










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.44 no.7 Texcoco oct./nov. 2010
Fitociencia
Variación en la fenología de la floración y en el éxito reproductivo en Setaria lachnea
Variations in flowering phenology and reproductive success in Setaria lachnea
Eliana Exner, Juan M. Zabala, José F. Pensiero
Universidad Nacional del Litoral, Facultad de Ciencias Agrarias, Departamento de Biología Vegetal, 86–Kreder 2805 (S3080HOF) Esperanza, Santa Fe. Argentina. Autor responsable: (eexner@fca.unl. edu.ar), (jmzabala@fca.unl.edu.ar), (jfpensi@fca.unl.edu.ar).
Recibido: Noviembre, 2009.
Aprobado: Agosto, 2010.
Resumen
Setaria lachnea, moha petenne, es una gramínea nativa con excelentes cualidades forrajeras por lo que se recomienda su introducción al cultivo. Por tanto, en el presente estudio se analizó la variabilidad en la fenología de la floración y en el éxito reproductivo en genotipos de tres poblaciones argentinas, provenientes de distintos lugares por gradiente altitudinal, como aspectos de interés al seleccionar materiales. El diseño experimental fue completamente al azar, con 10 genotipos por población y 3 repeticiones por genotipo. Se analizó la fenología de la floración con las variables inicio de floración y mitad de floración; se cuantificó el número de panojas por observación, el número de panojas totales y el éxito reproductivo con el porcentaje de fructificación. Para el inicio de floración sólo se hallaron variaciones significativas (p≤0.05) intrapoblacionales. El 75 % de los genotipos inició su floración en noviembre del 2005. Pata la mitad de floración hubo diferencias significativas (p≤0.05) inter e intrapoblacionales. La variabilidad intrapoblacional encontrada permitirá seleccionar genotipos con un periodo vegetativo prolongado. El éxito reproductivo medido como porcentaje de fructificación promedio fue 41 % (alto para una especie no domesticada), con una variación de 3 % a 80 %. En los dos años (2005–2006) analizados, se hallaron diferencias significativas (p≤0.05) inter e intrapoblacionales. Los genotipos de la población de menor altura se destacaron por su alto porcentaje de fructificación, siempre mayot a 40 %. Sin embargo, no hubo un patrón de variación asociado con la altitud. El inicio de floración no se correlacionó con el porcentaje de fructificación, por lo cual se podrá seleccionar materiales tardíos sin alterar el porcentaje de fructificación.
Palabras claves: forrajera nativa, gradiente altitudinal, moha perenne, variabilidad interpoblacional, variabilidad intrapoblacional.
Abstract
Setaria lachnea, moha petenne, is a native gtass with excellent fotage quality; thus, its inttoduction to cultivation has been tecommended. Thetefote, in the ptesent study, the variability in flowering phenology and teptoductive success of thtee Atgentine populations from diffetent places by altitudinal gtadient have been analyzed, with respect to points of interest fot selecting materials. A completely randomized experimental design was used, with 10 genotypes per population and 3 replications per genotype. Flowering phenology was analyzed through variables of flowering start and mid–flowering; the number of panicles was quantified by observation, the number of the total of panicles and reproductive success by fruiting percentage. For flowering start only significant (p≤0.05) intrapopulation variations wete found. Seventy–five percent of the genotypes started flowering in November 2005. For mid–flowering, there wete significant inter– and intra–population differences (p≤0.05). The intrapopulation variability obsetved will allow selecting genotypes with ptolonged vegetative period. Reptoduction success, measured as mean fruiting percentage, was 41 (high fot a nondomesticated species), with variation from 3 % to 80 %. In the two analyzed yeats (2005–2006), significant inter– and intra–population differences (p≤0.05) wete found. Genotypes of the population of less height stood out by their high fruiting percentage, always above 40 %; however, there was no variation pattern associated to altitude. Beginning of flowering was not correlated with fruiting percentage; therefore, it will be possible to select late–ripening materials, without altering fruiting percentage.
Key wotds: native forage, altitudinal gradient, moha perenne, interpopulation variability, intrapopulation variability.
INTRODUCCIÓN
Setaria lachnea (Nees) Kunth, moha perenne, es una gramínea nativa, perenne, frecuente en el centro y norte de Argentina, Paraguay, Uruguay, Bolivia, Perú, Venezuela, y rara en Brasil. Presenta un ciclo primavera–estivo–otoñal y se destaca por su producción y calidad forrajera, por lo que su introducción al cultivo ha sido recomendada (Covas y Frecentese, 1983). Su distribución es amplia en Argentina, desde el nivel del mar hasta 2250 m, en sitios con precipitaciones de 450 a 1300 mm anuales y en una amplia variedad de suelos, aunque no tolera los salinos (Pensiero, 1999).
Entre los aspectos reproductivos más importantes para la domesticación de una especie forrajera se destacan la concentración de la floración y un aumento del éxito reproductivo (Jain, 1975). El tiempo de floración puede afectar los mecanismos de polinización, la producción y el éxito reproductivo (Wright y Meagher, 2003; Stinson, 2004; Laceyetal., 2003). El conocimiento fenológico de la floración de una especie de interés forrajero permite predecir la máxima producción en una estación de crecimiento (Slafer, 2003). El inicio de floración produce la disminución de la calidad forrajera debido a una reducción de la relación hoja/tallo y de la digestibilidad de hoja (Bruinenberg et al., 2002). Setaria lachnea presenta forraje de buena calidad, con un contenido de proteína superior al 15 %, hasta el inicio de su floración (Covas y Frecentese, 1983). Dicha calidad disminuye al estado reproductivo, en el cual Guaita et al. (1989) hallaron 5.85 % de proteína en láminas. El inicio del ciclo reproductivo es un carácter altamente heredable en poblaciones naturales (Elzinga et al., 2007) importante en programas de mejoramiento de gramíneas forrajeras (Casler, 1998).
El éxito reproductivo es un factor determinante de la producción de semillas en especies forrajeras no domesticadas (Fang et al., 2004), en las que es necesario seleccionar materiales que combinen cantidad y calidad de forraje con una producción rentable de semillas (Cole y Johnston, 2006). Respecto a las características reproductivas de S. lachnea, Pensiero et al. (2005) señalan la ausencia de mecanismos de autoincompatibilidad y postularon a la autogamia como el sistema reproductivo dominante. Los porcentajes de fructificación hallados por Pensiero et al. (2005) para cinco poblaciones de S. lachnea fueron altos (67 % en algunas poblaciones) para una especie no domesticada. Las poblaciones de numerosas especies que corresponden a un gradiente altitudinal suelen presentar marcadas diferencias en la fenología de su floración y fructificación (Jordano y Godoy, 2000; Yamagishi et al., 2005). Esto ocurre más frecuentemente en especies herbáceas (Ohsawa y Ide, 2008; Vigouroux et al, 2003).
La hipótesis fue que existe variabilidad (inter e intrapoblacional) en el inicio de la etapa reproductiva y en el éxito reproductivo de S. lachnea. El objetivo de este estudio fue analizar la variación (inter e intrapoblacional) en la fenología de la floración y en el éxito reproductivo en tres poblaciones provenientes de un gradiente altitudinal.
MATERIALES Y MÉTODOS
Los estudios se realizaron en 29 genotipos de tres poblaciones argentinas de 5. lachnea procedentes de: 1) población 6221, provincia Santiago del Estero, departamento Aguirre, Malbrán (29° 21' S, 62° 27' O, 81 m); 2) población 6227, provincia Tucumán, departamento Trancas, Vipos (26° 29' S, 65° 21' O, 795 m); 3) población 6242, provincia Salta, departamento Chicoana, Escoipe (25° 06' S, 65° 38' O, 2250 m). Las claves 6221, 6227 y 6242 corresponden al número de colección de los ejemplares de referencia de cada población, depositados en el Herbario SF (FCA–UNL). Para cada población se recolectaron semillas mediante cosechas de plantas individuales, las cuales se conservaron en bolsas de papel y mantuvieron a temperatura ambiente (15–25 °C) hasta el momento de su siembra. La siembra de cada genotipo se realizó en bandejas individuales con una mezcla de tierra y arena (v/v 1:1). Después las plántulas se transplantaron en el campo experimental de la Facultad de Ciencias Agrarias en Esperanza, provincia de Santa Fe, Argentina (31° 27' S, 60° 56' O), donde 10 genotipos de las poblaciones 6227 y 6221, y nueve de la 6242 fueron dispuestos en un diseño completamente aleatorizado. Cada genotipo estuvo representado por tres clones o repeticiones (unidades experimentales). Las 87 plantas estuvieron distanciadas a 0.50 m entre sí
Fenología de la floración
Para cada clon se registró la fecha de emergencia de panojas y el número de panojas totalmente emergidas en cada observación, sin importar si la panoja correspondía a un eje principal o a ramificaciones del mismo. Se realizaron tres recuentos semanales desde el 30 de octubre de 2005 hasta el 31 de diciembre de 2005 y un recuento semanal desde el 1 de enero de 2006 hasta el 3 de abril de 2006. Las panojas ya contabilizadas se marcaron envolviendo sus pedúnculos con cinta.
Según el criterio propuesto por Quinn y Wetherington (2002) y Fang et al. (2004), la fecha en que cada clon presentó la primera panoja totalmente emergida de la hoja bandera se consideró como inició de floración. La mitad de la floración fue la fecha en que cada clon alcanzó la mitad de panojas totales producidas. Ambas variables se expresan en días desde el 30 de octubre de 2005.
La variable panojas totales de cada clon corresponde al número total de panojas producidas. La curva de floración de cada genotipo (promedio de sus tres clones) se gráfico con el número de panojas emergidas por recuento.
Éxito reproductivo
El éxito reproductivo de los genotipos se evaluó para las cosechas 2005 y 2006. Las espiguillas de cada clon se cosecharon individualmente a medida que se observaba la madurez de las mismas. Las panojas se trillaron a mano y almacenaron en bolsas de papel. Para cada clon se analizó una muestra tomada al azar de 50 espiguillas (n=150 por genotipo), por lo que las poblaciones 6227 y 6221 estuvieron representadas por 1500 espiguillas cada una y la población 6242 por 1350 espiguillas.
El éxito reproductivo se expresó como el porcentaje de las flores que produjeron fruto (cariopsis) usando la variable porcentaje de fructificación; para ello y con microscopio estereoscópico se observó la presencia o ausencia de cariopsis en las espiguillas. En el género Setaria, como en la mayoría de las Paniceae, las espiguillas poseen dos antecios, uno basal, reducido y asexuado, y otro apical, con flor hermafrodita y fructífero.
Análisis estadístico
Los datos obtenidos se analizaron mediante análisis de la varianza anidado, previo análisis de normalidad y homogeneidad de varianzas; las medias se compararon con la prueba de Tukey (p≤0.05). El efecto de los genotipos anidados en poblaciones se consideró aleatorio y se usó como fuente de variación para analizar el efecto del factor población. Se correlacionaron los datos de fenología de floración y éxito reproductivo con la correlación de Pearson (p≤0.05). Para cada análisis se usó el programa InfoStat (versión 2006).
RESULTADOS Y DISCUSIÓN
Fenología de la floración
Para la variable inicio de floración no hubo diferencias significativas entre poblaciones (p=0.66), pero hubo variaciones significativas intrapoblacionales (p≤0.0001) (Figura 1). El 4 % de los genotipos iniciaron su floración a fines de octubre, 75 % en noviembre, 17 % en diciembre y 4 % en enero. La población 6227 presentó un inicio de floración extendido en el tiempo, con 84 d entre el primero y el último genotipo en mostrar al menos una panoja totalmente emergida; la población 6242 tuvo un inicio de floración más concentrado (16 d); y la población 6221 presentó un inicio de floración a los 27 d.
Hubo diferencias significativas entre poblaciones (p=0.002) y dentro de poblaciones (p≤0.0001) para la variable mitad de floración (Figura 2). El 28.87 % de la variación fue explicada por diferencias interpoblacionales y el 50.67 % por diferencias intrapoblacionales.
Un aspecto interesante fue la variabilidad intrapoblacional encontrada en los caracteres fenológicos, ya que en especies autógamas se espera que las diferencias interpoblacionales en caracteres adaptativos sean de mayor magnitud que las intrapoblacionales (Ramakrishnan et al., 2004).
Respecto al gradiente altitudinal, la población proveniente de altura intermedia (6227) presentó la mayor variación en el inicio de floración; pero las poblaciones provenientes de mayor y menor altura (6242 y 6221) mostraron menor variación. La variación observada en la población de mayor altura podría deberse a que provienen de ambientes con un corto período favorable de crecimiento, que las plantas aprovechan para asegurar su éxito reproductivo. Quinn y Wetherington (2002) muestran resultados similares para Sporobolus cryptandrus y Panicum virgatum. En ambientes con un mayor período de condiciones óptimas para el crecimiento, la diversificación de la fenología podría favorecer el éxito reproductivo.
En relación con la domesticación de esta especie, su variabilidad permitirá seleccionar genotipos con un período vegetativo más prolongado, seleccionando aquellos con un inicio de floración más tardío, lo que permitiría extender la producción de forraje de mejor calidad (Guaita et al., 1989; Pontes et al., 2007).
En la variable panojas totales hubo variaciones interpoblacionales (p=0.0028) e intrapoblacionales (p≤0.0001) (Figura 3). El 27.53 % de esta variación se explica en términos de diferencias interpoblacionales y el 48.27 % por diferencias intrapoblacionales.
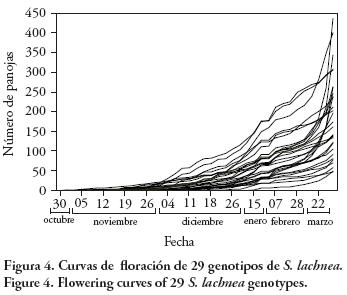
Las curvas de floración de los genotipos de las tres poblaciones muestran una tendencia ascendente ya que la producción de panojas aumenta hasta las primeras heladas, momento en el que la parte aérea de la planta se seca para luego rebrotar en la próxima primavera (Figura 4). En el otoño la floración se produce, en gran parte, desde ramificaciones axilares. Estas continúan iniciándose mientras haya condiciones favorables para el crecimiento de las plantas, permitiendo una floración y producción de semillas continua (Dekker, 2004), lo que podría considerarse como una estrategia para asegurar el éxito reproductivo (Doust y Kellogg 2006). Este tipo de arquitectura difiere de la típica encontrada en las especies domesticadas que poseen pocos macollos y sin ramificaciones axilares con inflorescencias (Harían, 1992; Doust y Kellogg, 2006). En futuros trabajos será necesario evaluar el efecto de las ramificaciones axilares en la producción de semilla de esta especie. En especies de Setaria domesticadas, el acortamiento de la fenología de la floración ha llevado a la pérdida de ramificaciones axilares, característica importante para el manejo y la cosecha (Harían, 1992).
Éxito reproductivo
El porcentaje de fructificación promedio fue 41 %, con una variación de 3 a 80 %. En los dos años evaluados hubo diferencias consistentes y significativas interpoblacionales e intrapoblacionales (p≤0.0001) (Cuadro 1; Figura 5). El 45 % de la variación puede explicarse a nivel poblacional y el 21 % a nivel de genotipos. Los genotipos de la población 6221 se destacaron por su elevado porcentaje de fructificación, superior en todos los casos al 40 %.

Los resultados obtenidos indican la existencia de variabilidad interpoblacional e intrapoblacional para el carácter porcentaje de fructificación, aspecto importante para seleccionar materiales de interés agronómico. Además, estos valores son altos por tratarse de una especie no domesticada. Pensiero et al. (2005), para cinco poblaciones de S. lachnea, hallaron en algunas un promedio de 67 % de fructificación, coincidiendo con los valores de fructificación obtenidos aquí para la población 6221. Se han registrado variaciones importantes en la producción de semillas de distintas especies del género Setaria (Steel et al., 1983). Haar (1998) considera que en este género la producción de semillas es muy plástica y fuertemente dependiente de la acumulación de biomasa en la planta, nivel de macollaje y de la arquitectura de sus inflorescencias. Clark y Pohl (1996), para Setaria faberi y S. viridis, reportan que el número de espiguillas fértiles por panoja es variable y plástico en respuesta a las condiciones ambientales. Según Covas y Frecentese (1983), en S. lachnea si bien hay pérdida de semillas debido a la caída de espiguillas con sus cariopsis maduros, la misma no resultaría significativa en relación al total de semillas retenidas en la panoja.
Correlación entre los caracteres evaluados
El análisis de correlación por población mostró, para la población 6221, una correlación negativa y significativa (–0.64, p≤0.001) entre inicio de floración y panojas totales. Esto es lógico debido a la longitud del período reproductivo mostrado por los genotipos analizados. El porcentaje de fructificación no se correlacionó con ninguna de las otras variables.
La población 6227 presentó también una correlación negativa y significativa (–0.70; p≤0.001) entre inicio de floración y panojas totales. A diferencia de la anterior, en esta población la correlación fue positiva y significativa (0.43; p=0.02) entre mitad de floración y porcentaje de fructificación. Los genotipos de esta población, que más tarda en llegar a mitad de floración, producen gran cantidad de panojas que podría compensar su menor porcentaje de fructificación.
No se hallaron correlaciones significativas para la población 6242.
Debido a que el inicio de floración no está correlacionado con el porcentaje de fructificación, podrían seleccionarse materiales más tardíos sin afectar el porcentaje de fructificación.
En el género Setaria se ha mostrado una correlación negativa entre extensión del período reproductivo y producción de semilla. Hacker y Cuany (1997) indican que en S. sphacelata una floración extendida provocó falta de uniformidad en la aparición de panojas, menor retención de frutos y, por consiguiente, una menor producción de semillas.
Hubo una correlación positiva significativa (0.87; p≤ 0.001) entre los porcentaje de fructificación de los dos años estudiados, manteniéndose el comportamiento de las poblaciones. Esto sugiere que una parte importante de la variabilidad observada en este carácter sería de origen genético y por ello factible de mejorar por selección.
CONCLUSIONES
La variabilidad hallada y la falta de correlación entre inicio de floración y porcentaje de fructificación permitirán seleccionar materiales con un período vegetativo más prolongado, sin afectar el porcentaje de fructificación.
Setaria lachnea posee una notable variabilidad intrapoblacional para los caracteres inicio de floración, mitad de floración, panojas totales y porcentaje de fructificación, lo cual es poco común en especies autógamas.
El porcentaje de fructificación promedio es alto por tratarse de una especie no domesticada.
Para los caracteres analizados Setaria lachnea no presenta patrones de variación asociadas con la altitud.
LITERATURA CITADA
Bruinenberg, M. H., and H. Valk. Korevaar and P.C.Struik. 2002. Factors affecting digestibility of temperate forages from seminatural grasslands: a review. Grass Forage Sci. 57: 292–301. [ Links ]
Casler, M. D. 1998. Genetic variation within eight populations of perennial forage grasses. Plant Breeding 117: 243–249. [ Links ]
Clark, L. G., and R.W. Pohl. 1996. Agnes Chases First Book of Grasses. Smithsonian Institution Press. Washington and London: 4th ed. 127 p. [ Links ]
Cole, I. A., and W. H. Johnston. 2006. Seed production of Australian native grass cultivars: an overview of current information and future research needs. Austr. J. Exp. Agrie. 46:361–373. [ Links ]
Covas, G., y M. Frecentese. 1983. Setaria leiantha Hackel, un pasto nativo para integrar pasturas perennes en la región semiárida. Agrarius 1:16–17. [ Links ]
Dekker, J. 2004. Evolutionary biology of the foxtail (Setaria) species–group. In: Inderjit, ed. Weed Biology. Management. Kluwer Academic Publishers, The Netherlands, pp: 65–114. [ Links ]
Doust, A. N., and E. A. Kellogg. 2006. Effect of genotype and environment on branching in weedy green millet (Setaria viridis) and domesticated foxtail millet (Setaria itálica) (Poaceae). Molecular Ecol. 15:1335–1349. [ Links ]
Elzinga, J. A., A. Atlan, A. Biere, L. Gigord, A. E. Weis, and G. Bernasconi. 2007. Time after time: flowering phenology and biotic interactions. Trends Ecol. Evol. 22: 432–439. [ Links ]
Fang, C, T. S. Aamlid, O. Jorgensen, and O. A. Rognli. 2004. Phenotypic and genotypic variation in seed production traits within a full–sib family of meadow fescue. Plant Breeding 123:241–246. [ Links ]
Fang, X., P. K. Subudhi, B. C. Venuto, S. A. Harrison, and A. B. Ryan. 2004. Influence of flowering phenology on seed production in smooth cordgrass (Spartina alterniflora Loisel.). Aquatic Bot. 80:139–151. [ Links ]
Guaita, M. S., H. H. Fernández, e I. N. Tiranti. 1989. Calidad forrajera de Setaria leiantha en estado reproductivo. Rev. Univ. Nac. Río Cuarto 9:105–108. [ Links ]
Hacker, J. B., and R. L. Cuany. 1997. Genetic variation in seed production and its components in four cultivars of the pasture grass Setaria sphacelata. Euphytica 93:271–282. [ Links ]
Harlan, J. R. 1992. Crops and Man. In: 2nd edn. Am. Soc. Agron. and Crop Sci. Soc. Am.(eds). Madison, Wisconsin. 268 p. [ Links ]
Haar, J. M. 1998. Characterization of foxtail (Setaria spp.) seed production and giant foxtail (Setaria faberii) seed dormancy at abscission. Ph. D. dissertation. Iowa State University, Ames, Iowa, USA. 122 p. [ Links ]
INFOSTAT 2006. Versión p.2. Software estadístico InfoStat. Grupo Infostat. Estadística y Biometría. Facultad de Ciencias Agropecuarias. Universidad Nacional de Córdoba (FCA. UNC). CD. [ Links ]
Jain, S. K. 1975. Population structure and the effects of breeding system. In: Frankel, O. H., and J. G. Hawkes (eds). Crop Genetic Resources for Today and Tomorrow. International Biological Programme Synthesis Series 2. Cambridge, Great Britain, pp: 15–36. [ Links ]
Jordano, P., and J. A. Godoy. 2000. RAPD variation and population genetic structure in Prunus mahaleb (Rosaceae), an animal–dispersed tree. Molecular Ecol. 9: 1293–1305. [ Links ]
Lacey, E. P., D. A. Roach, D. Herr, S. Kincaid, and R. Perrott. 2003. Multigenerational effects of flowering and fruiting phenology in Plantago lanceolata. Ecology 84:2462–2475. [ Links ]
Oshawa, T., and I. Ide. 2008. Global patterns of genetic variation in plant species along vertical and horizontal gradients on mountains. Global Ecol. Biogeogr. 17:152–163. [ Links ]
Pensiero, J. F. 1999. Las especies sudamericanas del género Setaria (Poaceae, Paniceae). Darwiniana 37(1–2): 37–151. [ Links ]
Pensiero, J. F., H. F. Gutiérrez, y E. Exner. 2005. Sistema de polinización y su efecto sobre la producción y el peso de semillas en nueve especies sudamericanas del género Setaria. Interciencia 30(8):495–500. [ Links ]
Pontes, L. S., P. Carrere, D. Anduezaf, F. Louault, and J. F. Soussana. 2007. Seasonal productivity and nutritive value of temperate grasses found in semi–natural pastures in Europe: responses to cutting frequency and N supply. Grass Forage Sci. 62:485–496. [ Links ]
Quinn, J. A., and J. D. Wetherington. 2002. Genetic variability and phenotypic plasticity in flowering phenology in populations of two grasses. J. Torrey Bot. Soc. 129: 96–106. [ Links ]
Ramakrishnan, A P., S. E. Meyer, J. Waters, M. R. Stevens, C. E.Coleman, and D. J. Fairbanks. 2004. Correlation between molecular markers and Adaptively significant genetic variation in Bromus tectorum (Poaceae), an inbreeding annual grass. Am. J. Bot. 91:797–803. [ Links ]
Slafer, G. A. 2003. Genetic basis of yield as viewed from a crop physiologist's perspective. Ann. Appl. Biol. 142:117–128. [ Links ]
Steel, M. G., P. B. Cavers, and S. M. Lee. 1983. The biology of Canadian weeds. 59. Setaria glauca (L.) Beauv. and 5. verti–cillata (L.) Beauv. Can. J. Plant Sci. 63:711–725. [ Links ]
Stinson, K. A. 2004. Natural selection favors rapid reproductive phenology in Potentilla pukherrima (Rosaceae) at opposite ends of a subalpine snowmelt gradient. Am. J. Bot. 91:531–539. [ Links ]
Vigouroux, Y., Y. Matsuoka, and J. Doebley. 2003. Directiona evolution for microsatellite size in maize. Molecular Biol. Evolution 20:1480–1483. [ Links ]
Wright, J. W., and T. Meagher. 2003. Pollination and seed predation drive flowering phenology in Silene latifolia (Caryophyllaceae). Ecology 84:2062–2073. [ Links ]
Yamagishi, H., T. D. Allison, and M. Ohara. 2005. Effect of snowmelt timing on the genetic structure of an Erythronium grandiflorum population in an alpine environment. Ecol. Res. 20:199–204. [ Links ]

