










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.46 no.5 Texcoco jul./ago. 2012
Ciencia Animal
Influence of season on intramuscular fatty acid profile of fighting bull
Efecto estacional sobre el perfil de ácidos grasos de la grasa intramuscular del toro de lidia
Alberto Horcada-Ibáñez1 , Oliva Polvillo-Polo1'2, Mercedes Valera-Córdoba1, Manuel Juárez-Davila1,3
1 MERAGEM Research Group. Universidad de Córdoba. Campus de Rabanales. Edif. Gregor Mendel. 14014 Cordoba, Spain.
2 Centro de Investigación, Tecnología e Innovación Universidad de Sevilla. Avda. Reina Mercedes, 4-B, 41012 Seville, Spain.
3 Lacombe Research Centre, Agriculture and Agri-Food Canada. 6000 C & E Trail. T4l 1W1 Lacombe, AB, Canada. *Author for correspondence. (albertohi@us.es).
Received: October, 2011.
Approbed: July, 2012.
Abstract
In Spain, beef from Fighting Bulls is traditionally consumed during spring and summer festivals but there are few studies on fatty acid profile from of these bulls. The objective this study was to evaluate the effect of slaughtering season and year of breeding on fatty acid composition of Spanish Fighting Bull intramuscular fat. The study was carried out during 2006 and 2009 seasons in Seville and Pamplona using a sampling survey methodology. Fighting Bulls (n=150) were raised in a traditional production system (Spanish dehesa) and slaughtered in spring (Sevilla) and summer (Pamplona). Supraespinatus muscle was collected at the abattoir 24 h postmortem for fatty acid analysis. Statistical analysis included location and slaughter as main effects and livestock farm as random effect. Bulls slaughtered in summer were older (p = 0.027) and heavier (p=0.002), but spring carcasses were heavier (p=0.008), with higher yields (p≤0.001), and a higher degree of fatness (p≤0.001). However, meat from bulls slaughtered in spring showed lower (p≤ 0.001) intramuscular fat content, leading to a higher (p≤ 0.001) polyunsaturated/ saturated fatty acid ratio. Intramuscular fat showed seasonal differences (p≤ 0.05) in most fatty acid concentrations. There were significant interactions (p≤ 0.05) between the slaughter season and years for several fatty acids, total lipids and monounsaturated. Conjugated linolenic acid (CLA) levels were similar to those found in conventional beef. The higher monounsaturated fatty acids and CLA content in bulls slaughtered at summer could be partly due to mobilization of polyunsaturated fatty acids in this period. Thus, although the intramuscular fat of Spanish Fighting Bulls may have a high nutritional value, the great influence of the rearing season and slaughter system should be considered when fatty acid composition is concerned.
Key words: traditional production system (dehesa), Fighting Bull, lipids, season.
Resumen
En España, la carne de Toro de Lidia se consume tradicionalmente durante los festivales de primavera y verano, pero hay pocos estudios sobre el perfil de ácidos grasos de esta carne. El objetivo de este estudio fue evaluar el efecto de la estación de sacrificio y el año de crianza en la composición de ácidos grasos de la grasa intramuscular del Toro de Lidia español. El estudio se realizó durante las temporadas 2006 y 2009 en Sevilla y Pamplona con una metodología de encuesta de muestreo. Los Toros de Lidia (n= 150) fueron criados en un sistema de producción tradicional (dehesa española) y sacrificados en primavera (Sevilla) y verano (Pamplona). A las 24 h post-mortem se tomó una muestra del músculo Supraespinatus en el matadero para analizar ácidos grasos. El análisis estadístico incluyó ubicación y año de sacrificio como efectos principales y la hacienda ganadera como efecto aleatorio. Los toros sacrificados en verano tenían mayor edad (p = 0.027) y más peso (p = 0.002), pero las canales en primavera fueron más pesadas (p = 0.008), con rendimientos más altos (p≤ 0.001), y mayor gordura (p≤ 0.001). La carne de los toros sacrificados en primavera mostró contenido de grasa intramuscular menor (p≤ 0.001), y una relación de ácidos grasos poliinsaturados/saturados mayor (p≤ 0.001). La grasa intramuscular mostró diferencias estacionales (p≤0.05) en los ácidos grasos mayoritarios. Hubo interacciones significativas (p≤ 0.05) entre época de sacrificio y año para varios ácidos grasos, lípidos totales y ácidos grasos monoinsaturados. Los niveles de ácido linolénico conjugado (CLA) fueron similares a los de la carne bovina convencional. El contenido más alto de ácidos grasos monoinsaturados y de CLA en toros sacrificados en el verano podría deberse en parte a la movilización de ácidos grasos poliinsaturados en este período. Así, aunque la grasa intramuscular del Toro de Lidia español puede tener un valor nutricional alto, la gran influencia de la época de cría y el sistema de sacrificio debieran considerarse en lo que respecta la composición de ácido graso.
Palabras clave: sistema de producción tradicional (dehesa), Toro de Lidia, lípidos, estación.
INTRODUCTION
Selection strategies and production systems used to raise Fighting Bulls are unique since the bulls are selected based on behaviour (aggressiveness, strength and vigour) and external traits (skin colour or horn shape and size). The traditional production system for Fighting Bulls is a free range semi-extensive breeding system using a concentrate supplement during the summer due to grass shortage, and bulls are sent to the bullring at 4-4 ½ years (Horcada et al., 2010). Few of the bulls produced each year are used for bull fighting and most of them (males and females) are ruled out after selection tests and sent to the abattoir. Besides, carcasses from Fighting Bulls slaughtered at the bullring must be processed in an official abattoir and then sold at meat markets.
Fighting Bull meat is produced and consumed in Spain, France, Portugal, México, Colombia, Venezuela, Ecuador and Perú. It has a marked seasonal trend which depends on the dates bull fightings take place. Meat from bulls slaughtered at the bullring can reach higher prices than regular beef, while meat from bulls slaughtered at the abattoir is sold at lower price due to lower carcass weight and beef quality grade.
Although genotype and diet are two of the main factors affecting intramuscular fatty acid (FA) composition, studies available on meat FA profile from Fighting Bull are scarce (Beriain et al., 2011). In 2002, RD 260/2002 was issued in Spain regarding Fighting Bulls meat commercialization. In addition, beef associated to organic production systems in dehesa (Fighting Bull producing farms) can be considered as a new niche product for development of rural areas (Joffre et al., 1999).
Several factors, such as genetic background, diet, slaughter weight, and fatness degree influence FA profile (Wood et al., 2008) but little is known about beef FA composition of Spanish Fighting Bulls. Besides, due to the traditional seasonal production, lipid profile may present high variability since there are seasonal and climatic effects on FA composition (Marchello et al., 1967; Perry et al., 1998). The objective of this research was to study the effect of slaughtering season (spring and summer) on FA profile of intramuscular fat from Spanish Fighting Bull raised on dehesa areas according to traditional production system.
MATERIAL AND METHODS
Experimental design and sampling
The study was carried out on 150 Fighting Bulls during 2006 and 2009 in Spain. In 2006, samples were collected from 30 bulls slaughtered in spring during the April Festival ("Feria de Abril", Seville) and from 30 bulls slaughtered in summer during the "San Fermín" festival (Pamplona). In 2009, samples from 45 bulls per festival were collected. These are two of the main events involving Fighting Bulls in Spain and are representative of the seasonality in Fighting Bull slaughters.
Bulls were produced in 20 livestock farms located in Southeast Spain. The bulls remained at the dehesa fields (Joffre et al., 1999) for 4 V years until transportation to the festival. Fighting Bulls grazed ad libitum on a permanent native pasture and received a concentrate suplement (14.1-15.0 % protein, 5.05-5.40 % fibre, 4.5-5.30 % total fat, 32.1-37.0 % starch, 1.00-1.10 % Ca and 0.25-0.40 % P). The average botanical composition of the native pasture was: 25 % grass (mainly Lolium, Bromus, Agrostis and Pod) and 26 % legumes (mainly Trifolium, Medicago and Ornithopus).
Bulls were transported (250 km to Seville and 845 km to Pamplona) according to welfare specifications of the Council Directive 86/609/EEC (European Communities, 1986) regulation, and slaughtered according to RD 1034/2001 regulation. In the spring bulls were slaughtered within 24 h after arrival to the abattoir. In summer, the bulls remained 10 d in individual stables and fed concentrate and barley straw ad libitum before being slaughtered.
After slaughter and cleaning, carcasses remained 4 h at room temperature (18 °C) and then refrigerated 24 h (2 °C). Carcasses were then weighed and conformation and degree of fatness assessed according to the European classification system (EEC Regulation No. 1183/2006). Samples were then taken from the supraespinatus muscle, vacuum packed and frozen at —20 °C until FA analysis.
Fatty acid analysis
Total FA were extracted, methylated and analysed at the General Agricultural Research Service, University of Seville (Spain), using an modification of the method described by Aldai et al. (2006). Fatty acid methyl ester (FAME) separation and quantification was carried out using a gas chromatograph (GC, Agilent 6890N, Inc., California, USA) equipped with a flame ionisation detector (FID) and with a BPX-70 capillary column (12.0 m, 0.25 mm i.d., 0.2 ^im film thickness, SGE, Australia), as reported by Juárez et al. (2009). Individual FAMEs were identified using standard (Sigma Chemical Co. Ltd., Poole, UK). Content of FA were expressed as percentage of total FAs, and grouped as follows: saturated (SFA), monounsaturated (MUFA), polyunsaturated (PUFA), n-3 and n-6. Content of trans octadecenoic (∑trans 18:1) and conjugated linolenic (∑CLA) FAs was expressed as a single value because of its incomplete chromatographic resolution. Besides, PUFA/SFA and Δ-9 desaturase activity [C18:1/(C18:0+C18:1)] indices were calculated (Malau-Aduli et al., 1998).
Statistical analysis
Statistical analysis was carried out with the MIXED procedure (SAS Institute, Inc. 2003). Location and year of slaughter and interactions were included as main effects; and producing farms as the random effect. LSMEANS and PDIFF options were applied for generating least squares means and comparison of treatments by F-test. Superscripts were used to separate means only when interaction between location and year was significant (p≤ 0.05).
RESULTS AND DISCUSSION
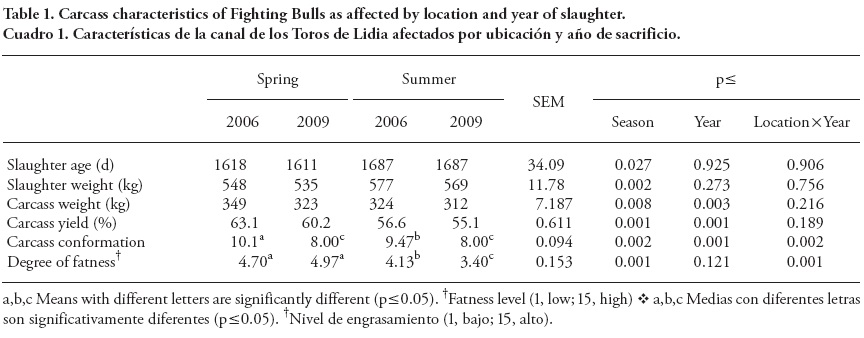
Bulls slaughtered in summer 2006 and 2009 were older (p = 0.027) and heavier (p = 0.002) than those slaughtered in spring (Table 1). Average age and weight of slaughter (4 V years and 557 kg) cannot be compared to those commonly used for commercial beef production in Spain (Serra et al., 2008) and other countries (Christensen et al., 2011), since Fighting Bulls are not specifically selected or raised for meat production, but rather for their aggressive behaviour. The difference in age between slaughtering seasons can be explained by the dates of festival celebration. This could be also linked to differences in weight, as during summer there is little grass for the bulls and concentrate is then supplied.
Spring carcasses were heavier (p= 0.008) and with higher yields (p≤ 0.001) than summer carcasses. Longer transportation in hot climates and fasting previous to slaughtering in summer could be the cause for this effect. Carcasses were lighter (p = 0.003) and yields lower (p≤ 0.001) in 2009 than in 2006 for both seasons. The production system used in Fighting Bull breeding is very susceptible to environmental variations because animals always remain in the field ( dehesa). Changes on the precipitation rate lead to changes in pasture availability during the seasons, affecting performance and carcass traits. Thus, annual precipitation average was higher in 2006 (518 mm) than in 2009 (474 mm) in dehesa where bulls were raised (MARM, 2011).
Besides, carcass conformation was lower in spring 2009 and it was higher in summer 2006 (p= 0.002). As shown by the interaction effect (p≤ 0.001), the degree of fatness was higher in 2006 and 2009 in bulls slaughtered in spring as compared to those slaughtered in summer. A sharp decrease in fatness degree was observed in bulls slaughtered in summer due to FA mobilization from the adipose tissue.
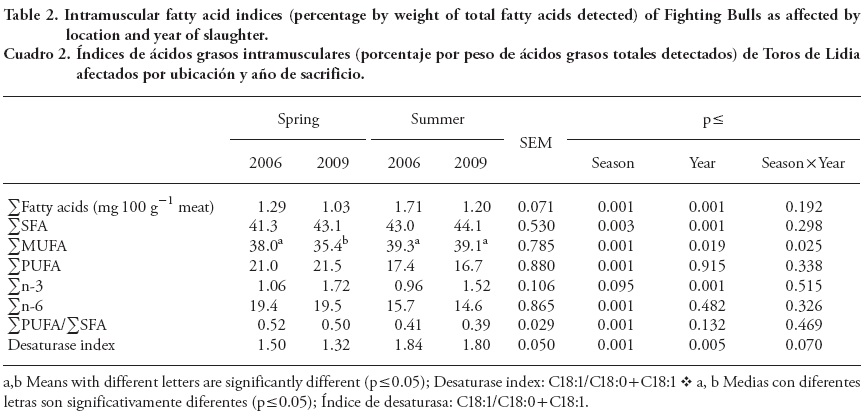
Total FAs and FA indices (% of total FAs) in intramuscular fat of Fighting Bulls in late spring and early summer are shown in Table 2. Data interpretation regarding FA composition could be difficult due to uncontrollable environmental factors. However, Aldai et al. (2009) suggested comparing FA composition of a given meat or meat product to health recommendations when limited or no information is available.
Average total FA content in Fighting Bulls beef was lower than that reported for longissimus (Christensen et al., 2011) or diaphragm (Dugan et al., 2010) muscles, using different bovine breeds and production systems. Fighting Bulls show poor marbling which may satisfy the Spanish consumers' demand for lean and low marbled beef (Beriain et al., 2011). In fact, the levels of intramuscular fat observed in the present study are closer to alternative species such as buffalo (Juárez et al., 2010) or bison (Janz et al., 2000), than to commercial beef. This is due to differences in both genotype and production systems. There was an effect of year on fatty acid content (Table 2) because in 2009 bulls showed lower levels of total fat content (p≤ 0.001) than those observed in 2006, which can be explained by variations on the precipitation rate and grass availability between years. Moreover, there was no interaction between year and season since in all cases meat from bulls slaughtered in spring had lower (p≤ 0.001) total fat content than in bulls slaughtered in summer, when there was less grass available and the bulls received a larger amount of concentrate. Therefore, the higher amount of energy provided by the concentrate as compared to the forage (Caton and Dhuyvetter, 1997) and the difference in age at slaughter could explain the differences in total fat content between spring and summer.
The low intramuscular fat content in Fighting Bulls led to FA composition characteristic of lean meats, high PUFA content (19.2 mg 100 g -1 meat mean value). According to Wood et al. (2008), low total lipid concentration in muscle with a high phospholipid proportion will lead to a higher PUFA proportion in total lipids. However, PUFA content in beef intramuscular fat is usually lower (Juárez et al., 2011; Noci et al., 2007) than that observed in the present study. Insausti et al. (2004) reported similar values in lean beef (<1.5 % fat). Content of SFA increased in intramuscular fat at both seasons (p≤ 0.001) in 2009; and in 2006 intramuscular fat from bulls slaughtered in spring had lower (p= 0.003) total SFA content and higher (p< 0.001) total PUFA as compared to bulls slaughtered in summer.
Total MUFA level in spring 2009 was the lowest, whereas total n-3 level increased in bulls at both seasons in 2009 (p≤ 0.001), and n-6 content was higher in bulls slaughtered in spring (p≤ 0.001). A decrease in total intramuscular fat results in a higher neutral lipid to phospholipid ratio; neutral lipids being rich in SFA and phospholipids in n-3 FA (Riely et al. 2000). Christie (1981) and Realini et al. (2004) report an increase in fat and a decrease in PUFA proportion in beef when concentrate was included in a forage-based diet. Seasonal and climatic effects have been also reported for FAs composition of ruminants (Marchello et al., 1967; Perry et al., 1998).
Thus, differences in feed quality affect pasture-fed rather than grain-fed cattle, which can explain the increase in MUFA in 2009 when the availability of grass was lower and supplement was higher. In a survey in Canada, Aldai et al. (2009) reported an increase in MUFA levels in retail beef during winter when concentrate is required, which is not the case in Spain. Moreover, climatic differences (cold versus hot regions) may also lead to changes in fat composition. In the present study, average PUFA/SFA ratio was 0.45; higher (p ≤ 0.001) in meat from bulls slaughtered in spring due to the higher PUFA content. Current nutritional recommendations establish that PUFA/ SFA ratio in human diets should be above 0.4, fats with low PUFA/SFA ratio are not recommended because they might increase cholesterolemia (British Department of Health, 1994). Therefore, according to our results and in agreement with Beriain et al. (2011), meat from Fighting Bulls has a balanced PUFA/SFA ratio for human health.
The high MUFA content in bulls slaughtered in summer could due to energy storage mobilization in fat depots. Patterns of relative FA mobilization in mammals are different (Gavino and Gavino, 1992; Price et al., 2008) and selective mobilization occurs in an undernutrition situation (Connor et al., 1996). Raclot and Groscolas (1993) show that FA mobilization from the adipose tissue was positively correlated to degree of unsaturation (fundamentally PUFA), and negatively to chain length. In in vitro studies, short chain FA with doubles bonds closer to the methyl end are preferentially mobilized (Raclot, 2003), which could also explain in vivo results in our study. There was a sharp decrease of fatness degree in bulls slaughtered in summer (Table 1), suggesting undernutrition due to less pasture, higher environmental temperature, longer transportation time, and pre-slaughter fasting. It can be concluded that environmental conditions and differences in the availability and quality of feeding have an effect on intramuscular FA composition (Mazzone, 2010). Besides, selective FA mobilization takes place during high energy demand, undernutrition and stress situations (Price et al., 2008; Soppela and Nieminen, 2002). The lower PUFA content and higher Δ-9 desaturase activity (p≤ 0.001) in fat of bulls slaughtered in summer (Table 2) were also in agreement with higher PUFA mobilization in undernutrition and stress situations. Raclot (2003) report differential FA mobilization according to the molecular structure and saturation degree; polyunsaturated FA are preferentially mobilized.
The most abundant FA were C16:0, C18:0, C18:1n-9 and C18:2n-6 (Table 3) which were observed in Spanish beef by Indurain et al. (2006). The second most abundant FA in spring was C18:0, and 16:0 (p≤ 0.001) in summer due to the higher supplementation with concentrate. French et al. (2000) report a linear decrease in SFA proportion when increasing grass intake due to a lower 16:0 content in grass as compared to concentrate. As observed in SFA content, C18:0 concentration was higher for both 2009 seasons, as compared to 2006 (p≤ 0.001). This is the only saturated FA with a net neutral impact on serum cholesterol and the reduction in C18:0 could be related to lower intramuscular fat level in 2009.
Among the four experimental groups, the concentration of C18:1n-9c, the most abundant MUFA, was at a minimum in spring 2009, whereas total C18:1 trans concentration was the lowest in summer 2009 (p= 0.013) and increased for both seasons in 2006 (p≤ 0.001). These FAs result from incomplete microbial hydrogenation of unsaturated FAs in the rumen (Bessa et al., 2000). The impact of FA concentration on human health is difficult to interpret without a complete isomer profile, since different isomers may have diverse effects. According to Dugan et al. (2008), the most abundant trans isomer in beef cattle fed with 73 % barley grain diet is C18:1(10t) rather than C18:1(11t), which suggests a negative impact of C18:1(10t ) on LDL cholesterol and cardiovascular diseases.
The increase in C18:2n-6, C18:3n-3 and C20:4n-6 (p≤ 0.001) in meat from bulls slaughtered in spring could be due to the higher consumption of pasture (Palmquist, 1988); particularly, C18:3n-3 concentration was high in pasture. Content of this FA depends on the season, and it is rapidly and extensively biohydrogenated in the rumen. Therefore, its higher availability from fresh herbage can increase the content in muscle fat. In fact, bulls exposed to more abundant pastures (late winter and early-middle spring grass) are not finished on concentrate (these bulls were slaughtered in April), whereas bulls exposed to less abundant pastures (early summer grass) and finished on concentrate during three months were slaughtered in July. The increased concentration in 2009 of n-3 FA, such as C20:5n-3 (p≤ 0.001), C22:5n-3 (p≤ 0.001) and C22:6n-3 (this last one only in spring; p= 0.017) was mainly related to the decrease in total intramuscular fat.
A greater PUFA mobilization, associated to undernutrition in summer and 10 days in individual stables, could be another cause for MUFA and CLA increase bulls slaughtered in aummer, although they consumed less grass. The increase in ruminal FA depends on PUFA mobilization. Ruminal PUFA, mostly CLA isomers, could be intermediates of C18:3n-3 biohydrogenation (Biondi et al., 2008). The low 18:3n-3 content observed in bulls slaughtered in summer suggested an increase in CLA production of CLA from C18:3n-3, partly due to a more intense fat mobilization during this season.
Finally, total CLA concentration showed an interactive effect between season and year (p≤ 0.001), being significantly lower in spring 2009. Natural augmentation of CLA within the lipid fraction of Fighting Bull meat can be accomplished through diets rich in grass and lush green forages. While CLA precursors can be found in both grains and lush green forages, grass-fed ruminant species produce 2 to 3 times more CLA than ruminants fed high grain diets, largely due to a more favourable rumen pH (Rule et al., 2002; Smith et al., 2002). As far as we know, the increase of CLA percentage in concentrate-fed ruminants has not yet been reported. Although little research has been conducted to assess seasonal changes in beef CLA, Lock and Garnsworthy (2003) observed that CLA percentages in milk fat varied throughout the year in UK, with the highest values registered in the summer (May-July), when cows received fresh grass. Furthermore, as aforementioned, variations in temperature and rainfall affect FA composition of free-ranged cattle in several ways (Tume, 2004). Thus, it may be that suggested the low quality of the grass available during 2009 in southern Spain may have been unfavourable for the production of CLA in the rumen, explaining the low content in that year.
CONCLUSIONS
There was a significant effect of breeding year and slaughtering season on the characteristics of intramuscular fat in Fighting Bulls. Intramuscular fat composition showed particular characteristics due to the use of a unique combination of breed and production system. A significant influence of the rearing season has to be considered in fatty acid quality of Fighting Bulls meat, primarily due to feed fatty acid composition supplied in a given season and secondly due to the fat mobilization effect in stressing conditions in summer. From a nutritional viewpoint, meat from Fighting Bulls has a well balanced PUFA/ SFA ratio. Fatty acids composition from these bulls are comparable in quality and nutritive values to other ruminants, and the high CLA and PUFA contents suggest that beef from Fighting Bulls could be a good alternative for human consumption.
ACKNOWLEDGEMENTS
The authors thank Vacuno de Navarra S. L. and Mercasevilla for their contribution to the development of this study.
LITERATURE CITED
Aldai, N., K. Osoro, L. Barron, and A. Nájera. 2006. Gasliquid chromatographic method for analysing complex mixtures of fatty acids including conjugated linoleic acids (cis-9, trans-11 and trans-10, cis12 isomers) and long-chain (n-3 or n-6) polyunsaturated fatty acids-Application to the intramuscular fat of beef meat. J. Chromat. A. 1110:133-139. [ Links ]
Aldai, N., M. E. R. Dugan, D. C. Rolland, and J. K. G. Kramer. 2009. Survey of the fatty acid composition of Canadian beef: Backfat and longissimus lumborum muscle. Can. J. Anim. Sci. 89: 315-329. [ Links ]
Beriain, M.J., A. Horcada, G. Lizaso, K. Insausti, and A. Purroy. 2011. Meat quality from Fighting Bulls in Spain. Rev. Cient. FCV-LUZ 21: 88-95. [ Links ]
Bessa, R., J. Santos-Silva, J. Ribeiro, and A. Portugal. 2000. Reticulorumen biohydrogenation and the enrichment of ruminant edible products with linoleic acid conjugatedisomers. Liv. Prod. Sci. 63: 201-211. [ Links ]
Biondi, L., M. A. Valvo, M. Di Gloria, E. Scinardo, V. Galorafo, and A. Priolo. 2008. Change in ewe milk fatty acids following turning out to pasture. Small Rum. Res. 75: 17-23. [ Links ]
British Department of Health. 1994. Nutritional aspects of cardiovascular disease. Report on health and social subjects no. 46. H.M. Stationery Office, London. [ Links ]
Caton, J., and D. Dhuyvetter. 1997. Influence of energy supplementation on grazing ruminants: requirements and responses. J. Anim. Sci. 75: 533-542. [ Links ]
Christensen, M., P. Ertbjerg, S. Failla, C. Sañudo, R. I. Richardson, G. R Nute, J. L. Olleta, B. Panea, P. Alberti, M. Juárez, J. F. Hocquette, and J. L. Williams. 2011. Relationship between collagen characteristics, lipid content and raw and cooked texture of meat from young bulls of fifteen European breeds. Meat Sci. 87: 61-65. [ Links ]
Christie, W. 1981. The composition, structure and function of lipids in the tissues of ruminant animals. In: Christie, W. (eds). Lipid Metabolism in Ruminant Animals. Oxford. Pergamon Press. pp: 95-191. [ Links ]
Council Directive 86/609/EEC 24 November 1986 on the approximation of laws, regulations and administrative provision of the Member States regarding the protection of animals used for experimental and other scientific purposes. OJ No. L 358, 18.12.1986 p. 1. [ Links ]
Connor, W., D. Lin, and C. Colvis. 1996. Differential mobilization of fatty acids from adipose tissue. J. Lipid Res. 37: 290-298. [ Links ]
Dugan, M., D. Rollan, J. Aalhus, N. Aldai, and J. Kramer. 2008. Subcutaneous fat composition of youthful and mature Canadian beef: emphasis on individual conjugated linoleic acid and trans-18:1 isomers. Can. J. Anim. Sci. 88: 591-599. [ Links ]
Dugan, M., N. Aldai, J. Kramer, D. Gibb, M. Juárez, and T. McAllister. 2010. Feeding wheat dried distillers grains with solubles improves beef trans and conjugated linoleic acidprofiles. J. Anim. Sci. 88: 1842-1847. [ Links ]
EEC Regulation No 1183/2006 del Consejo de 24 de julio de 2006 sobre el modelo comunitario de clasificación de las canales de vacuno pesado, pp: L214/1-L214/6 [ Links ]
French, P., E. O'Riordan, F. Monahan, P. Caffrey, M. Vidal, M. Mooney, D. Troy, and A. Moloney. 2000. Meat quality of steers finished on autumn grass, grass silage concentrate-based diets. Meat Sci. 56: 173-180. [ Links ]
Gavino, V. C., and G. R. Gavino. 1992. Adipose hormone sensitive lipase preferentially releases polyunsaturated fatty acids from triglycerides. Lipids 27: 950-954. [ Links ]
Horcada, I. A., A. Luque M., and J. M. Jiménez J. 2010. La producción bovina en Andalucía. In: La producción de carne en Andalucía. Junta de Andalucía editor. Seville, Spain. pp: 297-316. [ Links ]
Indurain, G., M. J. Beriain, M. V. Goñi, A. Arana, and A. Purroy. 2006. Composition and estimation of intramuscular and subcutaneous fatty acid composition in Spanish youngbulls. Meat Sci. 73: 326-334. [ Links ]
Insausti, K., M. J. Beriain, M. J. Alzueta, T. R. Carr, and A. Purroy. 2004. Lipid composition of the intramuscular fat of beef from Spanish cattle breeds stored under modified atmosphere, Meat Sci. 66: 639-646. [ Links ]
Janz, J., J. Aalhus, M. Price, and A. Schaefer. 2000. The influence of elevated temperature conditioning on bison (Bison bison) meat quality. Meat Sci. 56: 279-284. [ Links ]
Joffre, R., S. Rambal, and J. P. Ratte. 1999. The dehesas system of southern Spain and Portugal as a natural ecosystem mimic. Agrofor. System. 45: 57-79. [ Links ]
Juárez, M., A. Horcada, M. J. Alcalde, A. Molina, M. Valera, and O. Polvillo. 2009. Meat and fat quality of unweaned lambs as affected by slaughter weight and breed. Meat Sci. 83: 308-313. [ Links ]
Juárez, M., S. Failla, A. Ficco, F. Peña, C. Avilés, and O. Polvillo. 2010. Buffalo meat as affected by different cooking methods. Food Bioprod. Proced. 88: 145-148. [ Links ]
Juárez, M., M. Dugan, J. Aalhus, N. Aldai, J. Basarab, V. Baron, and T. McAllister. 2011. Effects of vitamin E and flaxseed on rumen-derived fatty acid intermediates in beef intramuscular fat. Meat Sci. 88: 434-440. [ Links ]
Lock, A., and P. Garnsworthy. 2003. Seasonal variation in milk conjugated linoleic acid and A9-desaturase activity in dairy cows. Liv. Produc. Sci. 79: 47-59. [ Links ]
Malau-Aduli, A., B. Siebert, C. Bottema, and W. Pitchford. 1998. Breed comparison of the fatty acid composition of muscle phospholipids in Jersey and Limousin cattle. J. Anim. Sci. 76: 766-773. [ Links ]
Marchello, J., D. Cramer, and L. Miller. 1967. Effect of ambient temperature on certain ovine fat characteristics. J. Anim. Sci. 26: 294-297. [ Links ]
MARM. 2011. Información Pluviométrica. Boletín Hidrológico Semanal, 7. Ministerio de Medio Ambiente y Rural y Marino. (http://www.marm.es). [ Links ]
Mazzone, G., M. Giammarco, G. Vignola, L. Sardi, and L. Lambertini. 2010. Effects of the rearing season on carcass and meat quality of suckling Apennine light lambs. Meat Sci. 86: 474-478. [ Links ]
Noci, F., P. French, F. Monahan, and A. Moloney. 2007. The fatty acid composition of muscle fat and subcutaneous adipose tissue of grazing heifers supplemented with plant oil-enriched concentrates. J. Anim. Sci. 85: 1062-1073. [ Links ]
Palmquist, D. 1988. The feeding value of fat. In: Orskov, E. R. (ed). Feed Science. Elsevier, Amsterdam. pp: 293-311. [ Links ]
Perry, D., P. Nicholls, and J. Thompson. 1998. The effect of sire breed on the melting point and fatty acid composition of subcutaneous fat in steers. J. Anim. Sci. 76: 87-95. [ Links ]
Price, E., A. Krokfors, and C. Guglielmo. 2008. Selective mobilization of fatty acids from adipose tissue in migratory birds. J. Exper. Biol. 211: 29-34. [ Links ]
Raclot, T. 2003. Selective mobilization of fatty acids from adipose tissue triacylglycerols. Progr. Lipid Res. 42: 257-288. [ Links ]
Raclot, T., and R. Groscolas. 1993. Differencial mobilization of white adipose tissue fatty acids according to chain length, unsaturation, and positional isomerism. J. Lipid Res. 34:1515-1526. [ Links ]
RD 1034/2001 2001. Real Decreto 1034/2001, de 21 de septiembre, por el que se modifica parcialmente el Reglamento de Espectáculos Taurinos, aprobado por Real Decreto 145/1996, de 2 de febrero. BOE, 240/2001 de 6 de octubre de 2001. [ Links ]
RD 260/2002, de 8 de marzo, por el que se fijan las condiciones sanitarias aplicables a la producción y comercialización de carnes de reses de lidia. BOE, 64 de 15 de marzo de 2002. [ Links ]
Realini, C., S. Duckett, G. Brito, M. Dalla Rizza, and D. De Mattos. 2004. Effect of pasture vs. concentrate feeding with or without antioxidants on carcass characteristics, fatty acid composition, and quality of Uruguayan beef. Meat Sci. 66: 567-577. [ Links ]
Riley, P., M. Enser, G. Nute, and J. Wood. 2000. Effects of dietary linseed on nutritional value and other quality aspects of pig muscle and adipose tissue. Anim. Sci. 71: 483-500. [ Links ]
Rule, D., K. Broughton, S. Shellito, and G. Maiorano. 2002. Comparison of muscle fatty acid profiles and cholesterol concentrations of bison, cattle, elk and chicken. J. Anim.Sci. 80 1202-1211. [ Links ]
SAS Institute Inc. 2003. Proceedings of the Twenty-Eighth Annual SAS® Users Group International Conference. Cary, NC: SAS Institute Inc. [ Links ]
Serra, X., L. Guerrero, M. D. Guardia, M. Gil, C. Sañudo, B. Panea, M. M. Campo, J. L. Olleta, M. D. García-Cachan, J. Piedrafita, and M. A. Oliver. 2008. Eating quality of young bulls from three Spanish beef breed-production systems and its relationships with chemical and instrumental meat quality. Meat Sci. 79: 98-104. [ Links ]
Smith, D., R. Wood, S. Tseng, and S. Smith. 2002. Increased beef consumption increases apolipoprotein A-1 but not serum cholesterol of mildly hypercholesterolemic men with different levels of habitual beef intake. Exp. Biol. Medic. 227: 266-275. [ Links ]
Soppela, P. and M. Nieminen. 2002. Effect of moderate wintertime undernutrition on fatty acid composition of adipose tissues of reindeer (Rangifer tarandus tarandus L.). Compar. Bioch. Phys. Part A: Molec. Integrat. Phys. 132: 403-409. [ Links ]
Tume, R. K. 2004. The effects of environmental factors on fatty acid composition and the assessment of marbling in beef cattle: A review. Aust. J. Expe. Agr. 44(7): 663-668. [ Links ]
Wood, J., M. Enser, A. Fisher, G. Nute, P. Sheard, R. Richardson, S. Hughes, and F. Whittington. 2008. Fat deposition, fatty acid composition and meat quality: a review. Meat Sci. 78: 343-358. [ Links ]


{kind=link}
{kind=link}