










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgrociencia
versão On-line ISSN 2521-9766versão impressa ISSN 1405-3195
Agrociencia vol.44 no.5 Texcoco Jul./Ago. 2010
Agua–suelo–clima
Glomalina y secuestro de Carbono en tepetates cultivados
Glomalin and Carbon sequestration in cultivated tepetates
Aurelio Báez–Pérez1,3, M. C. Ángeles González–Chávez1, Jorge D. Etchevers–Barra1*, Christian Prat2, Claudia Hidalgo–Moreno1
1 Edafología. Campus Montecillo, Colegio de Postgraduados. 56230, Montecillo. Estado de México. (etchev@colpos.mx).
2 IRD IRD–LTHE, BP 53,38 041 Grenoble, Cedex 9. Francia.
3 Dirección actual: INIFAP, Campo Experimental Bajío. Carretera Celaya–San Miguel Allende km 7.5. C.P. 381100, Celaya, Guanajuato.
Recibido: Octubre, 2009.
Aprobado: Junio, 2010.
RESUMEN
Los tepetates son tobas volcánicas endurecidas que afloran en la superficie debido a procesos erosivos; algunos de éstos pueden habilitarse para la agricultura y con ello tienen potencial para secuestrar carbono (C). Este elemento se acumula en los frágiles agregados que se forman en su estructura física después de cultivarse, pero se desconoce el mecanismo por el cual es secuestrado en la fracción mineral en estos sustratos volcánicos. Se presume que la actividad biológica contribuye la estabilización del C orgánico (COS), específicamente la de los hongos micorrízico arbusculares (HMA). Se ha postulado que la glomalina (una glicoproteína producida por los HMA) influye en ambos procesos. Para contribuir a la comprensión de este fenómeno, se realizó el presente estudio con los siguientes objetivos: 1) evaluar la acumulación del COS, la glomalina y el carbono de la glomalina (CG) en tepetates cultivados; 2) medir el aporte de C que hace la glomalina al COS; 3) evaluar la tendencia de acumulación del CG con los años de cultivo. Con base en entrevistas a productores de la cuenca del río Texcoco, se seleccionaron 83 parcelas con 10 a 20 años de cultivo, y se definieron 10 clases de manejo agronómico (I al X). Como referencia se usaron cuatro suelos Faeozem de la región agrupados en dos clases de manejo. Otras 21 parcelas de tepetate tenían de 4 a más de 100 años de cultivo. Se recolectaron muestras de tepetate (0–20 cm) y se determinó el COS, la glomalina y el CG. La relación entre el COS y las últimas variables fue estrecha (R>0.91). El manejo agronómico influyó significativamente (p<0.05) en la acumulación del COS, la glomalina y el CG. En función de la máxima concentración de COS observada, se calculó que los tepetates habilitados para la producción agrícola después de dos décadas de cultivo, y un manejo agronómico con constante incorporación de residuos orgánicos, tienen capacidad para almacenar aproximadamente 90 t ha–1 de C en los primeros 20 cm de profundidad, con una aportación de 15 % de C estabilizado, proveniente de la glomalina (p<0.05). La acumulación del CG en función del tiempo siguió una tendencia logarítmica.
Palabras clave: captura de carbono, hongos micorrízicos arbusculares, manejo agronómico, tobas volcánicas.
ABSTRACT
The tepetates are hardened volcanic tuffs that outcrop on the surface due to erosive processes; some of them can be ameliorated for agriculture and thus sequester carbon (C). This element accumulates in fragile aggregates that are formed as part of their developing physical structure after being cultivated, but the mechanism by which it is sequestered in the mineral fraction in these volcanic substrates is unknown. It is assumed that the biological activity contributes to the stabilization of the organic C (COS), specifically that of the arbuscular mycorrhizal fungi (AMF). It has been postulated that glomalin (a glycoprotein produced by the AMF) influences both processes. To contribute to the comprehension of this phenomenon, the present study was made with the following objectives: 1) to evaluate the accumulation of COS, glomalin and glomalin carbon (GC) in cultivated tepetates; 2) to measure the contribution of C made by the glomalin to the COS; and 3) to evaluate the tendency of accumulation of GC with years of cultivation. Based on interviews with producers of the Texcoco River basin, 83 plots were selected with 10 to 20 years of cultivation, and 10 classes of agronomic management were defined (I to X). As reference four Phaesozem soils of the region were used, grouped in two classes of management. Another 21 plots of tepetate had from 4 to over 100 years of cultivation. Tepetate samples were collected (0–20 cm) and COS, glomalin and GC were determined. The relationship between COS and the latter variables was close (R>0.91). The agronomic management had significant influence (p<0.05) on the accumulation of COS, glomalin and GC. As function of the maximum concentration of COS observed, it was calculated that the tepetates ameliorated for agricultural production after two decades of cultivation, with an agronomic management with constant incorporation of organic residues, have capacity for storing approximately 90 t ha–1 of C in the first 20 cm of depth, with a contribution of 15 % of stabilized C from glomalin (p<0.05). The accumulation of GC as a function of time followed a logarithmic tendency.
Key words: carbon sequestration, arbuscular micorrhyzic fungi, agronomic management, volcanic tuffs.
INTRODUCCIÓN
Los tepetates son capas de materiales de origen volcánico (tobas) endurecidas por procesos geológicos y pedológicos, muchas veces cubiertas por un suelo superficial (Etchevers et al., 2006). En el Eje Neovolcánico Transmexicano cubren poco más de 37 000 km2 de superficie (Zebrowski, 1992). Procesos erosivos naturales e inducidos por el hombre han provocado el afloramiento de las capas subyacentes de tepetates en extensas áreas de esa región. Se reconocen dos tipos de tepetate de acuerdo con su grado de dureza: fragipán y duripán (Hidalgo et al., 1997). El primero es duro cuando seco pero se reblandece cuando húmedo, y el segundo es duro en estado seco y húmedo. El tepetate tipo fragipán se puede habilitar para la producción agrícola o forestal mediante prácticas de roturación mecánica y un manejo agronómico adecuado, que incluye elevar su contenido de materia orgánica, lo cual favorece su agregación y fertilidad (Báez et al., 2007a; Navarro et al., 2003). La materia orgánica además induce la actividad biológica en el sustrato roturado, y mejora sus propiedades físicas como su capacidad de infiltración y de retención de agua (Oades, 1988).
De especial interés para la agregación son los hongos micorrízico arbusculares (HMA) que contribuyen a la formación y estabilidad de agregados del suelo. El micelio de dichos organismos y las sustancias que secretan aglutinan las partículas elementales del suelo (González–Chávez et al., 2004; Wright y Upadhyaya, 1996). Asociada con la actividad de las hifas de los HMA se ha identificado una glicoproteína llamada glomalina, un compuesto orgánico específico de estos hongos (Wright y Upadhyaya, 1998). La glomalina almacena C en subunidades de proteína y carbohidratos, que contiene 30 a 40 % de C orgánico, y funciona como cementante, aglutinando fuertemente las partículas de la fracción mineral del suelo y proporcionando estabilidad a los agregados (Franzluebbers et al., 2000; Weller, 2002). La glomalina es aparentemente recalcitrante pudiendo durar en los suelos de 7 a 42 años según las condiciones ambientales, el ecosistema y el manejo agrícola de los suelos (Wright y Upadhyaya, 1998; Rillig et al. 2001; Weller, 2002), es insoluble en agua y en soluciones salinas comúnmente usadas en extracciones de compuestos del suelo (Wright y Jawson, 2001). Según Rillig et al. (2003) la glomalina puede usarse como un indicador de los efectos del cambio de uso de suelo. Sin embargo, la proliferación de HMA, y por tanto la producción de glomalina, es limitada en suelos con alta disponibilidad de P y por algunas prácticas comunes en la agricultura como: fumigación, fertilización y uso de plaguicidas. Estos factores afectan negativamente el establecimiento y funcionalidad de los hongos micorrízico arbusculares (Ryan y Graham, 2002). Lo anterior presupone que la producción de glomalina sería limitada en esas condiciones.
Debido a que los tepetates prácticamente no contienen C orgánico (COS), N y P en su estado natural, cuando se acondicionan para producción agrícola o forestal, es posible estudiar la dinámica de acumulación de estos elementos. El COS fue evaluado por Báez et al. (2002), quienes concluyen que su tasa de acumulación depende del manejo agronómico y del tiempo de cultivo; sin embargo, su acumulación no es lineal, sino que sigue una tendencia logarítmica. Es decir, la tasa de acumulación del COS decrece después de una rápida acumulación de este elemento durante los primeros años de cultivo, alcanzando un equilibrio entre la acumulación y las pérdidas. Estas últimas son ocasionadas principalmente por efecto de la erosión hídrica y emisiones de CO2, resultado de la mineralización de la materia orgánica por la oxidación química y la actividad biológica (Báez et al., 2009). En los tepetates el COS no se ha relacionado con la actividad microbiana, en especial con la de los HMA, y se desconoce cuál es la proporción de C recalcitrante que aporta la glomalina a éstos, cuando son habilitados para la agricultura. Los objetivos del presente estudio fueron: 1) evaluar la acumulación de glomalina y del C contenido en ella, y analizar la relación que guarda con el COS en tepetates cultivados con diferente manejo agronómico; 2) medir el aporte de C de la glomalina al COS en estos materiales volcánicos; y 3) evaluar la tendencia de acumulación del CG con los años de cultivo.
MATERIALES Y MÉTODOS
El estudio se realizó con tepetates recolectados de la cuenca del río Texcoco (Figura 1), Estado de México (98° 45' y 98° 50' O y 19° 27' y 19° 32' N), entre 2300 y 2900 m de altitud, un régimen de humedad ústico a údico, con precipitación de 600 a 900 mm año–1. Estos materiales volcánicos se ubican en el piedemonte de la Sierra Nevada entre 2400 y 2800 m de altitud, en clima templado subhúmedo. Los tepetates se caracterizan por presentar una matriz mineral masiva, poco fragmentada y con abundante presencia de vidrio volcánico y materiales amorfos (Quantin et al., 1993).
Las muestras de material usadas para este experimento fueron las mismas recolectadas para un estudio preliminar sobre cambios en los aspectos físicos de los tepetates, provocados por el tiempo transcurrido desde su habilitación para la agricultura (Báez et al., 2002, 2007b). Entre ellas se encuentran también muestras recolectadas en camas de invernadero construidas con tepetate. De esos estudios proviene una base de datos que contiene la siguiente información acerca de los tepetates habilitados para la agricultura: año en que se roturaron las parcelas, especies cultivadas, manejo de residuos de cosecha, adición de fertilizantes químicos y orgánicos y uso de agua de riego. Las parcelas muestran una amplia variación en cuanto al tiempo transcurrido desde su habilitación y al manejo agronómico; con base en este último se había agrupado en 10 clases (Cuadro 1) correspondientes a los tepetates cultivados (83 parcelas). Como referencias con lo que acontece con un suelo verdadero se incluyeron cuatro suelos (Faeozem) de la misma región, los cuales por su manejo agronómico con diferente intensidad se agruparon en dos clases: SFI y SFII (Cuadro 1). Estos suelos se encuentran en la vecindad de las parcelas de tepetate muestreadas y sirvieron como contraste con los tepetates habilitados. Otras 21 parcelas de tepetate tenían de 4 a más de 100 años de cultivo y se usaron para calcular la tendencia de acumulación del CG en función del tiempo de cultivo.
El pH de los tepetates usados en el presente estudio varió de ligeramente ácido a alcalino (6.8 a 8.1) y la clase textural se ubicó en el intervalo de franca a arcillosa. Esta heterogeneidad es propia de las capas de toba volcánica. La clasificación del tipo de tepetate t3 propuesta por Quantin et al. (1993) consideró criterios geológicos, pedológicos, estratigráficos y de mineralogía. Sin embargo, existen otros factores que influyen en la heterogeneidad de estos materiales volcánicos relacionados con la forma de depósito de las cenizas volcánicas y el grado de alteración de los minerales primarios y secundarios.
Se recolectaron muestras compuestas (por 22 submuestras) de tepetate cultivado de 0 a 20 cm de profundidad; fueron secadas al aire a temperatura ambiente, se molieron y tamizaron (2 mm). Para medir el COS y el C inorgánico, las muestras se molieron con un mortero de ágata y se tamizaron por malla 100 (0.14 mm). Para determinar el C se usó un analizador automático de C total (Shimadzu, TOC–5050). Para la extracción de glomalina se usó citrato de sodio 50 mM a pH8 y autoclave por 1 h (1.3 kg cm–2) según los procedimientos de extracción, precipitación y diálisis propuestos por Wright y Jawson (2001). El CG se midió en los extractos dializados de glomalina en el analizador TOC–5050. El diseño experimental fue completamente al azar con un número desigual (3 a 15) de repeticiones por clase de manejo (Cuadro 1). La comparación de medias se efectuó con varianzas ponderadas por tamaño de muestra, procedimiento GLM y opción REGWQ (SAS, 1998). También se calculó el error estándar.
RESULTADOS Y DISCUSIÓN
COS y glomalina en tepetates cultivados
La menor cantidad de glomalina y COS se observó en la clase de manejo I; tepetate recién roturado, donde se midieron sólo trazas de ambas sustancias (0.14 mg g–1 y 0.11 %; Figura 2). Estos valores se consideraron como el punto de referencia inicial de los tepetates en su condición natural. La concentración de glomalina en esta clase de manejo fue muy baja comparada con las concentraciones típicas observadas en otros tipos de suelo (2–15 mg g–1), y más cercanos al intervalo de valores reportados (0.00311 mg g–1) para suelos áridos, que en ocasiones tienen escasa vegetación (Bird et al., 2002; Treseder y Turner, 2007). Según Bird et al. (2002) y Rillig et al. (2003), los suelos cubiertos con especies arbóreas o pastos presentan mayor concentración de glomalina que los suelos libres de vegetación, lo cual se explica por la presencia permanente de plantas que propician abastecimiento permanente de fotosintatos para alimentar los HMA estrechamente relacionados con producción de glomalina (González–Chávez et al., 2004). Sin embargo, en la presente investigación la presencia de plantas en el tepetate no presentó este mismo efecto. La concentración de COS y glomalina en las parcelas correspondientes a las clases de manejo agronómico II, III y IV, con escaso ingreso anual de residuos orgánicos, fue cuatro veces mayor (0.5 a 0.6 % de COS y 0.3 a 0.4 mg g–1 de glomalina; p>0.05) a lo observado en el tepetate de la clase I (recién roturado y sin cultivar). En estas últimas clases de manejo agronómico, los residuos de cosecha se usan principalmente para alimentar al ganado y la incorporación de abono orgánico al suelo es prácticamente nula. Por tanto, el aumento en el COS, la proliferación de HMA y, en consecuencia, la producción de glomalina es en parte limitada por la ausencia de materia orgánica. Esto se corrobora con los niveles significativamente mayores de COS, pero no de glomalina en la clase V, en la cual se aportan abonos orgánicos en mayor proporción (> 3 t ha–1 año–1) que en las clases anteriores (< 1 t ha–1 año–1).

En la clase de manejo agronómico VI, con leguminosas, la concentración promedio de glomalina fue 0.52 mg g–1, significativamente mayor (p>0.05) a la observada en la clase I. La incorporación de leguminosas, ya sea en rotación o en asociación con cereales, y la adición de estiércol, favorecen el rendimiento de trigo, cebada y maíz (Báez et al., 2007a; Navarro et al., 2003). El cultivo de leguminosas tiene un efecto biológico relevante en el suelo por su capacidad de asociación en doble simbiosis con bacterias fijadoras de N y HMA (Álvarez–Solís et al., 2000). Lo anterior significa una mayor disponibilidad regular de N y P para las plantas. La actividad de los HMA favorece además la producción de glomalina (González–Chávez et al., 2004).
La clase de manejo VII, con cultivos perennes de temporal sin labranza, mostró una concentración promedio de COS y glomalina semejante (p>0.05) a lo reportado en las clases V y VI, donde se adicionaron regularmente abonos orgánicos. Los cultivos establecidos, principalmente el agapando (Agapanthus africanus L.), favorecen un aporte regular de residuos de orgánicos al sistema y proporcionan una cobertura vegetal permanente en el suelo. Esto favorece la conservación de la humedad, la actividad biológica y debería incrementar la proliferación de HMA y la producción de glomalina, como lo reportan Borie et al. (2006) y West et al. (2005). Sin embargo, en el presente trabajo, no fueron significativamente superiores al observado en la clase I.
Las clases de manejo VIII y IX mostraron un aumento de glomalina mayor de 30 % (p<0.05) con respecto a la clase VII. Fue notable que los cultivos perennes con labranza cero tuvieron mayor capacidad para acumular COS y producir glomalina, comparado con los sistemas agrícolas con labranza convencional (barbecho, rastreo, surcado y aporque). La labranza convencional permite la oxidación de C y su pérdida como CO2. Con labranza cero la materia orgánica que queda en la superficie del suelo reduce la evaporación, la temperatura y el intercambio de gases que intervienen en la oxidación biológica de la misma (Reicosky y Lindstrom, 1993). En estas condiciones los HMA, relacionados con la glomalina, también son más abundantes con labranza cero, porque no hay ruptura de hifas y éstas contribuyen mayormente a la estabilidad de los agregados del suelo (Rillig et al., 2002; Franzluebbers et al., 2000) que participan en la protección física del C lábil del suelo y con efecto en el secuestro de carbono (Six et al. 2006).
La concentración de COS y glomalina fue 1.5 % y 1.2 mg g–1 en la clase VIII, la cual correspondió a tepetates usados como camas de invernadero. Se observó un aumento en la cantidad de glomalina (p<0.05) de más del doble, respecto a los casos anteriores. En la clase X, que también correspondió a camas de invernadero pero con adición de composta de madera, las concentraciones de COS y glomalina fueron más del doble respecto a lo observado en la clase VIII. Lo anterior representó el máximo nivel de COS (3.9 %) y glomalina (2 mg g–1) encontrado en los tepetates cultivados. Al respecto, West et al. (2005) mencionan que las enmiendas orgánicas en el suelo mejoran la actividad micelial de los HMA y la producción de glomalina, porque promueven la retención de agua en el suelo.
Al comparar las concentraciones de COS y de glomalina de la clase X con los suelos Faeozem sometidos a un manejo agronómico convencional (SFI) y usados como referencia, ya que los tepetates estrictamente no son suelos, se evidenció que los tepetates tienen potencial para acumular COS. Los resultados observados en este estudio muestran que a diferencia de las prácticas usadas en la agricultura convencional (monocultivos y labranza), las prácticas agronómicas conservacionistas (labranza cero, rotación de cultivos y uso de leguminosas) promueven una mayor producción de glomalina, lo cual coincide con lo reportado por Purin et al. (2006), Roldán et al. (2006), la cual se derivaría de la actividad fúngica en el suelo (Oehl et al. 2003; Kurle y Pfleger, 1994). Como resultado de ello se favorece la acumulación de materia orgánica del suelo (Six et al., 2006).
La concentración de COS y glomalina del suelo de referencia (SFII), con cultivos perennes, sin labranza y con riego, fue semejante a lo observado en la clase X (camas de invernadero). Lo anterior destaca el potencial que tienen los tepetates para la acumulación de COS, que según el caso más alto, fue cerca de 4 % de COS y 2.2 mg g–1 de glomalina. Lo anterior se logró con un manejo tendiente a conservar la materia orgánica (Báez et al., 2002). Resulta relevante mencionar que los diferentes cultivos en el SFII corresponden a especies vegetales micorrízicas, las cuales son hospederas naturales de hongos micorrízico arbusculares, mismos que no fueron medidos en este experimento.
El COS se relacionó estrechamente con la glomalina y con el CG (R=0.91 y R=0.94). Rillig (2004) reporta resultados similares y sugiere la factibilidad de utilizar esta glicoproteína como un indicador del cambio de uso del suelo. West et al. (2005) también reportan una alta correlación entre COS y glomalina, y sugieren que el mayor almacén de C puede ser dependiente de los hongos micorrizíco arbuculares.
Aporte de C de la glomalina al COS en los tepetates cultivados
En el presente trabajo, el C contenido en la glomalina (CG) representó una fracción mínima del COS (Figura 3), que estuvo en un intervalo de trazas en las clases de manejo I, II, III, IV, V y XI, hasta aproximadamente 15 % del COS en la clase de manejo X. Este porcentaje es similar al suelo de referencia II, donde se observó la mayor concentración de este elemento. Según Weller (2002), la glomalina puede aportar hasta 27 % del COS al suelo. En el presente estudio, el aporte de glomalina al COS fue menor debido a que el tepetate no es un suelo sino un sustrato habilitado artificialmente para la producción agrícola y que carece de manera natural principalmente de COS, N y P (Etchevers et al., 1992), elementos esenciales para la actividad biológica. Por tanto los tepetates en su estado natural contienen escasas poblaciones de microorganismos (Álvarez–Solís et al., 2000). Además su estructura física carece de agregados. La evolución de sus características físico–químicas y la formación de agregados depende del manejo agronómico y el tiempo de cultivo (Báez et al., 2007b). La humedad es otro factor que parece limitar la producción de glomalina en estos sistemas.
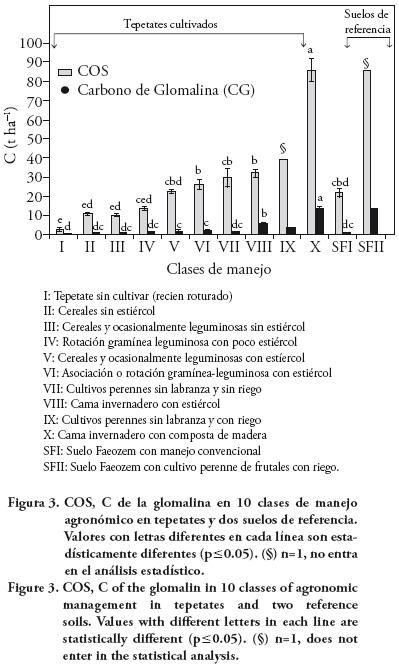
El potencial que tienen los tepetates para la acumulación de COS, según el caso más alto, fue aproximadamente 90 t ha–1 con una aportación promedio de 13.5 t ha–1 de C asociado a la glomalina, equivalente a 15 % del COS. Sin embargo, con concentraciones menores de COS, el aporte del C estabilizado proveniente de la actividad de los HMA es menor de 10 %.
Acumulación del carbono de la glomalina (CG) y años de cultivo
La tasa de acumulación del CG en función de los años de cultivo en estos tepetates se muestra en la Figura 4, donde se presentan dos modalidades de manejo agronómico contrastantes: uno, en monocultivo de cereales con escaso ingreso de residuos orgánicos, y otro, en rotación de cultivos (gramínea–leguminosa) con constante incorporación de residuos orgánicos. Es evidente que en ambos casos se observó una tendencia de acumulación logarítmica, y el comportamiento fue semejante a la tasa de acumulación del COS descrita por Báez et al. (2002) para los tepetates cultivados. Es decir, después de una rápida acumulación de este elemento en los primeros años de cultivo, ésta decrece, alcanzando un equilibrio entre la acumulación y las pérdidas de carbono. Las primeras son función del manejo agronómico y del ingreso de residuos orgánicos al agrosistema, y las segundas, de la erosión hídrica y la mineralización química–biológica. Estas últimas corresponden principalmente a las emisiones de CO2 (Báez et al., 2009).
El CG, fracción de C estabilizado representó en estas clases de manejo aproximadamente una décima parte del COS total durante los años de cultivo (Figura 4). El equilibrio alcanzado muestra que se requeriría mayores ingresos de C y menores pérdidas para aumentar los niveles de COS y CG. Esto explica en parte porque la acumulación de C en los tepetates es limitada aún después de décadas de cultivo. Para un mejoramiento sustancial de la estructura física de los tepetates, que está relacionada con la estabilidad de agregados, se requiere también un aumento significativo de C estabilizado.
CONCLUSIONES
La concentración de glomalina varió de trazas a 2 mg g–1 en los tepetates cultivados y de 0.2 a 2.2 mg g–1 en los suelos Faeozem, usados como referencia. Hubo una estrecha relación (R=0.91) entre la glomalina y CG con el COS. El potencial de acumulación de carbono estabilizado (CG) proveniente de la actividad de los HMA en los tepetates cultivados, fue semejante al acumulado en los suelos agrícolas de la misma región, con alrededor de 13.5 t ha–1. De acuerdo con el máximo valor de COS determinado en los tepetates, éstos pueden almacenar más de 90 t ha–1, en los primeros 20 cm de profundidad, con una aportación de 15 % del C por parte de la glomalina. La acumulación del CG durante los años de cultivo siguió una tendencia logarítmica, semejante a la observada en la acumulación del carbono orgánico.
AGRADECIMIENTOS
Al IRD (ex–ORSTOM), a INSTRUCT, al CONACYT y la Unión Europea (REVOLSO Proyecto ICA4–CT–2001–0052) por el financiamiento otorgado para iniciar esta investigación.
LITERATURA CITADA
Álvarez–Solís, D., R. Ferrera–Cerrato, y J. D. Etchevers. 2000. Actividad microbiana en tepetates con incorporación de residuos orgánicos. Agrociencia 34:523–532. [ Links ]
Báez, A., J. D. Etchevers, C. Hidalgo, C. Prat, V. Ordaz, y R. Núñez. 2002. C orgánico en tepetates cultivados de México. Agrociencia 36:643–653. [ Links ]
Báez, A., J. D. Etchevers, C. Prat, A. Márquez, y E. Ascencio–Zapata. 2007a. Manejo agronómico de los suelos endurecidos (tepetates) del eje neovolcánico de México. In: Gallardo–Lancho, J. (ed). La Captura de Carbono en Ecosistemas Terrestres Iberoamericanos. Sociedad Iberoamericana de Física y Química Ambiental. Salamanca, España, pp: 69–84. [ Links ]
Báez, A., J. D. Etchevers, C. Prat, and C. Hidalgo. 2007b. Formation of aggregates and carbon sequestration in ameliorated tepetates in the Río Texcoco basin, Mexico. Rev. Mex. Ciencias Geol. 24: 487–497. [ Links ]
Báez, A., J.D. Etchevers, M. Haulon, G. Werner, G. Flores, y C. Hidalgo. 2009. Pérdida de carbono por erosión hídrica y emisiones de CO2 en tepetates habilitados para la agricultura. In: Gallardo, J.F., J. Campo y M.E. Conti (eds). Emisiones de Gases con Efecto Invernadero en Ecosistemas Iberoamericanos. Sociedad Iberoamericana de Física y Química Ambiental. Salamanca, España. pp: 25–48. [ Links ]
Bird, S.B., J.E. Herrick, M.M. Wander, and S.F. Wright. 2002. Spatial heterogeneity of aggregate stability and soil carbon in semi–arid rangeland. Environ. Poll. 116: 445–455. [ Links ]
Borie, F., R. Rubio, J. L. Rouanet, A. Morales, G. Borie, and C. Rojas. 2006. Effects of tillage systems on soil characteristics, glomalin and mycorrhizal propagules in a Chilean Ultisol. Soil Tillage Res. 88: 253–261. [ Links ]
Etchevers, J. D., R. M. López, C. Zebrowski, y D. Peña. 1992. Características químicas del tepetate de referencia de los estados de México y Tlaxcala, México. Terra 10 (Número especial): 171–177. [ Links ]
Etchevers, J. D., C. Hidalgo, C. Prat, and P. Quantin. 2006. Tepetates of Mexico. Encyclopedia of Soil Science. Marcel Dekker, New York, USA. pp: 1745–1748. [ Links ]
Franzluebbers, A. J., S. F. Wright, and J. A. Stuedemann. 2000. Soil aggregation and glomalin under pastures in the Southern Piedmont USA. Soil Sci. Soc. Am. J. 64:1018–1026. [ Links ]
González–Chávez, M. C. A., M. C. Gutiérrez, y S. Wright. 2004. Hongos micorrízico arbusculares en la agregación del suelo y su estabilidad. Terra Latin. 22: 507–214. [ Links ]
Hidalgo M., C., P. Quantin, y F. Elsass. 1997. Caracterización mineralógica de los tepetates tipo fragipán del Valle de México. In: Zebrowski, C., P. Quantin, y G. Trujillo (eds). III Simposio Internacional: Suelos Volcánicos y Endurecidos. ORSTOM. Quito, Ecuador. pp: 62–72. [ Links ]
Kurle, J. E., and F. L. Pfleger. 1994. The effects of cultural practices and pesticides on VAM fungi. In: Pfleger F. L., and R. G. Linderman (eds). Mycorrhizae and Plant Health. Minnesota APS Press. pp: 101–132. [ Links ]
Navarro, H., A. Pérez, y D. Flores. 2003. Productividad de los tepetates con sistemas rotacionales. Terra Latin. 22:71–79. [ Links ]
Oades, J. M. 1988. The retention of organic matter in soils. Biogeochemistry 5:35–70. [ Links ]
Oehl, F., E. Sieverding, K. Ineichen, P. Mäder, T. Boller, and A. Wiemken, A. 2003. Impact of land use intensity on the species diversity of arbuscular mycorrhizal fungi in agroecosys–tems of Central Europe. Appl. Environ. Microbiol. 69: 2816–2824. [ Links ]
Purin, S., O. F. Klauberg, and S. L. Stürmer. 2006. Mycorrhizae activity and diversity in conventional and organic apple orchards from Brazil. Soil Biol. Biochem. 38:1831–1839. [ Links ]
Quantin, P., H. Arias, J. D. Etchevers, R. Ferrera–Cerrato, K. Olechko, H. Navarro, G. Werner, y C. Zebrowski. 1993. Tepetates de México: caracterización y habilitación para la agricultura: Terra Latin. 11:33–34. [ Links ]
Reicosky, D., and M. Lindstrom. 1993. Effect of fall tillage on short term carbon dioxide flux from soil. Agron. J. 85:1237–1243. [ Links ]
Rillig, M. C., S. F Wright, K. A. Nichols, W. F. Schmidt, and M. S. Torn. 2001. Large contribution of arbuscular mycorrhizal fungi to soil carbon pools in tropical forest soils. Plant Soil 233: 167–177. [ Links ]
Rillig, M. C., S. F. Wright, and V. T. Eviner. 2002. The role of arbuscular mycorrhizal fungi and glomalin in soil aggregation: comparing effects of five plant species. Plant Soil 238:325–333. [ Links ]
Rillig, M. C., P. W. Ramsey, S. Morris, and E. A. Paul. 2003. Glomalin, an arbuscular–mycorrhizal fungal soil protein, responds to land–use change. Plant Soil 253:293–299. [ Links ]
Rillig, M. C. 2004. Arbuscular mycorrhizae and terrestrial ecosystem processes. Ecol. Lett. 7:740–754. [ Links ]
Roldán, A., J.R. Salinas–García, M. M. Alguacil, and F. Caravaca. 2006. Soil sustainability indicators following conservation tillage practices under subtropical maize and bean crops. Soil Tillage Res. 93:273–782. [ Links ]
Ryan, M. G., and J. H. Graham. 2002. Is there a role for arbuscular mycorrhizal fungi in production agriculture? Plant Soil 244:263–271. [ Links ]
SAS Institute. 1998. SAS/SAT User's Guide. Version 6.09. SAS Inst., Cary. NC. pp: 100–120. [ Links ]
Six, J., S. D. Frey, R. K. Thiet, and K. M. Batten. 2006. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 70:555–569. [ Links ]
Treseder, K. K., and K. M. Turner. 2007. Glomalin in ecosystems. Soil Sci. Soc. Am. J. 71:1257–1266. [ Links ]
Weller, K. 2002. Glomalin: hiding place for a third the world's stored soil carbon. Agric. Res. Magazine 50: 4–7. [ Links ]
West, S. B., T. C. Caesar–TonThat, S. F. Wright, and J. D. Williams. 2005. Organic matter addition, N, and residue burning effects on infiltration, biological, and physical properties of an intensively tilled siltloam soil. Soil Tillage Res. 84:154–167. [ Links ]
Wright, S. F., and A. Upadhyaya. 1996. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Sci. 198:97–107. [ Links ]
Wright, S., and A. Upadhyaya. 1998. A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular micorrhizal fungi. Plant and Soil 198:97–107. [ Links ]
Wright, S. F., and L. Jawson. 2001. A pressure cooker method to extract glomalin from soils. Soil Sci. Soc. Am. J. 65:1734–1735. [ Links ]
Zebrowski, C. 1992. Los suelos volcánicos endurecidos en América Latina. Terra 10 (Número especial): 15–23. [ Links ]

