










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgrociencia
versão On-line ISSN 2521-9766versão impressa ISSN 1405-3195
Agrociencia vol.47 no.2 Texcoco Fev./Mar. 2013
Fitociencia
Eficiencia en el uso de la radiación, tasa de asimilación neta e integral térmica en función del fósforo en maíz (Zea mays L.)
Radiation–use efficiency, net assimilation rate and thermal integral as a function of phosphorus in maize (Zea mays L.)
Ernesto Díaz–López1*, Juan M. Loeza–Corte1, Jesús M. Campos–Pastelín1, Edgar J. Morales–Rosales , Aurelio Domínguez–López , Omar Franco–Mora
1 Ingeniería en Agroindustrias, Universidad de la Cañada. UNCA. Carretera Teotitlán–San Antonio Nanahuatipam Km. 1.7. 68540. paraje Titlacuatitla, Teotitlán de Flores Magón, Oaxaca. México. *Autor responsable: (lernesto@colpos.mx).
2 Programa de Ciencias Agropecuarias y Recursos Naturales (PCARN), Universidad Autónoma del Estado de México. UAEM. Instituto Literario # 100, Col. Centro. 50000. Toluca, Estado de México, México.
Recibido: mayo, 2012.
Aprobado: enero, 2013.
Resumen
El fósforo es un macroelemento esencial pata las plantas y junto con factores ambientales, como la radiación y la temperatura, influye en el crecimiento y rendimiento de los cultivos como el maíz (Zea mays L.). El objetivo del presente estudio fue comparar el efecto de seis niveles de fósforo (0, 40, 80, 120, 160 y 200 kg de P2O5 ha–1) en la eficiencia en el uso de la radiación, la tasa de asimilación neta y las unidades calor de los cultivares de maíz Amarillo Almoloya, Cacahuacintle y Cóndor, en condiciones de punta de riego, en el Cerrillo Piedras Blancas, Estado de México. El diseño experimental fue de bloques completos al azar con arreglo factorial; los 18 tratamientos resultaron de las combinaciones de los tres cultivares y los seis niveles de fósforo. Los datos se analizaron con un ANDEVA y las medias de los tratamientos con la prueba DSH (p<0.05). El análisis de los resultados indicó que Cacahuacintle respondió mejor al ambiente de estudio con 80 kg de P2O5 ha–1, ya que tuvo mayor rendimiento (7136.7 kg ha–1), eficiencia en el uso de la radiación (EUR; 0.33 MJ m–2) y tasa de asimilación neta (TAN; 0.023 g m–2 día–1) que los otros cultivares. Se concluyó que el fósforo afecta EUR, el crecimiento y el rendimiento de maíz con dosis bajas (0 y 40 kg ha–1) y altas (160 y 200 kg ha–1).
Palabras clave: cultivares, polinización libre, rendimiento agronómico, unidades calor, Zea mays.
Abstract
Phosphorus is an essential macroelement for plants and along with environmental factors, such as radiation and temperature influences the growth and yield of crops such as maize (Zea mays L.). The aim of this study was to compare the effect of six levels of phosphorus (0, 40, 80, 120, 160 and 200 kg P2O5 ha–1 on radiation–use efficiency, net assimilation rate and heat units of the maize cultivars Amarillo Almoloya, Cacahuacintle and Cóndor, in tip irrigation conditions in the Cerrillo Piedras Blancas, Estado de Mexico. The experiment was a randomized complete block design with a factorial arrangement; the 18 treatments were combinations of three cultivars and six levels of phosphorus. Data were analyzed with an ANOVA and treatment means with the HSD test (p<0.05). Analysis of results indicated that Cacahuacintle responded better to the study environment with 80 kg P2O5 ha–1, since it had higher yield (7136.7 kg ha–1), radiation–use efficiency (RUE; 0.33 MJ m–2) and net assimilation rate (NAR, 0.023 g m–2 day–1) than the other cultivars. It was concluded that phosphorous affect RUE, corn growth and yield with low (0 and 40 kg ha–1) and high doses (160 and 200 kg ha–1).
Key words: cultivars, open pollination, agronomic yield, heat units, Zea mays.
INTRODUCCIÓN
La radiación solar, como factor ambiental, es importante para los seres vivos porque es la fuente de energía para procesos físicos y biológicos, de ella dependen los vegetales para elaborar carbohidratos mediante la fotosíntesis (Taiz y Zeiger, 2002). La radiación, al interactuar con otros factores como la temperatura y precipitación, influye en el crecimiento y rendimiento de los cultivos (Rivetti, 2007). Así, la agricultura es una forma de cosechar la energía solar cuando se complementa con un adecuado suministro de agua, nutrimentos y dióxido de carbono (Castillo et al., 2007). Los efectos de la radiación solar sobre las plantas se clasifican en: 1) fotoenergéticos, que involucran la fotosíntesis y 2) fotoestimulantes, relacionados con el crecimiento y los movimientos násticos (Salisbury y Ross, 1994). La eficiencia en el uso de la radiación (EUR) se usa para estudiar el efecto de la radiación incidente en los cultivos (De la Casa et al., 2011). Así, Giménez et al. (1994) cuantificaron valores mayores de EUR con densidades altas de plantas (5.7 m–2) que con densidades bajas (2.9 m–2). La densidad de población alta de soya (Glicine max L.) intercepta mayor cantidad de radiación, incrementa la biomasa (1400 g m–2) y afecta el índice de área foliar (IAF) con valor de 4.0 (Mercau et al., 2001). El nitrógeno también influye en el desarrollo vegetativo y modifica EUR en girasol (Helianthus annuus (L) RB).
La tasa de asimilación neta (TAN) indica la cantidad de biomasa acumulada en función del área foliar y el tiempo (Morales et al., 2006; Morales et al., 2009). Al respecto, Pearcy et al. (1994) señalaron que TAN es una medida de la eficiencia de las hojas para producir nuevo crecimiento vegetal y tiene mayor importancia que otras variables fisiológicas, como la tasa de crecimiento relativo (TCR). Así, Flores et al. (2009) mostraron que tubérculos de papa (Solanum tuberosum L.) para semilla alcanzaron un valor máximo de TAN (10 g m–2 d–1), a pesar de ser una especie con metabolismo C3, cuando crecieron con 100 % de la radiación durante todo el ciclo de cultivo, y disminuyó en las plantas que crecieron con 40 y 70 % de radiación incidente.
El efecto de la temperatura en el desarrollo de los cultivos se evalúa con la integral térmica o con los grados día de desarrollo (Díaz et al., 2012). Esta variable relaciona la temperatura ambiental con las etapas fenológicas del cultivo y permite pronosticar cuándo ocurrirá una etapa fenológica e incluso calcula el rendimiento de un cultivo (Kantolic y Slafer, 2001).
El objetivo de este estudio fue evaluar el efecto de seis niveles de fósforo en tres cultivares de maíz (Zea mays L.), la integral térmica sobre la EUR, la TAN y el rendimiento de maíz. La hipótesis fue que los niveles de fósforo afectan la EUR, la TAN y el rendimiento de maíz.
MATERIALES Y MÉTODOS
El estudio se realizó en condiciones de campo, en el Cerrillo Piedras Blancas, Estado de México, (19° 24' N, 99° 54' O; 2600 msnm). Se aplicó un riego hasta alcanzar la capacidad de campo antes del surcado. El suelo fue tipo vertisol en proceso de formación, con pH 6.6 a 0.30 m de profundidad, 2.6 % de materia orgánica, 5.8 mg P kg–1 (determinado con el método propuesto por Bray y Kurtz de acuerdo con Díaz et al., 2011), capacidad de intercambio catiónico (CIC) de 0.5 Cmol(+) kg–1 de suelo secado al aire, conductividad eléctrica (CE) 0.03 dSm–1, y 5.1 mg Ca kg–1 y 15 mg Mg kg–1 intercambiables. El clima de la región es Cw1eg, que corresponde a clima templado, con lluvias en verano y precipitación de 800 a 1250 mm, temperatura media anual de 12 °C (de 7 a 14 °C) y mayo el mes más cálido antes del solsticio de verano (García, 2005).
El diseño experimental fue bloques completos al azar con arreglo factorial de dos factores: 1) los cultivares Amarillo Almoloya, Cacahuacintle (ambos de polinización libre) y Cóndor (híbrido), cultivados con densidad de 7.5 plantas m–2; 2) el nivel de fertilización fosfatada (superfosfato de calcio triple) 0, 40, 80, 120, 160 y 200 kg ha–1, aplicado con la siembra. Además, todo el cultivo fue fertilizado con 190 kg ha–1 de N (urea, 46 % N) y 60 kg ha– 1 de K2O (60 % K). La unidad experimental tuvo cinco surcos de 4 m de largo, con separación de 0.80 m y la parcela útil tuvo los tres surcos centrales.
Hubo cuatro muestreos destructivos a los 34, 63, 95 y 120 d después de la siembra (dds) (Arenas et al., 2011; Díaz et al., 2011) y se midió biomasa seca (BS) del vástago. Se separaron las hojas, el tallo, las brácteas e inflorescencias masculina y femenina, se secaron 72 h a 70 °C hasta alcanzar su peso constante, en una estufa con aire forzado, y se pesaron en una balanza analítica. El rendimiento de grano (R) se evaluó en madurez fisiológica (humedad de 13 %) y se obtuvo el promedio de 10 mazorcas (g planta–1).
La radiación interceptada (RI) se midió a los 34, 63, 95 y 120 dds, con un sensor lineal modelo (Li—Cor, 191) que mide RI en porcentaje y la radiación fotosintéticamente activa entre 400–700 nm. EUR se calculó con la ecuación 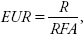 donde R es el rendimiento y RFA es la radiación fotosintéticamente activa, determinada con los datos de radiación global (cal cm–2 d–1) (RG), obtenidos de la estación meteorológica automatizada de la Facultad de Ciencias Agrícolas de la Universidad Autónoma del Estado de México. Los datos de RG se multiplicaron por la constante 0.50, para obtener los valores de RFA, éstos, a su vez, se multiplicaron por 0.04148 para transformarlos a MJ m–2 d–1 (Soil Science Society of America, 2000). TAN se calculó con la fórmula
donde R es el rendimiento y RFA es la radiación fotosintéticamente activa, determinada con los datos de radiación global (cal cm–2 d–1) (RG), obtenidos de la estación meteorológica automatizada de la Facultad de Ciencias Agrícolas de la Universidad Autónoma del Estado de México. Los datos de RG se multiplicaron por la constante 0.50, para obtener los valores de RFA, éstos, a su vez, se multiplicaron por 0.04148 para transformarlos a MJ m–2 d–1 (Soil Science Society of America, 2000). TAN se calculó con la fórmula  donde PS1 y PS2 son los pesos de las estructuras vegetales en los tiempos T1 y T2 de cada muestreo, AF1 y AF2 son las áreas foliares, y lnAF1 y lnAF2 son los logaritmos neperianos del área foliar. Durante el ciclo del cultivo se registraron la temperatura máxima (Tmáx) y mínima (Tmín) diarias para estimar la integral térmica del cultivo con el método residual de Díaz et al. (2012), con la ecuación
donde PS1 y PS2 son los pesos de las estructuras vegetales en los tiempos T1 y T2 de cada muestreo, AF1 y AF2 son las áreas foliares, y lnAF1 y lnAF2 son los logaritmos neperianos del área foliar. Durante el ciclo del cultivo se registraron la temperatura máxima (Tmáx) y mínima (Tmín) diarias para estimar la integral térmica del cultivo con el método residual de Díaz et al. (2012), con la ecuación  donde TT es el tiempo térmico o integral térmica, Tmáx es la temperatura máxima, Tmín es la temperatura mínima diaria, TB es la temperatura base del cultivo (7 °C para maíz) (García y López, 2002).
donde TT es el tiempo térmico o integral térmica, Tmáx es la temperatura máxima, Tmín es la temperatura mínima diaria, TB es la temperatura base del cultivo (7 °C para maíz) (García y López, 2002).
Los datos fueron analizados mediante ANDEVA, con el procedimiento GLM de SAS (2002) y las medias de los tratamientos se compararon con la prueba DSH (p<0.05).
RESULTADOS Y DISCUSIÓN
La temperatura máxima osciló entre 23 y 27 °C y la mínima entre 5 y 10 °C; RG durante el ciclo de cultivo fue 3654.7 MJ, osciló entre 14 y 20 MJ, y la máxima se observó durante la primer decena de mayo; la RG menor se presentó en la primer decena de noviembre (Figura 1).
Hubo diferencias significativas entre los cultivares y las dosis de fósforo (Cuadro 1) y la interacción fósforo × variedad fue significativa. Cacahuacintle presentó la R mayor (6459.6 kg ha–1) y la de Amarillo Almoloya (5390.8 kg ha–1) no fue estadísticamente diferente (p>0.05) a la de Cóndor (5597 kg ha–1). La mejor respuesta se observó con 80 kg de P2O5 ha–1, pues R fue 7136.7 kg ha–1, y la peor fue con 200 kg de P2O5 ha–1 y con el testigo (R, 5139.4 y 4686.7 kg ha–1). Esta diferencia puede deberse al gran porte arqueotípico y a la respuesta mayor de Cacahuacintle para extraer fósforo. González et al. (2006) atribuyeron una respuesta similar a la gran variabilidad genética y adaptación del cultivar a las condiciones ecológicas del valle de Toluca, pues con 80 kg de P2O5 ha–1 su R fue superior a 7000 kg ha–1.
Amarillo Almoloya, al interaccionar con 80, 120 y 160 kg de P2O5 ha–1, tuvo R máximas (5370, 6261 y 6386 kg grano ha–1); la R mayor la presentó Cacahuacintle al interactuar con 80 kg P2O5 ha–1; y el híbrido Cóndor con 80 y 160 kg ha–1 (6845 y 5903 kg de grano ha–1) (Cuadro 2). Estos resultados coinciden con los obtenidos por González et al. (2007) con maíz Cacahuacintle (5290 kg grano ha–1) y 80 kg de P2O5 ha–1 en El Cerrillo Piedras Blancas, Estado de México, y contrastan con los del maíz palomero Toluqueño (4200 kg grano ha–1), que es un maíz amarillo. Los autores atribuyeron estas respuestas al porcentaje mayor de variación entre los genotipos en los días a floración femenina, acame, pudrición de mazorca y rendimiento de grano.

Hubo diferencias significativas en el efecto de los niveles de fósforo aplicados y las variedades generaron diferencias significativas en la biomasa producida, y la interacción fósforo × variedad fue significativa (Cuadro 3). Cacahuacintle presentó valores de biomasa más elevados que Amarillo y Cóndor; es decir, aquél fue el más eficiente en la conversión de la energía solar a biomasa.
Amarillo Almoloya mostró EUR (0.31 y 0.32 g m–2 MJ–1) mayor que Cacahuacintle y Cóndor al interactuar con las dos dosis altas de P2O5; en contraste, estas dos variedades alcanzaron EUR máximas (0.48 y 0.32 g m–2 MJ–1) con 80 kg de P2O5 ha–1 (Cuadro 4). Este comportamiento correspondió con la R mayor en Cacahuacintle y Cóndor con 80 kg fósforo ha–1. Estos valores de EUR difieren de los de Andrade et al. (1993) de 1.52 a 1.53 g m–2 MJ –1 también con maíz. Las diferencias pueden deberse a los efectos del sitio donde se realizaron los estudios y a los genotipos utilizados.
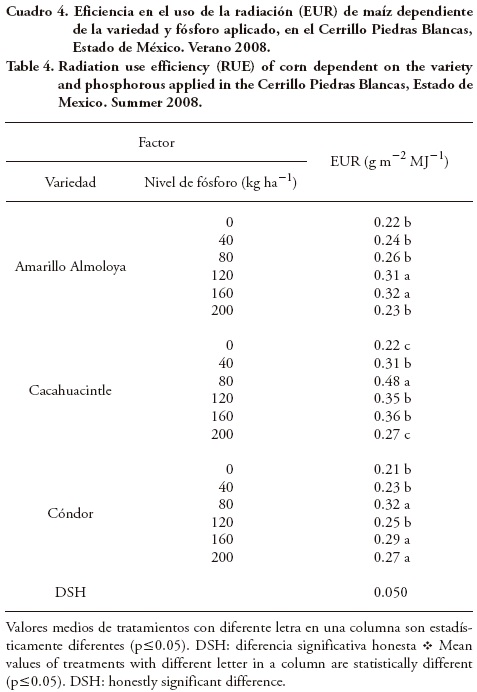
Existieron diferencias significativas en TAN entre las variedades en los cuatro muestreos realizados; el fósforo generó diferencias sólo 120 dds y la interacción fue significativas 34 dds. Destacó que el CV fue bajo (entre 16 y 22 %) a los 34, 63 y 95 dds, pero 120 dds el CV tuvo el valor mayor (33.8) (Cuadro 5).
La TAN presentó diferencias significativas dependientes del fósforo únicamente 24 y 120 dds, y las diferencias entre las variedades fueron significativas en los cinco muestreos. Además, la interacción fue significativa a los 34 dds. TAN disminuyó en las tres variedades a lo largo de los muéstreos (Cuadro 6). Así los valores mayores correspondieron con los 34 dds, y de ellos el mayor (0.023 g m–2 d–1) fue el de Cacahuacintle; pero después fue superado por Amarillo Almoloya y Cóndor, a los 63 y 95 dds.
Las diferencias de TAN debidas al fósforo se observaron 34 y 120 dds, con 200 kg ha–1 y las TAN menores las presentó el testigo 34 y 120 dds (Cuadro 7). Lo anterior indica que los niveles altos de fósforo (200 kg ha–1) incrementaron la expansión foliar y el IAF, con lo que se incrementó la intercepción de radiación (Aguilar et al., 2006); además, incrementó la elaboración de fotosintatos, aumentó la materia seca y se afectó la TAN. La interacción fósforo × cultivar indicó que los valores mayores de TAN (0.032, 0.025 y 0.020 g cm–2 d–1) correspondieron a Cacahuacintle con 80, 160 y 200 kg P2O5 ha–1 (Figura 2) y el valor menor correspondió al testigo de Cóndor (0.0033 g cm–2 d–1). Así, el incremento de TAN parece deberse a la respuesta de los híbridos a la fertilización fosfórica, pues los niveles de fósforo inferiores a 60 kg ha–1 mantienen el rendimiento bajo y, en consecuencia, la ganancia de biomasa seca (Aceves et al., 2002).
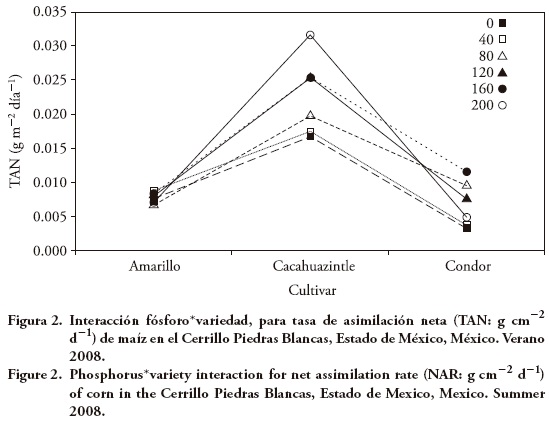
La integral térmica, o unidades calor se ajustó al modelo lineal UC = 6.17 dds + 53.2 (Figura 3), el tiempo térmico total fue 1158.6 UC, 77.03 de estas se acumularon entre la siembra a la emergencia (Ve), 677.08 entre la emergencia y la floración masculina (Vt), 18.49 entre la floración masculina y la floración femenina (R1), 59.81 entre la floración femenina y el estado de ampolla (R2), 49.4 de este estado al de grano lechoso (R3), 68.2 de grano lechoso a grano masozo (R4), 106 de grano mañoso a grano duro (R5) y 102.5 de grano duro a madurez fisiológica (R6).
CONCLUSIONES
Con la aplicación de fósforo se afectó la R del cultivo en maíz; la R mayor de Cacahuazintle se atribuye a su mejor adaptación a la zona de estudio y a su reacción mayor a la ausencia de este nutrimento que Amarillo Almoloya y Cóndor. Los valores elevados de EUR coinciden con los rendimientos altos en de Cacahuazintle y Cóndor. El cultivo de maíz alcanzó su desarrollo ontogénico adecuado desde la siembra hasta la madurez fisiológica, con una integral térmica de 1158.6 UC para la zona de estudio.
LITERATURA CITADA
Aceves, R. E., F. A. Turrent, F. J. I. Cortéz, y H. V. Volke. 2002. Comportamiento agronómico del híbrido H–137 y materiales criollos de maíz en el valle de Puebla. Rev. Fitotec. Mex. 25(4): 339–347. [ Links ]
Aguilar, L., M. G., S. A. J. Carrillo, P. A. Rivera, y V. A. González H. 2006. Análisis de crecimiento y relaciones fuente demanda en dos variedades de papa (Solanum tuberosum L.) Rev. Fitotec. Mex. 29(2): 145–156. [ Links ]
Andrade, F. H., S. A. Uhart, and A. G. Cirilo. 1993. Temperature effects radiation use efficiency in maize. Field Crops Res. 32: 17–25. [ Links ]
Arenas, J. Y. R., M. R. Delgado, R. E. J. Morales, C. A. Laguna, M. O. Franco, y S. E. Urbina. 2011. Rendimiento de raíces tuberosas de Dahlia variabilis wild (Desf.) bajo diferentes prácticas de manejo agronómico. ΦYTON. 80: 107–112. [ Links ]
Castillo, A. M., G. I. Nikolskii, S. C. A. Ortiz, H. Vaquera H., B. G. Cruz, S. E. Mejía, y H. A. González. 2007. Alteración de la fertilidad del suelo por el cambio climático y su efecto en la productividad agrícola. Interciencia 32(6): 368–376. [ Links ]
De la Casa, A., G. Ovando, L. Bressanini, J. Martínez, y A. Rodríguez. 2011. Eficiencia en el uso de la radiación en papa estimada a partir de la cobertura del follaje. Agriscientia 28: 21–30. [ Links ]
Díaz, L. E., P. J. M. Campos, R. A. Morales, B. G. Salgado, V. A. Castillo, y G. H. Gil. 2012. Ciencias Agrícolas Informa 21(2): 86–96. [ Links ]
Díaz, L. E., R. E. J. Morales, M. O. Franco, y L. A. Domínguez 2011. Atenuación de luz, radiación interceptada y rendimiento del maíz en función del fósforo. Terra Latinoamericana 29: 65–72. [ Links ]
Díaz, L. E., J. A. Escalante E., M. T. Rodríguez, G., y E. A. Gaytán A. 2011. Producción de frijol ejotero en función del tipo de espaldera. Rev. Chapingo Ser. Hortic. 16(3): 215–221. [ Links ]
Flores, L. R., C. F. Sánchez, P. J. E. Rodríguez, A. R. Mora, M. T. Colinas L., y S. H. Lozaya. 2009. Influencia de la radiación solar en la producción de semilla — tubérculo de papa bajo cultivo sin suelo. Rev. Chapingo Ser. Hortic. 15(1): 25–30. [ Links ]
García, E. 2005. Modificaciones al Sistema de Clasificación Climática de Köppen. Ed. UNAM. México. 217p. [ Links ]
García, P. A. D., y C. López C. 2002. Temperatura base y tasa de expansión foliar del maíz. Rev. Fitotec. Mex. 25(4): 381–386. [ Links ]
Giménez, C., D. J. Connor, and F. Rueda. 1994. Canopy development, photosynthesis and radiation use efficiency in sunflower in response to nitrógeno. Field Crops Res. 15: 17–37. [ Links ]
González, H. A., C. J. Sahagún, L. D. Pérez, L. A. Domínguez, C. R. Serrato, F. V. Landeros, y C. E. Dorantes. 2006. Diversidad fenotípica del maíz cacahuazintle en el Valle de Toluca, México. Rev. Fitotec. Mex.. 29(3): 255–261. [ Links ]
González, H. A., G. L. M. Vázquez, C. J. Sahagún, P. J. E. Rodríguez, y L. D. J. Pérez. 2007. Rendimiento de maíz de temporal y su relación con la pudrición de mazorca. Agric. Técnica Méx. 33(1): 33–42. [ Links ]
Kantolic, A. G., and G. A. Slafer. 2001. Photoperiod sensitivity after flowering and seed number determination in indeterminate soybean cultivars. Field Crops Res. 72: 109–118. [ Links ]
Mercau, J. L., V. O. Sadras, E. H. Satorre, C. Messina, C. Balbi, M. Uribelarrea, and A. J. Hall. 2001. On–farm assessment of regional and seasonal variation in sunflower yield in Argentina. Agric. Syst. 67: 83–103. [ Links ]
Morales, R. E. J., E. J. A. Escalante, y S. J. A. López. 2009. Biomasa y rendimiento de frijol con potencial ejotero en unicultivo y asociado con girasol. Rev. Chapingo Ser. Hortic. 15(1): 33–39. [ Links ]
Morales, R. E. J., E. J. A. Escalante, Ch. L. Tijerina, H. V. Volke, y M. E. Sosa. 2006. Biomasa, rendimiento, eficiencia en el uso del agua y de la radiación solar del agrosistema frijol–girasol. Terra Latinoamericana 24(1): 55–64. [ Links ]
Pearcy, R. W., J. Ehleringer, H. A. Mooney, and P. W. Rundel. 1994. Plant Physiological Ecology. Chapman & Hall. Great Britain. 457 p. [ Links ]
Rivetti, R. A. 2007. Producción de maíz bajo diferentes regímenes de riego complementario en Río Cuarto, Córdoba, Argentina. II. Producción de materia seca. Rev. Fac. Cienc. Agrar. 38(2): 25–36. [ Links ]
Salisbury, F. B., y C. W. Ross. 1994. Fisiología Vegetal. Grupo Editorial Iberoamérica. pp: 180–204. [ Links ]
SAS Institute. 2002. SAS/STAT 9.0 User's guide. SAS Inst., Cary, NC. [ Links ]
Soil Science Society of America. 2000. Conversion factor for SI and non — SI units. 44 p. [ Links ]
Taiz, L., and E. Zeiger. 2002. Plant Physiology. Sinauer. U.S.A. pp: 145–150. [ Links ]

