










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.45 no.3 Texcoco abr./may. 2011
Ciencia animal
Digestibilidad de proteína, aminoácidos, fósforo, calcio y energía en dietas sorgo-pasta de soya adicionadas con fitasa en cerdos jóvenes
Digestibility of protein, amino acids, phosphorus, calcium and energy in sorghum-soybean meal diets added with phytase in pigs
Edrei Sánchez-Torres1, Miguel Barrera-Silva2, Miguel Cervantes-Ramírez1* , Adriana Morales-Trejo1, José L. Landero-Viera1, Benedicto Araiza-Piña1, Jorge Yáñez-Hernández3
1 Instituto de Ciencias Agrícolas, Universidad Autónoma de Baja California. Avenida Bernabeu # 3276, Puerta de Alcalá Res. Mexicali, Baja California, México. (miguel_cervantes@uabc.mx). *Autor responsable.
2 Departamento de Agricultura y Ganadería, Universidad de Sonora. Hermosillo, Sonora, México.
3 Escuela de Medicina Veterinaria y Zootecnia, Universidad Autónoma de Tlaxcala. Tlaxcala, México.
Recibido: Febrero, 2010.
Aprobado: Marzo, 2011.
Resumen
Los fitatos en cereales y pasta de soya (PS) interactúan con fósforo (P) y calcio (Ca) impidiendo su utilización. Estas moléculas también pueden interactuar con aminoácidos (AA), proteínas, y almidón, afectando su digestibilidad. En el presente estudio se realizó un experimento para determinar si, además de elevar la digestibilidad ileal aparente (DIA) del P y Ca, la fitasa también mejora la DIA de proteína cruda (PC) y AA, así como, la digestibilidad fecal (DF) de la energía bruta (EB). Diez cerdos castrados (18.6±1.3 kg peso inicial), canulados en íleon terminal, se asignaron a cinco dietas (tratamientos) en un Cuadro Latino repetido 5X5. Los tratamientos (T) fueron: T1) dieta testigo con sorgo-PS más P inorgánico (+PI); T2) dieta base, sorgo-PS sin P inorgánico ( —PI —FTU); T3, T4 y T5, dieta base más 250, 500 y 1000 unidades fitasa (FTU) kg—1 de alimento ( —PI+FTU). La absorción y DIA de P y Ca aumentaron, y la excreción disminuyó (p≤0.05) por la adición de fitasa a la dieta base. La excreción de P y Ca fue mayor en la dieta +PI que en la —PI —FTU o las —PI+FTU (p≤0.01). La DF de la EB aumentó (p=0.043) con fitasa, pero en menos de 1 %. La DIA de PC fue superior en las dietas — PI —FTU que en la +PI (p = 0.009), pero no se afectó por la adición de fitasa (p = 0.192). Estos resultados sugieren que la adición de fitasa a dietas sorgo-PS mejora la DIA de P y Ca, pero no cambia la de PC y AA en cerdos en crecimiento; niveles superiores a 500 FTU kg -1 de fitasa en el alimento aumentan adicionalmente la absorción de P y Ca.
Palabras clave: cerdos, fitato, fitasa, digestión, proteína.
Abstract
Phytates in grains and soybean meal (SBM) interact with phosphorus (P) and calcium (Ca), preventing their use. These molecules can also interact with amino acids (AA), proteins, and starch, affecting digestibility. In this study an experiment was conducted to determine whether in addition to raising the apparent ileal digestibility (AID) of P and Ca, phytase also improves AID of crude protein (CP) and AA, as well as fecal digestibility (FD) of gross energy (GE). Ten barrows (initial BW 18.6± 1.3 kg), fitted with cannulas at the distal ileum, were assigned to five diets (treatments) in a 5X 5 replicated Latin square design. Treatments (T) were: T1) control diet with sorghum-SBM plus inorganic P (+IP); T2) basal diet, sorghum-SBM without inorganic P ( — IP —FTU); T3, T4 and T5 were the basal diet plus 250, 500 and 1000 phytase units (FTU) kg—1 of feed ( —IP+FTU). Absorption and AID of P and Ca increased, and excretion decreased (p≤ 0.05) by phytase addition to basal diet. P and Ca excretion were higher in diet +IP than in —IP —FTU or the —IP+FTU (p≤0.01). FD of GE increased (p = 0.043) with phytase, but the increase was less than 1 %. AID of CP was higher in diets IP FTU than in + IP (p= 0.009), but was not affected by addition of phytase (p= 0.192). These results suggest that phytase addition to sorghum-SBM diets improves AID of P and Ca, but did not change that of CP and AA of growing pigs; phytase levels higher than 500 FTU kg -1 of feed additionally increased the absorption of P and Ca.
Key words: pigs, phytate, phytase, digestion, protein.
Introducción
Los cereales, especialmente maíz y sorgo, y la pasta de soya (PS), contienen fitatos, compuestos presentes naturalmente en ingredientes de origen vegetal (Maga, 1982). El ácido mio-inositol-hexafosfórico es un éster de ácido fosfórico e inositol y es la forma más común de fitatos (Reddy et al., 1982). Además del P, los fitatos interactúan con Ca, Zn, Fe y Mg formando complejos fitato-minerales, los cuales reducen la disponibilidad del P y Ca en cerdos (Cromwell, 1992). Según Reddy et al. (1982), los complejos fitato-minerales interactúan con proteínas y almidón formando complejos fitato-mineral-proteína-almidón.
La adición de fitasa microbiana a dietas para cerdos libera el P de los fitatos, aumentando su di-gestibilidad (Woyengo et al., 2008; Akinmusire y Adeola, 2009). La estrecha relación de fitatos con proteínas, por tanto, sugiere que la adición de fitasa aumentaría la digestibilidad ileal aparente (DIA) de las proteínas en cerdos. Sin embargo, el efecto de esta enzima sobre la DIA de PC y AA ha sido inconsistente. La adición de fitasa a dietas con trigo (Nortey et al, 2007) o maíz (Radcliffe et al., 2006), aumentó la DIA de AA, pero otros autores (Liao et al., 2005; Woyengo et al., 2008; Pomar et al., 2008), no encontraron efecto. Esta inconsistencia puede atribuirse en parte al tipo de ingrediente usado en las dietas experimentales.
En esos estudios las dietas experimentales se formularon principalmente con base en trigo, maíz o cebada, pero en ninguna se usó sorgo. Por tanto, el objetivo de este estudio fue evaluar el efecto de una fitasa exógena en dietas donde el sorgo es el principal cereal, y su función sobre la DIA de la PC, AA, P y Ca, y en la absorción y excreción de Ca, P y N, así como en la digestibilidad fecal (DF) de nutrientes energéticos.
Materiales y Métodos
Diez cerdos castrados (Yorkshire-Hampshire-Landrace; 18.6± 1.3 kg peso inicial), fueron canulados quirúrgicamente, con una cánula simple T en íleon terminal; aproximadamente 15 cm antes de la válvula ileocecal. Los cerdos se recuperaron de la cirugía en 7 d, lo cual se evidenció por la falta de inflamación alrededor de la incisión y un consumo estandarizado de alimento. El alojamiento fue en jaulas metabólicas individuales (1.2X0.6 m). Antes del experimento, los cerdos consumían una dieta trigo-PS, con 16.5 % PC. La fitasa usada fue de Aspergillus niger, con 5000 FTU g -1; una FTU es la cantidad de enzima que libera 1 μMol P inorgánico min 1 de una solución 5.1 mMol L—1 Fitato-Na, a pH 5.5 y 37 °C (Engelen et al., 1994).
Los tratamientos (T) fueron dietas sorgo-PS adicionadas con vitaminas, minerales, lisina, treonina y metionina cristalinas (Cuadro 1). El T1 fue una dieta testigo más P inorgánico (+PI) que cubrió todas las necesidades de nutrientes de cerdos en crecimiento (NRC, 1998). El T2 fue una dieta similar a la del T1, pero se excluyó el P inorgánico, por tanto era deficiente en P disponible y fue la dieta base (—PI — FTU). Los T3, T4 y T5 fueron la dieta base más 250, 500 y 1000 FTU kg—1 ( — PI + FTU). Las dietas se formularon a 19 % PC y contenían 0.4 % de óxido crómico para determinar la DIA de AA. La composición de AA en las dietas se muestra en el Cuadro 2.

El diseño experimental fue un Cuadro Latino 5X5 repetido (Steel et al., 1997). Cada periodo fue de 9 d: 7 para adaptación a la dieta y 2 para recolección de contenido ileal y heces. El alimento se ofreció a las 07:00 y 19:00 h, mezclado con agua en una relación 1:1. El consumo de alimento se limitó a 3.0 veces el requerimiento de ED para mantenimiento (NRC, 1998), con base en el peso vivo promedio al inicio de cada periodo. Los cerdos consumieron su ración en 15 min o menos.
El contenido ileal se recolectó durante 12 h continuas en cada día de recolección, en bolsas de plástico atadas a la cánula. La recolección se inició a las 07:00 h en los días 8 y 9. Las bolsas se retiraron de la cánula cuando se llenaron de contenido intestinal, pero no permanecieron atadas a la cánula más de 15 min. Las muestras de heces se recolectaron 3 veces en el d 8 de cada periodo. Las muestras se almacenaron a —20 °C inmediatamente después de su recolección.
Las muestras ileales y fecales se descongelaron, homogenizaron, sub-muestrearon y liofilizaron, previo al análisis en laboratorio. Éstas y las muestras de alimento se molieron a través de una malla de 1 mm y se analizaron para materia seca, P, Ca y N (AOAC, 2006). El alimento y las heces se analizaron para energía bruta en una bomba calorimétrica adiabática (IKA-Werke GMBH & Co. KG, Staufen, Germany). La composición de AA en alimento y contenido ileal se analizó en los Laboratorios de Química de la Estación Experimental de la Universidad de Missouri (método 982.30; AOAC, 2006). El óxido crómico se analizó utilizando la técnica de Hill y Anderson (1958).
Los datos se analizaron de acuerdo con el diseño indicado, usando el procedimiento GLM de SAS (1996). Las medias de tratamientos se compararon usando cuatro contrastes que evaluaron los efectos siguientes: C1, adición de P inorgánico, +PI vs. — PI —FTU; C2, P inorgánico contra fitasa, +PI vs. —PI + FTU (T3, T4 y T5); C3, adición de fitasa, —PI —FTU vs. —PI + FTU (T3, T4 y T5); y C4, efecto lineal de fitasa (T2, T3, T4 y T5). Los niveles de significancia y tendencia fueron p<0.05 y p≤0.10, para cada contraste.
Resultados
En el Cuadro 3 se presentan los datos de absorción (ABS) y excreción (EXC) de P, Ca y N; así como la DIA de P y Ca. Con respecto al P, la ABS (p = 0.001) y DIA (p = 0.024) fueron mayores en la dieta +PI que en la — PI —FTU. La ABS promedio en las dietas — PI + FTU (T3 al T5) fue menor (p = 0.001), pero la DIA no fue diferente a la +PI (p = 0.220). La fitasa aumentó linealmente la ABS (p= 0.003) y la DIA (p= 0.002) en respuesta al aumento en sus niveles. En contraste, la EXC en — PI —FTU (p = 0.010) o el promedio en las PI+ FTU (p= 0.001) fue menor que en la + PI; la fitasa redujo en forma lineal la EXC (p= 0.004).
En el caso del Ca, la ABS en la dieta + PI fue mayor que en PI FTU y en el promedio de las dietas —PI + FTU (p = 0.001); en estas últimas la ABS fue mayor que en la PI FTU (p= 0.043). La EXC fue mayor en + PI que en PI FTU (p = 0.008) y en las —PI + FTU (p = 0.001); en estas últimas este valor fue menor (p= 0.041) que en la — PI —FTU. La DIA en la dieta +PI no fue diferente a la de PI FTU (p= 0.466) o al promedio de las — PI + FTU (p = 0.178); pero en éstas fue mayor que en la PI FTU (p= 0.034). No se observó tendencia lineal para ABS (p = 0.158), EXC (p = 0.109) o DIA (p= 0.200) de Ca en respuesta al aumento en fitasa.
La ABS de N tendió a ser mayor (p= 0.090) y la EXC fue menor (p= 0.043) en las dietas sin P inorgánico que en la + PI. Asimismo, la EXC tendió a ser menor (p= 0.066) en las dietas con fitasa, comparadas con la + PI. No hubo efecto de fitasa en la ABS y EXC de N, en comparación con la dieta base.
En el Cuadro 4 se presentan los resultados de la DIA de PC y AA, y la DF de EB. La DF de EB en la dieta + PI fue mayor que en la — PI — FTU (p = 0.001) o el promedio de las —PI + FTU (p = 0.022). Este valor fue mayor que en la dieta PI FTU (p= 0.043), pero no se observó tendencia lineal (p= 0.285). La DIA de PC fue superior en la dieta PI FTU (p = 0.009) y en las —PI + FTU (p = 0.034) que en la + PI, pero no hubo efecto de fitasa en la dieta base (p = 0.192). La DIA de los AA no fue diferente (p≥0.10) entre las dietas +PI y — PI —FTU; tampoco hubo diferencia entre las dietas PI+ FTU y + PI (p> 0.10). En promedio, la fitasa tampoco afectó la DIA de AA no esenciales.
Discusión
Fitasa se incluye en dietas sorgo- o maíz-PS para cerdos con dos propósitos: liberar el P de los fitatos y reducir o eliminar el uso de fuentes inorgánicas de P. Con esto se busca también disminuir la EXC de P en las excretas y el riesgo de contaminar suelos y agua con P. Lo anterior se debe a que la mayor parte del P en los cereales y en la PS está ligada al ácido mioinositol-hexafosfórico o fitato, donde no está disponible debido a que los cerdos no producen fitasa (Lei y Stahl, 2001). De acuerdo con Cromwell (1992), sólo 10 a 15 % del P en maíz y sorgo, y de 25 a 30 % en la PS está disponible para cerdos.
La eliminación de P inorgánico de dietas formuladas con sorgo-PS para cerdos, reduce la EXC de P y Ca en las heces. En la Figura 1 se presentan los porcentajes en que se redujo la EXC debido a la eliminación de P inorgánico y la adición de fitasa a la dieta; a la dieta testigo se le dio un valor arbitrario de 100. La EXC de P y Ca se reduce en 20 y 22 % por la eliminación de fosfato dicálcico de la dieta. Esta eliminación combinada con la adición de fitasa redujo en 43 y 35 %, en promedio, la EXC de de P y Ca. El análisis de estos datos muestra que la fitasa en la dieta PI puede ayudar a cubrir el requerimiento de P disponible para cerdos en crecimiento. Estos resultados coinciden con los obtenidos cuando se usan dietas elaboradas con trigo, maíz o cebada (Liao et al., 2005; 2007).

En la Figura 2 se presentan los porcentajes en que se incrementó la ABS de P (37 a 45 %) y Ca (10 a 20 %) debido a la adición de fitasa a la dieta base. La dieta base contenía 0.16 % de P disponible y la adición de 500 FTU lo elevó a 0.23 %, el cual es semejante al requerimiento de cerdos con pesos de 20 a 50 kg (NRC, 1998). El análisis de estos datos indica que la fitasa puede sustituir al P inorgánico en dietas sorgo- o maíz-PS, sin afectar el suministro de P disponible requerido, coincidiendo con lo publicado por Liao et al. (2005, 2007). El comportamiento lineal en la ABS y EXC de P en respuesta a la adición de fitasa (Cuadro 3), indica que niveles superiores a las 500 FTU kg -1 recomendadas, adicionalmente incrementan la ABS y reducen la EXC de P. Sin embargo, es importante enfatizar que con 500 FTU kg -1 de alimento aumenta la disponibilidad de P a niveles semejantes a los requeridos por cerdos con peso mayor a 20 kg.
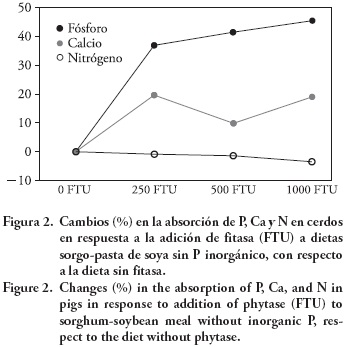
La falta de efecto de fitasa en la EXC y la ABS de N (Figura 1 y 2) coincide con lo observado en la DIA de PC y AA y con lo publicado por otros autores. Traylor et al. (2001) y Woyengo et al. (2008) no encontraron efecto de fitasa en pasta de soya; Pomar et al. (2008) en dietas elaboradas con trigo, maíz y pasta de soya; Yáñez et al. (2011) en residuos secos de destilería. Liao et al. (2005) tampoco observaron efecto de fitasa en la DIA de AA en dietas trigo-PS-pasta de canola, y sugieren que el efecto de la fitasa en la DIA de AA depende de la composición de la dieta. La interacción de fitatos con proteínas y AA de la dieta y con enzimas pancreáticas e intestinales podría explicar la falta de respuesta en esta variable (Reddy et al., 1982). En este sentido, estudios in vitro sugieren que los fitatos pueden formar complejos con proteína y AA libres en el tubo digestivo de cerdos. Honig y Wolf (1991) encontraron complejos fitato-proteína en concentrados proteínicos de soya a pH 8. Esta interacción puede provocar cambios en la estructura de la proteína, afectar la actividad enzimática y la digestibilidad de la proteína (Greiner y Konietzny, 2006). Reddy et al. (1982) señalan que los fitatos podrían inhibir la actividad enzimática mediante la interacción directa o a través de cationes necesarios para su activación. Con base en estos hallazgos, se esperaría que la fitasa liberara a las proteínas y AA de los fitatos e incrementara su DIA. Sin embargo, los resultados de éste y otros estudios in vivo (Liao et al., 2005, 2007; Woyengo et al., 2008; Yáñez et al., 2011), no coinciden con los realizados in vitro.
La explicación de esta discrepancia es compleja y debe incluir el análisis de aspectos bioquímicos y físico-químicos asociados a la digestión de las proteínas. Una explicación parcial es que los complejos proteína-AA-fitatos se estabilizan por interacciones electrostáticas débiles (Láztity y Láztity, 1995), las cuales pueden romperse con los cambios normales de pH en el intestino delgado de cerdos, sin necesidad de adicionar ninguna enzima específica a la dieta. Okubo et al. (1976) encontraron que la capacidad máxima de unión de fitatos con glicinina de PS era a pH 2.5, pero no hubo ninguna unión al punto isoeléctrico de la proteína (pH 4.9). Desphande y Damodaran (1989) observaron interacciones de fitatos con tripsina y quimotripsina a pH 3.0, pero ninguna a pH 7.8. A este pH, proteínas y fitatos poseen cargas negativas que impiden su interacción. Aunque esos autores reportan una reducción en la actividad de tripsina a pH ácido, éste no ocurre en el intestino delgado de cerdos (pH 6.2 a 7.4; Jong-bloed et al., 1992). Rawlings y Barret (1994) indican que en el sitio activo de estas enzimas radican residuos de serina, histidina y aspartato; ninguno de estos interactúa con fitatos a pH neutro o alcalino en la ausencia de Ca2+. Además, la cantidad de fitatos que permanecen libres y podrían inhibir las enzimas digestivas es muy baja y de escaso impacto debido a la fuerte afinidad de los fitatos por los cationes al pH intestinal (Desphande y Damodaran, 1989). Por tanto, el efecto de adicionar fitasa a dietas sorgo-pasta de soya en la DIA de PC y AA parece ser insignificante.
La adición de fitasa incrementó menos de 1 % la DF de la EB. Este aumento coincide con lo publicado por Liao et al. (2005) en dietas trigo-PS-pasta de canola (0.63 %); sin embargo, no se observó ningún efecto en estudios realizados por Liao et al. (2005) en dietas maíz-PS, trigo-PS o cebada-pasta de canola; Woyengo et al. (2008) en dietas base trigo; Yáñez et al. (2011) en residuos secos de destilería. La reducida magnitud del efecto de fitasa en la DF de EB se puede explicar de manera similar a lo explicado con respecto a la DIA de AA.
Conclusiones
La adición de fitasa a dietas sorgo-pasta de soya aumenta la absorción y la digestibilidad, y reduce la excreción de P y Ca, pero no afecta la digestibilidad ileal aparente de proteína y aminoácidos en cerdos en crecimiento. Niveles superiores a 500 FTU kg -1 aumentan la absorción de P y Ca. Aunque la fitasa incrementó la digestibilidad de nutrientes energéticos, la magnitud del incremento tiene un impacto mínimo en la alimentación de los cerdos.
Literatura Citada
Akinmusire, A. S., and O. Adeola. 2009. True digestibility of phosphorus in canola and soybean meals for growing pigs: Influence of microbial phytase. J. Anim. Sci. 87: 977-983. [ Links ]
AOAC. 2006. Official Methods of Analysis of AOAC. 18th ed. Association of Official Analytical Chemists. Gaithersburg, MD, USA. 2000 p. [ Links ]
Cromwell, G. L. 1992. The biological availability of phosphorous in feedstuffs for pigs. Pig News and Information 13: 75N-78N. [ Links ]
Desphande, S. S., and S. Damodaran. 1989. Effect of phytate on solubility, activity and conformation of trypsin and chymotrypsin. J. Food Sci. 54: 695-699. [ Links ]
Engelen, A. J., F. C. van der Heeft, P. H. G. Ransdorp, and E. L. C. Smit. 1994. Simple and rapid determination of phytase activity. J. AOAC Int. 77: 760-764. [ Links ]
Greiner, R., and U. Konietzny. 2006. Phytase for food application. Food Technol. Biotech. 44: 125-140. [ Links ]
Hill, F. N., and D. L. Anderson. 1958. Comparison of metabolizable energy and productive energy determination with growing pigs. J. Nutr. 64: 587-603. [ Links ]
Honig, D. H., and W. J. Wolf. 1991. Phytate-mineral-protein composition of soybeans: gel filtration studies of soybean meal extracts. J. Agric. Food Chem. 39: 1037-1042. [ Links ]
Jongbloed, A. W., Z. Mroz, and P. A. Kemme. 1992. The effect of supplementary Aspergillus niger phytase in diets for pigs on concentration and apparent digestibility of dry matter, total phosphorus, and phytic acid in different sections of the alimentary tract. J. Anim. Sci. 70: 1159-1168. [ Links ]
Láztity, R., and L. Láztity. 1995. Phytic acid in cereal technology. In: Pomeranz, Y. (ed). Advances in Cereal Science and Technology. American Association of Cereal Chemists. pp: 309-372. [ Links ]
Lei, X. G., and C. H. Stahl. 2001. Biotechnological development of effective phytases for mineral nutrition and environmental protection. Appl. Microbiol. Biotechnol. 57: 474-481. [ Links ]
Liao, S. F., A. K. Kies, W. C. Sauer, Y. C. Zhang, M. Cervantes, and J. M. He. 2005. Effect of phytase supplementation to a low- and a high- phytate diet for growing pigs on the digestibilities of crude protein, amino acids and energy. J. Anim. Sci. 83: 2130-2136. [ Links ]
Liao, S. F., W. C. Sauer, J. K. Htoo, M. Cervantes, A. K. Kies, B. A. Araiza, and A. Morales. 2007. Effect of phytase supplementation to a high- and a low-phytate diet for growing pigs on the utilization of phosphorus and calcium. Interciencia 32: 195-201. [ Links ]
Maga, J. A. 1982. Phytate: Its chemistry, occurrence, food interactions, nutritional significance, and methods of analysis. J. Agric. Food Chem. 30: 1-9. [ Links ]
Nortey, N., J. F. Patience, P. H. Simmins, N. L. Trottier, and R. T. Zijlstra. 2007. Effects of individual or combined xylanase and phytase supplementation on energy, amino acid, and phosphorus digestibility and growth performance of grower pigs fed wheat-based diets containing wheat millrun. J. Anim. Sci. 85: 1432-1443. [ Links ]
NRC. 1998. Nutrient Requirements of Swine. 10th ed. National Academy Press. Washington, DC, USA. 183 p. [ Links ]
Okubo, K., D. V. Meyers, and G .A. Iacobucci. 1976. Binding of phytic acid to glycinin. Cereal Chem. 53: 513-524. [ Links ]
Pomar, C., F. Gagné, J. J. Matte, G. Barnett, and C. Jondreville. 2008. The effect of microbial phytase on true and apparent ileal amino acid digestibilities in growing-finishing pigs. J. Anim. Sci. 86:1598-1608. [ Links ]
Radcliffe, J.S., R. S. Pleasant, and E. T. Kornegay. 2006. Estimating equivalency values of microbial phytase for amino acids in growing and finishing pigs fitted with steered ileo-cecal valve cannulas. J. Anim. Sci. 84: 1119-1129. [ Links ]
Rawlings, N. D., and A. J. Barrett. 1994. Families of serine peptidases. Meth. Enzymol. 244: 19-61. [ Links ]
Reddy, N. R., S. K. Sathé, and D. K. Salunkhe. 1982. Phytates in legumes and cereals. Adv. Food Res. 28: 1-92. [ Links ]
SAS (Statistical Analysis System). 1996. SAS/STAT User's Guide (Release 6.12.). SAS Institute Inc., Cary, N. C. 1028 p. [ Links ]
Steel, D. R. G., J. H. Torrie, and D. A. Dickey. 1997. Principles and Procedures of Statistics: a Biomedical Approach (3rd ed.). McGraw-Hill Book Co., New York. 622 p. [ Links ]
Traylor, S. L., G. L. Cromwell, M. D. Lindeman, and D. A. Knabe. 2001. Effects of level of supplemental phytase on ileal digestibility of amino acids, calcium, and phosphorus in dehulled soybean meal for growing pigs. J. Anim. Sci. 79: 2634-2642. [ Links ]
Woyengo, T. A, J. S. Sands, W. Guenter, and C. M. Nyachoti. 2008. Nutrient digestibility and performance responses of growing pigs fed phytase- and xylanase-supplemented wheat-based diets. J Anim Sci 86: 848-857. [ Links ]
Yáñez, J. L., E. Beltranena, M. Cervantes, and R. T. Zijlstra. 2011. Effect of phytase and xylanase supplementation or particle size on nutrient digestibility of diets containing distillers dried grains with solubles (DDGS) co-fermented from wheat and corn in ileal-cannulated grower pigs. J. Anim. Sci. 89: 113-123. [ Links ]

