyacentes; desde 2100 m existe una marcada transición hacia bosques montanos altos con presencia importante de líquenes y epifitas, y en su parte más alta por especies de mayor afinidad con los bosques templados, como las del género Quercus (Meave et al., 2006).
Métodos de muestreo
Para medir el C almacenado en biomasa y suelo en las diferentes comunidades, se usó un diseño de muestreo jerárquico anidado (Hughes et al., 1999; Jaramillo et al., 2003) y se delimitaron 4 subparcelas de 750 m2 dentro de una parcela mayor de 3000 m2 por comunidad vegetal. En cada subparcela se midió el diámetro a la altura del pecho (DAP, 1.3 m) para todos los árboles vivos y muertos con DAP > 10 cm. Los árboles del subdosel con altura mayor a 1.3 m y DAP < 10cm, así como el material leñoso muerto sobre el suelo (MLMS) se midieron en 5 parcelas de 25×4 m distribuidas de manera concéntrica al interior de la parcela mayor. Para el MLMS, se midió diámetro y longitud y se tomaron muestras de los diferentes grados de descomposición para determinar la densidad de la madera en laboratorio. La biomasa de los árboles se calculó mediante ecuaciones alométricas desarrolladas para bosques montanos de la Sierra Norte de Oaxaca (Acosta et al., 2002). La biomasa de arbustos y herbáceas se cuantificó dentro de cada parcela de 25×4 m en dos cuadros de 1x1 m (n=10 por comunidad vegetal). Dentro de las mismas parcelas se marcaron 4 cuadros de 0.5x0.5 m (n=20 por comunidad vegetal) y se recolectó el mantillo. El material cosechado fue pesado y registrado, seleccionando una muestra para determinar su masa seca en estufa a 80 °C y la concentración de C.
Determinación de C en suelo mineral y raíces finas
Para medir el almacén de carbono en el suelo mineral y en las raíces finas (<5 mm de diámetro), se perforaron 2 perfiles de 25x4 m (n=10 por comunidad veetal) hasta una profundidad de 100 cm. Las muestras de suelo se tomaron por horizonte con tres repeticiones extraídas con un cilindro de 100 cm3 de capacidad para evaluar densidad aparente y raíces finas. Se recolectó una muestra adicional para cuantificar el C en laboratorio. Se consideraron las raíces finas hasta una profundidad de 30 cm, fueron extraídas manualmente, secadas a 80 °C y pesadas para calcular su biomasa. El almacén total de raíces (finas, medias y gruesas) se calculó con la relación biomasa raíces / biomasa arbórea del 19 % (de Jong et al., 1999).
Los almacenes de biomasa arbórea (Mg ha–1) se transformaron a almacenes de C aplicando el factor de conversión de 0.5 (IPCC, 2001). Para mantillo y raíces finas se tomaron muestras directas en los diferentes sitios y se determinó la concentración del C orgánico total con un analizador de carbono (UIC. Mod. CM5012), la cual se aplicó como factor de conversión para obtener los respectivos almacenes de C. La concentración de C en las muestras del suelo mineral (g C kg–1 suelo) fue determinada por combustión en húmedo (van Reeuwijk, 2002).
El almacén de biomasa total se obtuvo de la suma de sus componentes: biomasa arbórea (BA), herbáceo–arbustivas, material leñoso y raíces. El almacén de C en suelo mineral se calculó con la concentración promedio de C en cada horizonte, su densidad aparente y profundidad. También se calculó la media, desviación estándar y coeficiente de variación por almacén de C, expresado en Mg C ha– 1) para cada comunidad vegetal estudiada.
RESULTADOS Y DISCUSIÓN
Almacenes de carbono en biomasa
El contenido de biomasa arbórea denotó un patrón complejo con respecto a la altitud: el valor máximo se registró a 2500 m y representó casi el doble (1.8 veces) de lo encontrado en 1950 y 2400 m y 1.2 veces el valor a 1500 m de altitud (Cuadro 1 y Figura 2). Estas diferencias se relacionan con la estructura y composición florística de las comunidades vegetales descritas por Meave et al. (2006) y por Álvarez (2010), que apoyan lo descrito por Brown y Lugo (1982) y Clark y Clark (1996), quienes indican que la presencia de árboles con DAP > 70 cm en bosques tropicales y subtropicales incrementa considerablemente los contenidos de biomasa para estos bosques. Esto se manifestó notablemente en la comunidad vegetal situada a 2500 m, donde este segmento (DAP > 70 cm) representó casi 70 % de la biomasa arbórea, en tanto que para los sitios restantes sólo representó entre 9 y 33 %.
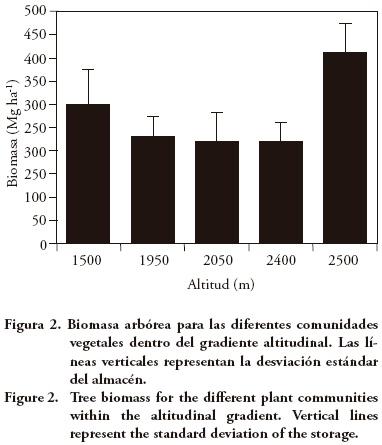
La biomasa del mantillo (4.6 a 6.13 Mg ha–1) no tuvo variaciones significativas dentro del intervalo altitudinal, condición similar a la reportada por Kitayama y Aiba (2002) y Edwards y Grubb (1977), para bosques montanos y apoyan a Brown y Lugo (1982) quienes indican que los contenidos de mantillo son estables para diferentes tipos de bosques en zonas tropicales y subtropicales. La proporción del MLMS con respecto a la biomasa aérea total fue mayor en la comunidad situada a 1500 m con 62.6 Mg ha–1 (17 %) que en el resto del intervalo (8 %). Los valores registrados para este segmento son elevados, pero justificados si se considera que la práctica de extracción de material leñoso no es recurrente e incluso está prohibida por las autoridades locales y, por tanto, todo este material queda sujeto a los procesos de descomposición natural que implicarían largos periodos de tiempo bajo las condiciones ambientales imperantes.
La proporción más alta de biomasa en árboles muertos se encontró a 1950 m (21.72 Mg ha–1) con 8 % de la biomasa aérea total y en conjunto la participación del material leñoso (material leñoso sobre el suelo + árboles muertos en pie) fue 6 a 18 %, valores parecidos a los encontrados por Delaney et al. (1997) para bosques montanos bajos de Venezuela.
El contenido de biomasa en raíces finas en los primeros 30 cm de profundidad fue mayor para las comunidades situadas a 1950 y 2400 m (entre 12 y 19 Mg ha–1) y menor para las de 1500 y 2500 m (4.2 Mg ha–1). Estos contenidos son superiores a los reportados por Kitayama y Aiba (2002) (5.2 a 14.4 Mg ha–1). Las diferencias descritas se justifican de acuerdo con Grubb (1977) y Bruijnzeel y Veneklaas (1998) en la necesidad de mantener un sistema radical superficial muy desarrollado debido a las condiciones adversas de toma de nutrimentos presentes en la mayoría de los suelos en que se desarrollan estos bosques. En el presente caso las restricciones son evidentes y se reflejan en la acidez extrema, fuerte compactación del horizonte de eluviación y los rasgos de hidromorfismo detectados por Álvarez et al. (2008), que se magnifican para este segmento del gradiente altitudinal. Por tanto, es notable que entre 1950 y 2500 m la distribución de raíces finas y gruesas queda prácticamente limitada a la capa orgánica superficial. Esta condición es menos evidente a 1500 m altitud en la que prácticamente desaparecen estas limitantes y se observa un desarrollo radical a mayor profundidad.
Almacenes de carbono en suelo
Los almacenes de C en la fracción mineral del suelo se mantuvieron constantes entre 1500 y 2000 m (220 ), pero desde 2400 m declinaron hasta 158 Mg C ha– 1 (Cuadro 2) a los 2500 m. Esta variación se entiende por dos condiciones dentro del intervalo: 1) el espesor de la capa orgánica superficial se mantuvo casi constante hasta 2400 m, pero disminuyó drásticamente a partir de 2500 m; 2) los horizontes minerales del suelo acumularon menores contenidos de C conforme aumentó la altitud (Cuadro 2).

Comportamiento de los almacenes de C dentro del intervalo de altitud
Grubb (1977), Bruijnzeel y Veneklaas (1998) y Bubb et al. (2004) reconocen la influencia de la niebla persistente sobre la estructura y morfología de las comunidades vegetales, como es la disminución en diámetro y altura de los árboles, pero está poco documentado el efecto sobre los almacenes de C en biomasa y suelo. En este estudio se constató que la capacidad de estas comunidades forestales para almacenar C en sus diferentes componentes varía dentro del intervalo de altitud (Cuadro 2) y queda definida por tres condiciones generales (Figura 3): 1) a 1500 m donde la comunidad forestal corresponde a un bosque montano bajo en el que los dos principales almacenes de C (biomasa y suelo) son elevados y se encuentran en proporciones similares; 2) entre 1950 y 2400 m donde coincide con la mayor condensación de nubes y hay una reducción notable en el diámetro y altura del arbolado, lo cual implica una disminución directa en el almacén de C en biomasa, pero el C en el suelo tiende a registrar contenidos altos debido a la persistencia de una espesa capa orgánica superficial, y esta condición permite inferir la prevalencia de tasas de mineralización menores y la preservación de materia orgánica poco descompuesta; 3) a 2500 m, altitud que coincide con el parteaguas de la cuenca y en la cual la nubosidad se dispersa.

Dentro de esta última zona se registra la menor densidad de árboles pero su DAP y altura alcanza valores más altos, lo cual les confiere mayor capacidad para almacenar C en este segmento de la biomasa viva. El almacén de C en la fracción mineral del suelo se reduce debido a la disminución en el espesor de la capa orgánica superficial y también por la menor capacidad de este componente para formar complejos estables con la materia orgánica del suelo debido a la reducida proporción de arcillas dentro del complejo edáfico (Álvarez et al., 2008).
En el presente estudio se propone que los cambios en los patrones de acumulación de C en los diferentes almacenes pueden estar correlacionados con los cambios biofísicos registrados dentro del intervalo altitudinal, como su composición florística y estructural (Meave et al. 2006; Álvarez, 2010), procesos pedogenéticos (Álvarez et al., 2008) y climáticos (Velázquez Rosas et al., 2003).
Considerando el promedio de valores para el almacén total del bosque montano de niebla, los 384 ±45 obtenidos en este estudio (Cuadro 2) es inferior al reportado por de Jong (1999) que fue 468 para bosques montanos de Chiapas, pero similar al registrada por Acosta et al. (2001) para bosques de liquidámbar en la región mazateca de Oaxaca. La presente determinación es congruente con estudios que ubican a estos ecosistemas entre 250 a 783 (Álvarez, 2010), lo cual muestra la extrema variabilidad de estos bosques para almacenar carbono, pero también a las diferencias metodológicas de cada estudio.
CONCLUSIONES
Los almacenes de carbono en biomasa aérea y suelo en comunidades de Bosque Montano de Niebla, en la Sierra Norte de Oaxaca, son importantes reservorios de carbono regional. Así, su conservación es prioritaria en las políticas y planes de ordenamiento territorial estatal y federal.
Las variaciones en los reservorios de carbono, tanto en biomasa como suelo, están supeditadas a los cambios en la estructura y composición florística de las comunidades vegetales, así como en los procesos pedogenéticos imperantes dentro del gradiente altitudinal, para la zona de estudio. Estas variaciones biofísicas fungen como variables integradoras de la heterogeneidad ambiental propia de este tipo de ecosistemas forestales.
AGRADECIMIENTOS
Este proyecto fue auspiciado por el Consejo Nacional de la Ciencia y Tecnología (CONACyT), proyecto PAPIIT–UNAM (IN216906–3) y el Posgrado en Ciencias Biológicas–UNAM. Agradecemos también al M. en C. Abel Ibáñez Huerta y a la M. en C. Elizabeth Fuentes Romero por su apoyo en los trabajos de campo y laboratorio, a Rodrigo Velázquez–Durán por los análisis de C en las muestras de mantillo y raíces finas y al Biólogo Jesús Ricardo de Santiago Gómez por realizar la recolección e identificación de las especies arbóreas.
LITERATURA CITADA
Acosta, M., M., J. J. Vargas H., A. Velázquez M., y J. D. Etchevers B. 2002. Estimación de la biomasa aérea mediante el uso de relaciones alométricas en seis especies arbóreas en Oaxaca, México. Agrociencia 36: 725–736. [ Links ]
Álvarez A., G., N. E. García–Calderón, P. V. Krasilnikov, S.N. Sedov, V.O. Targulian and N. Velázquez–Rosas. 2008. Soil altitudinal sequence on base–poor parent material in a montane cloud forest in Sierra Juárez, Southern México. Geoderma 44: 593–612. [ Links ]
Álvarez A., G. 2010. Evaluación de la captura y estabilidad del carbon en un ecosistema de bosque mesófilo de montaña a lo largo del gradiente altitudinal en la Sierra Norte de Oaxaca. (Tesis de doctorado). Facultad de Ciencias. UNAM. 127 p. [ Links ]
Brown, S., and A. F. Lugo. 1982. The storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica 14: 161–187. [ Links ]
Bruijnzeel, L. A., and E. J. Veneklaas. 1998. Climatic conditions and tropical montane forest productivity: the fog has not lifted yet. Ecology 79:3–9. [ Links ]
Bubb, P., I. May, L. Miles and J. Sayer. 2004. Cloud forest agenda report. UNEP WCMC, Cambridge. UK. 36 p. [ Links ]
Centeno–García, E. 2004. Configuración geológica del estado. In: García–Mendoza A., J., M. de J. Ordóñez y M. Briones–Salas (eds). Biodiversidad de Oaxaca. UNAM–FOCN-WWF, México. D.F. pp: 29–42. [ Links ]
Clark, D. B. and D. A. Clark. 1996. Abundance, growth and mortality of very large trees in neotropical lowland rain forests. For. Ecol. Manage. 80: 235–244. [ Links ]
de Jong, B. H., M. A. Cairns, P. Haggerty, N. Ramírez–Marcial, S. Ochoa–Gaona, J. Mendoza–Vega, M. González–Espinosa, and I. March–Mifsut. 1999. Land–use change and carbon flux between 1970s and 1990s in Central Highlands of Chiapas, México. Environ. Manage. 23: 373–385 [ Links ]
Delaney, M., S. Brown, A. E. Lugo, A. Torres–Lezama, and N. Bello Quintero. 1997. The quantity and turnover of dead wood in permanent forest plots in six life zones of Venezuela. Biotropica 30:2–11. [ Links ]
Edwards, P. J., and P. J. Grubb. 1977. Studies of mineral cycling in a montane rain forest in New Guinea. The distribution of organic matter in the vegetation and soil. J. Ecol. 65: 943–969. [ Links ]
García, E. 2004. Modificaciones al Sistema de Clasificación Climática de Köppen. Serie de Libros No. 6. Quinta Edición. IG, UNAM. México. D.F. 105 p. [ Links ]
Grubb, P. J. 1977. Control of forest growth and distribution on wet tropical mountains: with special reference to mineral nutrition. Ann. Rev. Ecol. and Systematics 8: 83–107. [ Links ]
Holdridge, L. 1967. Life Zone Ecology. Tropical Science Center, San José, Costa Rica. 206 p. [ Links ]
Hughes, R., J. B. Kauffman and V. J. Jaramillo. 1999. Biomass, carbon, and nutrient dynamics of secondary forests in a humid region of México. Ecology 80:1892–1907. [ Links ]
IPCC. 2001. Climate change 2001: The scientific basis. Intergubernamental Panel for Climatic Change. Summary for policymakers. Cambridge University Press. United Kingdom. 20 p. [ Links ]
Jaramillo, V. J., J. B. Kauffman, L. Rentería–Rodríguez, D. L. Cummings, and L. J. Ellingson. 2003. Biomass, carbon, and nitrogen pools in Mexican tropical dry forest landscapes. Ecosystems 6:609–629. [ Links ]
Kitayama, K., and S. I. Aiba. 2002. Ecosystem structure and productivity of tropical rain forests along altitudinal gradients with contrasting soil phosphorus pool in Mount Kinabalu, Borneo. J. Ecol. 90: 37–51. [ Links ]
Martin, G. J. 1993. Ecological classification among the chinantec and mixe of Oaxaca, México. Etnoecológica 1:17–33. [ Links ]
Meave A., J., A. Rincón, and M. A. Romero–Romero. 2006. Oak forests of the hyper–humid region of La Chinantla, Northern Oaxaca range, México. In: Kappelle, M. (ed). Ecology and Conservation of Neotropical Montane Oak Forests. Ecological Studies 185. Springer–Verlag. Berlin, Heidelberg. pp: 113–125. [ Links ]
Rodríguez Laguna R., J., Jiménez Pérez, O. A. Aguirre, y E. J. Treviño Garza. 2006. Estimación del carbono almacenado en un bosque de niebla en Tamaulipas, México. UANL. Ciencias. 9: 179–186. [ Links ]
Rzedowski, J., y R. Palacios Chávez. 1977. El bosque de Engelhardtia (Oreomunnea) Mexicana en la region de la Chinantla (Oaxaca, México), una reliquia del Cenozoico. Bol. Soc. Bot. Méx. 36: 93–127. [ Links ]
Swift, M. J., O. W. Anderson, J. M. Anderson, D. J. G. Smith, P. Piteka, and A. Frank. 1979. Decomposition in Terrestrial Ecosystems. University of California Press. Berkeley. USA. 372 p. [ Links ]
Velázquez–Rosas N., J. Meave, and S. Vázquez–Santana. 2002. Elevational variation of leaf traits in montane rain forest tree species at La Chinantla, southern México. Biotropica 34: 534–546. [ Links ]
van Reeuwijk, L. P. (ed). 2002. Procedures for Soil Analysis. 6th edition. ISRIC–FAO. ISRIC Technical Paper No. 9. [ Links ]

