










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.46 no.3 Texcoco abr./may. 2012
Protección vegetal
Distribución espacial de Bactericera cockerelli (Sulc)(HEMIPTERA: TRIOZIDAE) en tomate de cáscara (Physalis ixocarpa (Brot.))
Spatial distribution of Bactericera cockerelli (Sulc) (HEMIPTERA: TRIOZIDAE) on green tomato (Physalis ixocarpa (Brot.))
Leonardo A. Crespo-Herrera1, Jorge Vera-Graziano1*, Hiram Bravo-Mojica1, José López-Collado1, Roberto Reyna-Robles1, Aureliano Peña-Lomelí2, Víctor Manuel-Pinto3, Ramón Garza-García4
1 Programa de Entomología y Acarología, Campus Montecillo, Colegio de Postgraduados. 56230. Km 36.5 Carretera México-Texcoco, Montecillo, Estado de México. *Autor responsable. (graziano@colpos. mx)
2 Departamento de Fitotecnia, Instituto de Horticultura.
4 Campo Experimental Valle de México, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. 56230. Estado de México, México.
Recibido: febrero, 2011.
Aprobado: marzo, 2012.
Resumen
El tomate de cáscara (Physalis ixocarpa Brot.) es hospedero de varias plagas, entre ellas Bactericera cockerelli (Sulc) que causa hasta 45 % de perdidas en rendimiento debido a la transmisión de un fitoplasma del grupo I. Sin embargo, no hay estimadores de los parámetros de los programas de muestreo para este insecto, los cuales son fundamentales para determinar tamaños de muestra de poblaciones. Por tanto, se estudió la distribución espacial de Bactericera cockerelli en tres parcelas experimentales en Texcoco, Estado de México, con base en los estimadores de la varianza/media (S2/X) y la Ley de la Potencia de Taylor (LPT). Los datos fueron analizados con el programa SUPRA. Los huevos y ninfas presentaron una disposición agregada, S2/X de 5 a 23.5 y 1.5 a 6.7; mientras que los adultos, altamente móviles en el campo, presentaron una distribución aleatoria (S2/X entre 0.83 y 20.60). El método LPT concuerda con los patrones de distribución para los tres estados biológicos de la plaga.
Palabras clave: tomatillo, psilido del tomate, ley de la potencia Taylor, relación varianza/media.
ABSTRACT
Green tomato (Physalis ixocarpa Brot.) is host to several pests, including Bactericera cockerelli (Sulc) that causes up to 45 % of yield losses due to transmission of a phytoplasm of group I. However, there are no estimates of the parameters of the sampling programs for this insect, which are crucial for determining sample size of populations. Therefore, we studied the spatial distribution of Bactericera cockerelli in three experimental plots in Texcoco, Estado de México, based on the estimates of the variance/mean (S2/X) and Taylor’s Power Law (TPL). Data were analyzed with the SUPRA program. The eggs and nymphs showed an aggregated arrangement (S2/X) of 5 to 23.5 and 1.5 to 6.7, while adults that are highly mobile in the field showed a random distribution (S2/X between 0.83 and 20.60). The TPL method agrees with the distribution patterns for the three biological stages of the pest.
Key words: green tomato, psyllid tomato, Taylor’s power law, variance/mean ratio.
INTRODUCCIÓN
El tomate de cáscara (Physalis ixocarpa Brot.) es la cuarta especie hortícola más cultivada en México, antecedida por el jitomate (Solanum lycopersicum), la papa (Solanum tuberosum) y el chile (Capsicum annuum) (Anónimo, 2002). Además es hospedero de diversas plagas que limitan su capacidad productiva cuando éstas encuentran condiciones favorables para su reproducción; entre ellas destaca Bactericera cockerelli (Sulc), la cual ataca también a otras especies cultivadas como jitomate, chile y papa (Almeyda et al., 2008). Este insecto se encuentra ampliamente distribuido en las principales regiones productoras de jitomate: Villa de Arista, San Luis Potosí, Yurécuaro, Michoacán, la región de La Laguna en los estados de Durango y Coahuila, San Quintín, Baja California, y en Morelos, Puebla, Guanajuato, Nayarit, Sinaloa y Estado de México (Vega et al., 2008). Bactericera cockerelli provoca dos tipos de daño en sus plantas hospedantes: 1) durante sus estados ninfales inyecta una toxina que causa clorosis en la planta y 2) transmite un fitoplasma que origina la enfermedad permanente del tomate, la cual produce hasta 45 % de pérdidas en rendimiento en las 30 000 ha cultivadas en México (Almeyda et al., 2008).
La distribución espacial se refiere a la ubicación que los individuos ocupan en un determinado tiempo y espacio (Vera et al., 2002), como las superficies cultivadas. Los estudios de distribución espacial tienen importancia ecológica (Southwood, 1978; Dajoz, 2002). Además, de ésta depende la forma de realizar muestreos para determinar los tamaños de muestra y dinámicas poblacionales de los insectos en los cultivos. Esta información es fundamental para diseñar sistemas de control de plagas, porque la distribución de una plaga influye en la efectividad y el comportamiento de parasitoides (Southwood, 1978; Maron y Harrison, 1997; Tripet et al., 2002).
En Chapingo, Estado de México, este insecto causa daños directos al succionar la savia de varias especies de solanáceas de importancia económica, así como daños indirectos al transmitir fitoplasmas (Vega et al., 2008). Por tanto, el objetivo de la presente investigación fue estudiar el patrón de distribución espacial de adultos, huevos y ninfas de B. cockerelli en el cultivo de tomate de cáscara (Physalis ixocarpa Brot.), que permita contribuir a la planeación de programas de muestreo y sistemas de control de insectos.
MATERIALES Y MÉTODOS
El 22 y 23 de marzo de 2007 se trasplantaron plántulas de tomate de cáscara del cultivar CHF1-Chapingo, y el cultivo se condujo mediante riego por goteo y sin aplicar insecticidas. Las observaciones de la presencia de insectos se realizaron del 30 de marzo al 11 de junio de 2007 en tres lotes experimentales de la Universidad Autónoma Chapingo, Texcoco, México (19° 29' 05" N y 98° 53' 11" O), y a una distancia aproximada de 1600 m entre sí. Los lotes fueron: Xaltepa-3 (X-3) con 324 m2 , San. Martín (Sn. M-11) con 1036 m2 , y Estación meteorológica-Chapingo (Ch) con 384 m2 . Las condiciones meteorológicas durante el periodo fueron: humedad relativa media diaria 53 % (de 7 a 90 %), temperatura media diaria 16.7 °C (mínima 16.2 °C y máxima 29 °C), velocidad media del viento 2.1 km–1 h y 191 mm de precipitación.
Registro y análisis de datos
Semanalmente se contabilizaron adultos, huevos y ninfas de B. cockerelli en brotes de P. ixocarpa entre las 9 h y 12 h aproximadamente. El registro de datos se realizó en dos días consecutivos: en el primero se examinaron los dos primeros lotes, y en el segundo se registraron los datos del último lote. En cada caso se realizó un muestreo sistemático para elegir al azar una planta en los primeros 3 m del primer surco seleccionado. Las observaciones se realizaron cada 3 m a lo largo del surco para elegir la planta más cercana a este punto, y así sucesivamente hasta llegar al extremo del surco. Para minimizar efectos de factores ambientales diurnos, el tiempo de muestreo se limitó a 90 min por parcela experimental y fecha de muestreo y, por tanto, el número de observaciones varió de acuerdo al desarrollo del cultivo y las condiciones físicas de los lotes experimentales. El conteo se realizó desde la primera hoja completamente desarrollada de cada brote hasta la hoja más pequeña del mismo. Esta metodología se aplicó en los tres lotes experimentales. En el lote San Martín, el de mayor extensión, el número de observaciones en las últimas fechas fue menor que en las primeras.
Los muestreos se iniciaron el 16 de abril de 2007 y en las primeras semanas se registraron densidades bajas de la plaga y únicamente en estado adulto. La densidad de todos los estados biológicos aumentó gradualmente y el análisis de la distribución espacial se realizó con los registros obtenidos el 14, 21 y 28 de mayo y 4 y 11 de junio de 2007, para los lotes X-3 y San Martín; 15, 22 y 29 de mayo y 5 y 12 de junio de 2007 para el lote Chapingo. Estas fechas coincidieron con las etapas fenológicas de plena floración e inicio de fructificación del cultivo, a 53 d después del trasplante. En el modelo de la prueba de agregación se relacionan la varianza y la media por medio de la expresión s2=axb , donde a es un factor de muestreo y b es la pendiente de la línea Log (S2) = Log (a) + b Log (x) (Southwood, 1978). Los datos se analizaron con el programa SUPRA (López, 2004) y se obtuvo la relación varianza/media (S2/ ) para cada fecha y a través de las fechas de muestreo, se obtuvieron los estimadores de los parámetros a y b, como indicador de la agregación en la Ley de la potencia de Taylor (LPT). Las estadísticas de prueba para evaluar los resultados fueron la ji-cuadrada (Ho: S2/=1) y la prueba de t (Ho: b=1).
) para cada fecha y a través de las fechas de muestreo, se obtuvieron los estimadores de los parámetros a y b, como indicador de la agregación en la Ley de la potencia de Taylor (LPT). Las estadísticas de prueba para evaluar los resultados fueron la ji-cuadrada (Ho: S2/=1) y la prueba de t (Ho: b=1).
RESULTADOS Y DISCUSIÓN
Relación varianza/media de adultos
Los adultos del lote X-3 mostraron disposición de contagio el 28 de mayo y 11 de junio con valores de la relación S2/ de 1.48 a 1.83 (Cuadro 1). Lo mismo ocurrió el 15 de mayo y 12 de junio en el lote Chapingo con valores de la relación S2/ de 1.43 a 20.6 que señalan agregación. En las otras fechas se observó un patrón aleatorio, y en el lote San Martín el patrón de disposición fue aleatorio en todos los casos (Cuadro 1).
Taylor (1984) señala que todas las formas de comportamiento de los individuos interactúan para formar verdadero contagio iniciando con la presencia de un individuo en el campo hasta que la densidad de la población aumenta de modo que afecta la presencia de otros individuos, por lo cual bajas densidades de población producen disposición aleatoria. Sin embargo, en el presente estudio, algunas de las densidades menores no produjeron resultados de aleatoriedad; por ejemplo, el 28 de mayo de 2007, del lote X-3, y el 15 de mayo, del lote Chapingo, se observaron disposiciones en agregado (Cuadro 1).
El contagio observado pudo deberse al comportamiento de la plaga en respuesta a factores del ambiente, como temperatura y precipitación, e incluso la defoliación causada por el insecto Lema trilineata (Olivier), observada durante la toma de datos en todos los lotes. Las densidades registradas el 28 de mayo del lote X-3 y la fecha 5 de junio del lote Chapingo fueron similares, pero la varianza fue superior en la fecha primeramente citada del lote X-3 donde hubo una disposición de contagio, pero en las otras fechas fueron aleatorias.
En este caso los patrones de disposición en agregado pudieron originarse porque los individuos de la plaga lograron ocupar los sitios de oviposición y apareamiento, ya que el agrupamiento de individuos se puede generar por una característica de su comportamiento, por la presencia de gradientes en la densidad de la plaga, o por un ambiente heterogéneo que propicia el agrupamiento de individuos para explotar los sitios óptimos para su supervivencia (Southwood, 1978; Soberón y Leovinsohn, 1987; Perry, 1998). Aparentemente en los lotes X-3 y Chapingo el contagio fue producido por adultos apareándose, lo cual sugiere la existencia de un mecanismo de agregación sexual.
Relación varianza/media de huevecillos y ninfas
En el caso de huevos y ninfas, las varianzas de los muestreos fueron mayores que la media, la relación S2/ fue 5 a 23.5 para huevos y 1.5 a 6.7 para ninfas, con alta significancia en todos los casos (Cuadros 2 y 3). Esto se observó en poblaciones de insectos (Vera et al., 2002) y la misma situación ocurrió en el presente estudio (Cuadros 2 y 3), porque al aumentar el número de individuos por sitio de observación, es decir la media, también aumenta la varianza. Un ejemplo, en el caso de huevos (Cuadro 2) para el lote X-3 la media fue 2.35 con una varianza de 28.52 y otra media de 11.56 con varianza de 272.30 el 21 de mayo y 11 de junio. Esta situación fue similar para las ninfas en todos los lotes, lo cual denota la existencia de un verdadero contagio en estos estados biológicos de la plaga, debido a la nula y reducida movilidad de huevos y ninfas, respectivamente. De acuerdo con los resultados de la relación S2/, los huevos y ninfas de B. cockerelli presentan una disposición de contagio en todas las fechas y en todos los lotes.
La disposición en agregado de huevos y ninfas se debe a los hábitos de oviposición de la hembra, la cual coloca huevos en grupos a lo largo del borde de las hojas o en envés. Así, al tomar los datos se contabilizaron más de 30 individuos por sitio de muestreo, y el resultado fue valores superiores de la varianza en comparación con la media, siendo la relación varianza/media mayor de 1 (Vera et al., 2002; Castillo, 2005).
Ley de la Potencia de Taylor (LPT) para adultos
Los resultados de la LPT, señalaron un patrón espacial tipo aleatorio de los adultos en los lotes X-3 y San Martín, con un valor b igual a 1.1 y 0.89 (Ho: b = 1, t=–0.698, gl=4, p=0.535), mientras que en el lote Chapingo la disposición observada fue de contagio con b igual a 2.3 (Ho: b=1, t= 5.519, gl=4 y p= 0.011) (Cuadro 4).
El grado alto de agregación ocurrido en la fecha cinco del lote Chapingo (S2/=20.56), propició gran disparidad de la varianza respecto de la media de esa misma fecha. Esta diferencia originó que el procedimiento de la LPT, en el lote Chapingo, haya detectado agregación.
Ley de la Potencia de Taylor para huevos y ninfas
Los parámetros de la LPT para huevos y ninfas señalaron diferencias en la disposición espacial de estos últimos, de acuerdo con la prueba de t donde la hipótesis nula es b=1 (Cuadro 5). Las diferencias observadas pueden estar relacionadas con las variaciones meteorológicas durante la investigación, ya que la máxima oscilación térmica diaria durante el estudio fue 20.1 °C.
Esta variación en temperatura pudo influir en el desarrollo de los individuos, pues de acuerdo con Marín (2002) el ciclo de la plaga se completa en 17 d a 29 °C promedio diario, mientras que a 16 °C promedio diario, el ciclo se concluye en 35 d, lo cual es muy cercano a la temperatura media durante la investigación (16.7 °C). También hubo rachas de viento que posiblemente influyeron en la disposición de los adultos, lo que a su vez intervino en los sitios de oviposición. Además, la precipitación fue muy intensa (191 mm) durante el registro de datos, lo que afectó especialmente la presencia de huevos y ninfas en los sitios de observación, que al estar expuestos en los brotes pudieron haber sido esparcidos por las gotas de lluvia. La aparente discordancia de la estimación en la disposición espacial entre la ley de potencia de Taylor y la relación varianza/media se debe a que, generalmente, se interpreta únicamente el parámetro b del modelo de Taylor (Taylor, 1961; Taylor et al., 1978). Sin embargo, de manera particular la agregación de individuos ocurre siempre que a>0 y b≥1, lo que ocurrió en huevos y ninfas, excepto para ninfas en la parcela Chapingo. Se puede considerar el caso general de agregación si la varianza es mayor que la media 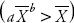 , es decir, cualquier combinación de a y b que estime una varianza mayor que la media. Por tanto, en todos las parcelas donde se hicieron muestreos de huevos y ninfas el modelo de Taylor, incluyendo ambos parámetros, indica agregación.
, es decir, cualquier combinación de a y b que estime una varianza mayor que la media. Por tanto, en todos las parcelas donde se hicieron muestreos de huevos y ninfas el modelo de Taylor, incluyendo ambos parámetros, indica agregación.
Taylor y Woiwod (1982) señalan que si σ>4 y μ>2 (se refieren a los parámetros de este estudio), los patrones de agregación se pierden y son difícilmente distinguibles. Sin embargo, en la medida que la varianza sea mayor que la media, la relación entre ambas denotará un grado alto de agregación, como ocurre con los huevecillos y ninfas.
Según Southwood (1978) la Ley de la Potencia de Taylor es un método robusto, de amplia utilización y aceptación para la descripción de la disposición espacial de las especies. A su vez, Kuno (1991) señala que puede ser bien empleada sobre un rango amplio de valores de medias.
Las diferencias en los valores del parámetro b sugieren que las condiciones ecológicas fueron cambiantes, porque incluso dentro de una misma localidad indicó diferentes patrones de disposición espacial. Debido a que no existe un índice que cubra todas las condiciones ecológicas posibles (Perry et al., 2002), es conveniente complementar el procedimiento de la LPT con otros índices, por ejemplo la relación varianza/media, para completar los resultados obtenidos, como se presentó en el presente estudio.
Debido a la variabilidad observada en los resultados se requieren más estudios para corroborar si esta variabilidad en los patrones de disposición es peculiar de B. cockerelli o se debió a los tamaños de muestra usados durante el estudio o a las densidades encontradas en el campo. Al respecto, Taylor (1984) señala que cuando en las observaciones se encuentran numerosos sitios sin individuos, las densidades son bajas o los tamaños de muestra son pequeños, la varianza tiende a ser igual a la media, por lo cual denotaría un patrón pseudoaleatorio y puede dificultar la interpretación de los valores de la LPT.
CONCLUSIONES
En las condiciones de Chapingo, México, el patrón de disposición espacial de B. cockerelli en el cultivo de tomate es variable. Mediante los valores estimados de la relación varianza/media se puede concluir que los individuos adultos de la plaga que tienen movilidad alta tienden a la aleatoriedad, mientras que los huevos y ninfas tienden a la agregación debido a los hábitos de oviposición de las hembras y al movimiento reducido de las ninfas.
LITERATURA CITADA
Almeyda, L. I. H., J. A. Sánchez, y J. A. Garzón T. 2008. Vectores causante de punta morada de la papa en Coahuila y Nuevo León, México. Agric. Tec. Mex. 34: 141-150. [ Links ]
Anónimo. 2002. Anuario estadístico de la producción agrícola de los Estados Unidos Mexicanos. INEGI. Tomo I. D. F. México. 1476 p. [ Links ]
Castillo, M. L. E. 2005. Elementos de Muestreo de Poblaciones. Universidad Autónoma Chapingo. Chapingo, México. 267 p. [ Links ]
Dajoz, R. 2002. Tratado de Ecología (2ª edición). Ed. Mundi- Prensa. Madrid. 600 p. [ Links ]
Kuno, E. 1991. Sampling and analysis of insect populations. Ann. Rev. Entomol. 36: 285-304. [ Links ]
López, C. J. 2004. Surface response program for the analysis of spatial data. Colegio de Postgraduados, Campus Veracruz. Veracruz, México. CONACYT. 15 p. [ Links ]
Marín, J. A. 2002. Características morfológicas y aspectos biológicos del psílido del tomate Bactericera cockerelli (Sulc) (=Paratrioza cockerelli). In: Memoria del taller sobre Paratrioza cockerelli (Sulc), como plaga y vector de fitoplasmas en hortalizas. Culiacán, Sinaloa. México. pp: 47-55. [ Links ]
Maron, J. L., and S. Harrison. 1997. Spatial pattern formation in an insect host-parasitoid system. Science 278: 1619-1621. [ Links ]
Perry, J. N. 1998. Measures of spatial pattern and spatial association for insect counts. In: Population and community ecology for insect management and conservation. Proceedings of the ecology and population dynamics section of the 20th international congress of entomology. Florence, Italy. pp: 21-33. [ Links ]
Perry, J. N., A. M. Liebhold, M. S. Rosenberg, J. Dungan, M. Miriti, A. Jakomulska, and S. Citron-Pousty. 2002. Illustrations and guidelines for selecting statistical methods for quantifying spatial pattern in ecological data. Ecography 25: 578-600. [ Links ]
Soberón, M. J., and M. Loevinsohn. 1987. Patterns of variations in the numbers of animal populations and the biological foundations of Taylor's law of the mean. Oikos 48: 249-252. [ Links ]
Southwood, T. R. E. 1978. Ecological Methods. Chapman and Hall. London, England. 391 p. [ Links ]
Taylor, L. R. 1961. Aggregation, variance and the mean. Nature 189: 732-735. [ Links ]
Taylor, L. R. 1984. Assessing and interpreting the spatial distributions of insect populations. Ann. Rev. Entomol. 29: 321-357. [ Links ]
Taylor, L. R., and I. P. Woiwood. 1982. Comparative synoptic dynamics. I. Relationships between inter and intra-specific spatial and temporal variance/mean population parameters. J. Anim. Ecol. 51: 879-906. [ Links ]
Taylor, L. R., I. R Woiwood, and J. N. Perry. 1978. The density-dependence of spatial behavior and the rarity of randomness. J. Anim. Ecol. 47: 383-406. [ Links ]
Tripet, R, R Christie, and A. P. Moller. 2002. The importance of host spatial distribution for parasite specialization and speciation: a comparative study of bird fleas (Syphonaptera: Ceratophyllidae). J. Anim. Ecol. 71: 735-748. [ Links ]
Vega, G. M. T., J. C. Rodríguez M., O. Díaz G, R. Bujanos M, D. Mota S, .J. L. Martínez C, A. Lagunes T., y A. Garzón, T. 2008. Susceptibilidad a insecticidas en dos poblaciones mexicanas del salerillo, Bactericera cockerelli (Sulc) (HEMIPTERA: TRIOZIDAE). Agrociencia 42: 463-471. [ Links ]

