










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.43 no.7 Texcoco oct./nov. 2009
Protección vegetal
Etiología e histopatología de la muerte descendente de árboles de mamey (Pouteria sapota (Jacq.) H. E. Moore y Stearn) en el estado de Guerrero, México
Etiology and histopathology of dieback disease on mamey trees (Pouteria sapota (Jacq.) H. E. Moore and Stearn) in Guerrero, Mexico
Alfonso Vásquez–López1,* , José A. Mora–Aguilera2, Elizabeth Cárdenas–Soriano2, Daniel Téliz–Ortiz2
1 Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Unidad Oaxaca, Instituto Politécnico Nacional. Calle Hornos 1003, 71230, Santa Cruz Xoxocotlán, Oaxaca. *Autor responsable: (bremia43@gmail.com).
2 Fitosanidad–Fitopatología, Campus Montecillo, Colegio de Postgraduados. Carretera México–Texcoco km. 36.5, 56230. Montecillo, Estado de México.
Recibido: Junio, 2008.
Aprobado: Mayo, 2009.
RESUMEN
En Alpoyeca, Guerrero, México, la muerte descendente de los árboles de mamey (Pouteria sapota) es una enfermedad de etiología desconocida que reduce la productividad y longevidad de los huertos comerciales. En este estudio se determinó la etiología de la enfermedad y los daños anatómicos que causa en ramas. De ramas sintomáticas se aisló consistentemente a Lasiodiplodia sp. y su patogenicidad se verificó en ramas de mamey con epidermis y peridermis (con y sin heridas). Después de 30 d el hongo produjo lesiones necróticas en ramas con peridermis inoculadas por heridas y en 24 meses indujo muerte descendente de brotes vegetativos. Las ramas testigo se mantuvieron asintomáticas. Los daños anatómicos en ramas sintomáticas fueron ruptura de peridermis; colapso y nectosis de células corticales, floema, laticíferos, fibras externas del floema y cambium vascular. En floema se formaron cristales y precipitados granulosos; los radios y el paténquima axial se lignificaron y se formaron esclereidas. El xilema y la médula no presentaron daños. Con base en sus características morfológicas y moleculares el hongo que indujo muerte descendente de árboles de mamey en Alpoyeca, Guerrero fue Lasiodiplodia theobromae. La secuencia genética del patógeno se depositó en el banco de genes (NCBI) con número de acceso EU564805.
Palabras clave: Lasiodiplodia theobromae, Histopatología, muerte descendente, zapote–mamey.
ABSTRACT
In Alpoyeca, Guerrero, México, dieback on mamey trees (Pouteria sapota) is a disease of unknown etiology that reduces productivity and longevity of commercial orchards. In this study the etiology of the disease and anatomic damage it causes to branches were determined. From symptomatic branches, Lasiodiplodia sp. was consistently isolated, and its pathogenicity was verified on mamey branches with epidermis and peridermis (with and without lesions). After 30 d, the fungus produced necrotic lesions on branches with peridermis inoculated through wounds, and in 24 months it induced dieback of vegetative shoots. The control branches remained asymptomatic. The anatomical damage on symptomatic branches included peridermis rupture, collapse and necrosis of cortical, phloem, and laticifetous cells, and external phloem fibers and vasculat cambium. In the phloem, crystals and granular precipitates formed; the radial and the axial parenchyma lignified, and sclereids formed. The xylem and medulla were not damaged. Based on its motrphological and molecular characteristics, the fungus that induced dieback on mamey trees in Alpoyeca, Guerrero, was Lasiodiplodia theobromae. The genetic sequence of the pathogen was deposited in the gene bank (NCBI) with access number EU564805.
Key words: Lasiodiplodia theobromae, Histopathology, dieback, mamey sapote.
INTRODUCCIÓN
El mamey (Pouteria sapota (Jacq.) H. E. Moore y Stearn) es un cultivo primario que soporta aproximadamente 95 % de las familias en los municipios de Huamuxtitlán y Alpoyeca, Guerrero, México generando ingresos importantes de noviembre a marzo de cada año, debido a la precocidad de la producción respecto a otras zonas productoras del país. Sin embargo, los problemas fitosanitarios reducen la productividad y longevidad de los huertos comerciales. Con el fin de diseñar un programa de manejo integrado del cultivo se estudiaron las enfermedades relevantes de este frutal. En Alpoyeca, Guerrero, se encontró una enfermedad de etiología desconocida que induce muerte descendente de ramas estructurales y reproductivas en árboles mayores de 10 años de edad. En el mundo la enfermedad no se había estudiado con precisión. El síndrome de muerte descendente, rajadura de corteza y cáncer de tallo en árboles de zapote (P sapota) en Guatemala se asoció a Botryosphaeria sp. e Hypoxylon sp. (Álvarez, 1997) y el tizón y muerte de ramas jóvenes se asoció a Botryosphaeria theobromae Pat. y a Cephaleurus virescens Kunze (Patel y Chauhan, 1993; Balerdi et al, 1996). En este trabajo se postula que Lasiodiplodia sp. induce muerte celular en el tejido infectado y causa síntomas de muerte descendente, por lo que se estudió la etiología y daños anatómicos que induce la muerte descendente en árboles de mamey en Alpoyeca, Guerrero, México.
MATERIALES Y MÉTODOS
Etiología
Sitio de estudio y aislamiento de hongos
En el municipio de Alpoyeca 17° 40' N y 98° 31' O, 960 m de altitud (INEGI, 2006) se recolectaron 25 ramas reproductivas (2 cm diámetro) con síntomas de muerte descendente de árboles de mamey de 14 años de edad, en mayo de 2004. De cada rama se cortaron cinco secciones de tejido interno de 0.5 cm3 que se desinfestaron con hipoclorito de sodio a 1.0 % por 3 min, se lavaron con agua destilada estéril, se secaron y sembraron en medio de cultivo papa–dextrosa–agar (PDA) (200 g papa + 12 g dextrosa + 18 g agar L–1 agua destilada). Las cajas se incubaron a 25 °C con luz blanca por 3 d. De los aislamientos obtenidos se realizaron cultivos monospóricos en agua agar (18 g agar en 1.0 L–1 agua destilada) y aumentaron en PDA.
Pruebas de patogenicidad
Fase de laboratorio. Esta fase experimental fue para seleccionar un aislamiento de Lasiodiplodia y verificar su patogenicidad en árboles de mamey. Se evaluó la severidad del daño de 16 aislamientos de Lasiodiplodia en frutos de mamey en madurez fisiológica. Estos aislamientos se obtuvieron en 58 % de las 25 ramas muestreadas. Se inoculó un fruto por aislamiento y hubo cuatro testigos. Se desinfestaron 20 frutos con hipoclorito de sodio a 1.5% por 5 min y se lavaron con agua destilada estéril. En cada fruto se sobrepusieron discos miceliales (0.5 cm diámetro) de 20 d de edad en dos sitios con y sin herida en las secciones apical, central y basal de cada fruto. Las heridas, hechas con un bisturí estéril, fueron cortes (1.0 cm longitud) que atravesaron el epicarpio del fruto. En frutos testigo se colocaron discos de PDA sin inoculo. Los frutos se incubaron en cámara húmeda a 22±2 °C por 12 d. Se evaluó la severidad de daños mediante la dimensión (largo, ancho y profundidad) y volumen del área necrótica de las pudriciones fungosas en la pulpa del fruto y se seleccionó el aislamiento más agresivo de Lasiodiplodia sp., para verificar su patogenicidad en ramas anuales.
Patogenicidad en campo. La patogenicidad del aislamiento de Lasiodiplodia seleccionado en laboratorio se verificó en árboles de mamey de 14 años de edad en 2005. Se probaron cuatro tratamientos de inoculación: ramas con epidermis con (T1), y sin heridas (T2), y ramas con peridermis con (T3) y sin heridas (T4). Cada tratamiento tuvo 10 repeticiones. La unidad experimental en Ti y T2 fue una rama (0.5 cm diámetro) menor a un año de edad y en T3 y T4 fue una rama (2.5 cm diámetro) mayor a un año de edad. Las heridas fueron cortes (0.5 cm de longitud) que atravesaron la peridermis y epidermis de las ramas. Previo a la inoculación, las ramas se desinfestaron con hipoclorito de sodio a 1.0 % por 3 min y se lavaron con agua destilada estéril. En cada unidad experimental se colocó un disco micelial (0.5 cm diámetro) de 20 d de edad. En ramas testigo se colocaron discos de PDA sin inoculo. Las ramas experimentales se cubrieron, en los primeros 20 d, con una bolsa de plástico desinfestada con alcohol al 70 % y un algodón estéril humedecido con agua destilada estéril en su interior. La incidencia de muerte descendente por tratamiento se evaluó 30 d y 24 meses después de la inoculación con la ecuación: li = ∑ ni/Ni donde: li = incidencia de ramas enfermas en el momento i; ni = número de ramas enfermas en el momento i; Ni = población total de ramas inoculadas. De tejidos infectados se reaisló al hongo en cultivo puro y se comparó morfológicamente con el hongo inoculado.
Identificación del fitopatógeno
Caracterización morfológica. Una colonia monoconidial de Lasiodiplodia se cultivó en PDA al 2 % (BD Bioxon®) a 25 °C y luz blanca por 20 d e identificó siguiendo las claves taxonómicas para género de Barnett y Hunter (2006) y para especie la referencia y claves de Punithalingam (1976) y Burgess et al. (2006).
Caracterización molecular. El ADN de Lasiodiplodia sp. se extrajo con la técnica de Ahrens y Seemüller (1992). La amplificación de las regiones ITSl e ITS2 de los genes ribosomales (rRNA) se efectuó por PCR usando los iniciadores ITS4 (TCC TCC GCT TAT TGA TAT GC) e ITS 5 (GGA AGT AAA AGT CGT AAC AAG G) (Ahrens y Seemüller, 1992), con la siguiente modificación: agua ultrapura estéril (13.22 μL), solución amortiguadora TBE IX (2.5 μL), MgCl2 a 2.5 mM (2.08 μL), dNTPs a 0.2 mM (2 μL), iniciadores ITS4 e ITS5 a 20 ρmol (2 μL de cada uno), DNA polimerasa (Biogenica®) a 1U (0.2 μL) y muestra problema de ADN a 80 ng (1 mL). El producto amplificado se purificó con el kit Wizard (Promega®) y secuenció con el Genetic Analizer modelo 3100®, Applied Biosystem. Las secuencias obtenidas se analizaron con el software Lasergene® 2001, V5 (DNASTAR®, Inc.) y se alinearon con la base de datos del banco de genes del Nacional Center for Biotechnology Information (NCBI) (www.ncbi.nlm.nih.gov/). Las secuencias con el valor más alto de simila–ridad se consideraron para su comparación con las secuencias obtenidas en este estudio.
Histopatología
De 25 ramas con síntomas de muerte descendente se cortaron en secciones de tejido de 1 cm y se colocaron en solución fijadora FAA [alcohol etílico al 100 % (500 mL), ácido acético glacial (50 mL), formaldehído (100 mL) y agua destilada (350 mL)] por 3 d. El estudio histológico se realizó con las muestras de donde se aisló persistente y exclusivamente a Lasiodiplodia sp. El material se retiró de la solución fijadora, se lavó con agua y se deshidrató e infiltró en un procesador automático de tejidos Tissue–Tek® II, modelo 4640–B® (Sakura Finetechnical Co., LTD. Tokio, Japan). La deshidratación fue gradual con una serie ascendente de alcohol etílico (50 %, 70 %, 96 % y 100 %) (4 h/ alcohol) y la infiltración en etanol absoluto+xileno (1:1), tres veces en xileno y dos veces en parafina (Paraplast marca SIGMA) (4 h/inmersión). El tejido infiltrado se incluyó en paraplast fundido y se dejó solidificar por 24 h. El tejido incluido se montó en un microtomo de desplazamiento modelo Spencer 820® (American Optical Company), se obtuvieron cortes de 10 mm de espesor, se colocaron en un baño de flotación con 3.0 g de grenetina disuelta a 68 °C por 30 s y se montaron en portaobjetos. La técnica de tinción diferencial safranina–verde rápido se realizó con la metodología de Johansen (1940) y Curtis (1986).
RESULTADOS Y DISCUSIÓN
Descripción de síntomas en campo
La enfermedad se observó en árboles mayores de 10 años de edad. Los síntomas en hojas jóvenes de ramas con epidermis fueron: formación de áreas de color rojo, marchitez, necrosis marginal, enrollamiento hacia el haz, muerte y caída prematura. El ápice de la rama mostró marchitez, necrosis y muerte. La enfermedad descendió a la base de la rama anual e invadió ramas secundarias. Los frutos se momificaron, ennegrecieron y quedaron adheridos a la rama enferma. La enfermedad puede causar la muerte de la mayoría de las ramas o incluso del árbol. El tejido interno de las ramas enfermas fue de consistencia corchosa y con necrosis.
Etiología
Hongos aislados
De 125 trozos de tejido de ramas sintomáticas se aisló a Lasiodiplodia sp. (58 %), Pestalotiopsis sp. (21.6 %) y Fusarium sp. (10.4 %). En 10 % de las secciones no hubo crecimiento micelial.
Pruebas de patogenicidad
Fase de laboratorio. La severidad de daños en la pulpa de los frutos, causados por los cinco aislamientos más agresivos de Lasiodiplodia, se indican en el Cuadro 1. Se seleccionó el aislamiento Lasiodiplodia1 como el de mayor agresividad. Lasiodiplodia colonizó la pulpa del fruto solo cuando se inoculó en sitios con herida. El hongo produjo micelio algodonoso gris en la pulpa. En los frutos testigo se observó un área necrótica alrededor del sitio con herida pero no hubo crecimiento micelial.
Patogenicidad en campo. Lasiodiplodia sp. indujo lesiones necróticas de aproximadamente 10 cm longitud×1.0 cm ancho que necrosaron el tejido cortical y profundizaron hacia la médula (Figura 1A) en 20 % de las ramas con peridermis inoculadas por herida a 30 d de la inoculación.
En las ramas testigo se produjo un punto necrótico en el sitio donde se provocó la herida (Figura 1B). A 24 meses de la inoculación, los brotes vegetativos, adyacentes a los sitios inoculados, se marchitaron, necrosaron y murieron en 60 % de las ramas con peridermis inoculadas por herida (Figuras 1C–G). Esta sintomatología fue similar a la que se presentó por infección natural en árboles de mamey en Alpoyeca. Sólo una de las 10 ramas testigo presentaron síntomas, lo cual evidenció una infección latente ya preexistente. Por ausencia de síntomas externos entre el mes 2 y 23 después de la inoculación, no se evaluó la sintomatología interna de muerte descendente. Los estudios sobre enfermedades de ramas de P. sapota son limitados, Álvarez (1997) describió síntomas de muerte descendente, rajadura de corteza y cáncer de tallo en árboles de zapote (P. sapota) en Guatemala y los relacionó a Botryosphaeria sp. e Hypoxylon sp., mientras que Patel y Chauhan (1993) y Balerdi et al. (1996) indicaron que Botryodiplodia theobromae Pat. y Cephaleurus virescens Kunze (alga roja) causan tizón y muerte de brotes jóvenes (retoños) de P. sapota en la India y en los EE.UU.
En el presente estudio, Lasiodiplodia sp. solo causó infección cuando se inoculó directamente sobre el tejido cortical; su hábito de penetración micelial por heridas fue mencionado por Ploetz et al. (1996) y Pavlic et al. (2004). Sin embargo, cuando el hongo colonizó el tejido cortical de las ramas experimentales actuó como un patógeno primario al expresar su potencial de causar enfermedad. La tasa de incidencia en campo posiblemente se limite debido al lento crecimiento e invasión del hongo en el tejido vivo ya que primero debe inducir muerte celular, como lo hace Botryodiplodia theobromae en tejidos leñosos de cítricos (Davis et al., 1987). La muerte descendente de árboles frutales puede resultar de la interacción de L. theobromae con estrés hídrico, suelos arcillosos, escasa materia orgánica, elevada cantidad de caliza activa, fertilización inadecuada, exceso de riego, alta humedad por sombra intensiva, quemadura de sol, presencia de heridas y suelos compactados que complican el desarrollo radicular, nutrición y aireación (Reckhaus y Adamou, 1987; Agustí, 2003).
Los árboles de mamey en Alpoyeca pueden ser susceptibles a la enfermedad por factores como presencia de heridas en ramas por causas naturales o por el uso de herramientas de corte en la cosecha; alcalinidad del suelo (pH 7–6–8.0) y desbalance hídrico y nutricional. En la zona de estudio, L. theobromae libera la mayor cantidad de esporas asexuales de junio a noviembre y es posible que se dispersen por las gotas de lluvias a distancias cortas y por el aire a mayores distancias, como sucede en cítricos (Ko et al, 2004). La secadera de ramas sobre la producción de mamey es significativa porque disminuye la cantidad de frutos y se reduce la longevidad de los huertos comerciales, como sucede en cítricos (Agustí, 2003).
En el presente estudio no se verificó la patogenicidad de Pestalotiopsis sp. debido a su hábito endófito en los árboles de mamey en Alpoyeca. Fusarium sp. no mostró capacidad patogénica cuando se inoculó en tallos con heridas de plántulas de mamey.
Identificación morfológica y molecular del hongo fitopatógeno
La colonia en PDA a 30 d de incubación fue café oscuro con micelio aéreo. Picnidios ostiolados solos o agregados en tejido estromático. Conidios inmaduros amerosporas, hialinos, elipsoides, granulosos, grosor de pared <2 μm y base truncada. Conidios maduros didimosporas de color café oscuro, elipsoides de 19.44–26.73×10.21–13–37 mm (promedio 23–07×11.84 mm), estrías longitudinales y base truncada. Relación largo/ancho de los conidios 1.95:1– Las características de este aislamiento coincidieron con las de L. theobromae reportadas por Punithalingam (1976) y Burgess et al. (2006). El análisis molecular confirmó a L. theobromae como agente casual de muerte descendente. La secuencia genética de este hongo se homologó en 98 % con L. theobromae depositado en el banco de genes (NCBI) con número de acceso EF622073 (542 pb). La secuencia genética de L. theobromae patogénico en esta investigación se depositó en el NCBI con número de acceso EU564805 (540 pb).
Histopatología
Descripción anatómica de ramas asintomáticas
Peridermis con células ovales, parenquimatosas, paredes anticlinales y periclinales engrosadas, lignificadas, con puntuaciones y polifenoles (Figuras 2A–B). Peridermis con lenticelas (Figura 2A). Corteza con células rectangulares y parenquimatosas y células isodiamétricas con polifenoles. Estrato externo de la corteza con abundantes esclereidas y estrato interno con abundantes laticíferos (Figura 2A). Entre la corteza y floema se observó un estrato discontinuo de fibras externas del floema (Figuras 2A y C). Floema con células dispuestas en planos axial y radial. Plano axial con tubos cribosos con células acompañantes; grupos de fibras con células poligonales y esclerenquimatosas con paredes gruesas, lignificadas y células parenquimatosas con polifenoles (Figuras 2A, D y E). Piano radial con células meristemáticas con polifenoles. Cambium vascular con células parenquimatosas dando origen al floema y xilema secundario (Figura 2A). Xilema con un sistema axial y radial. En el primero hubo traqueidas, vasos, fibras y células parenquimatosas (Figura 2F). El sistema radial se conformó de células parenquimatosas con polifenoles. Médula con células parenquimatosas isodiamétricas, algunas con polifenoles, y abundantes laticíferos (Figura 2G).
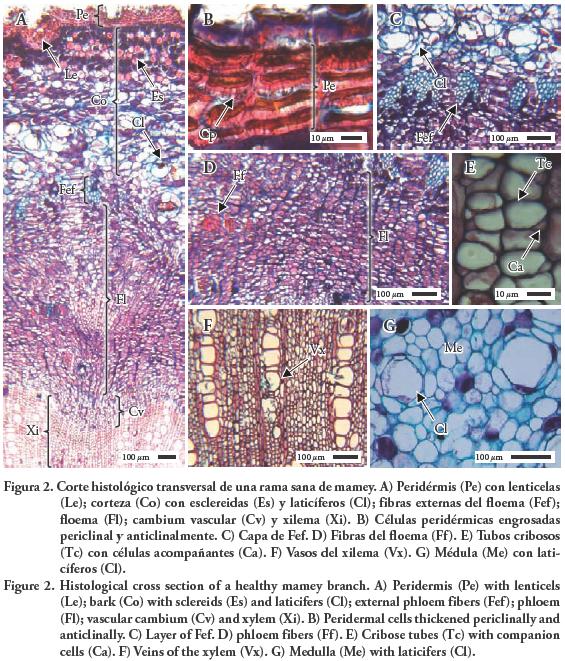
Descripción anatómica de ramas sintomáticas
El tejido peridérmico se fracturó, sin embargo, las células conservaron su morfología (Figuras 3A y B). Las células de la corteza inicialmente se colapsaron y necrosaron; el avance de la enfermedad ocasionó colapso y necrosis de laticíferos; las esclereidas conservaron su forma (Figuras 3B y C). Las fibras externas del floema se colapsaron (Figura 3D). Las células del floema secundario se colapsaron y necrosaron (Figura 3A) y conforme la necrosis avanzó, se formaron cristales rectangulares y precipitados granulosos (Figuras 3E y F). En etapas finales de la enfermedad el floema secundario aumentó y la corteza sufrió necrosis por lo que la mayor parte del floema lo ocuparon los radios dilatados cuyas células parenquimatosas sufrieron esclerenquimatización. Lo mismo ocurrió en el parénquima axil por lo que se formaron grupos de esclereidas (Figuras 3A y E) en la parte interna del floema funcional. El cambium vascular colapso y exhibió necrosis (Figura 3A). El tejido xilemático conservó su forma y estructura, no se observó obstrucción por oclusiones o sustancias ergásticas en el interior de los vasos. La médula mantuvo las características descritas para el tejido asintomático. No se encontraron antecedentes de estudios histológicos en mamey.
Atia et al. (2003) observaron algunos síntomas microscópicos similares a los presentados en ramas de mamey en ramas de uva (Vitis vinifera L.) infectadas con L. theobromae. Milholland (1970) mencionó que este tipo de hongos crecen y avanzan lentamente en el tejido vivo del hospedante debido a su carácter saprofítico porque primero debe inducir muerte celular. En cítricos la enfermedad conocida como muerte súbita causa síntomas similares a los descritos en el presente estudio: colapso y necrosis de los tubos cribosos y las células acompañantes del floema funcional, presencia de cristales romboédricos y lignificación en células parenquimatosas del floema. Estos antecedentes sugieren que el colapso y necrosis de los elementos cribosos del floema pueden impedir el flujo de fotoasimilados, y subsecuentemente, provocar la muerte de ramas por inanición como sucede en árboles de cítricos con daños en el floema debido al ataque del virus tristeza de los cítricos (Román et al, 2004).
CONCLUSIONES
En México, este estudio es el primero que determina a Lasiodiplodia theobromae como agente causal de la muerte descendente del mamey (P. sapota).
Lasiodiplodia theobromae produjo lesiones necróticas internas y externas en ramas de mamey con crecimiento secundario (peridermis) inoculadas por heridas. El hongo indujo muerte descendente en brotes vegetativos; otros daños anatómicos internos fueron rompimiento de peridermis, colapso y necrosis de la corteza, fibras externas, floema funcional y laticíferos; formación de cristales y precipitados granulosos en corteza y floema. El hongo no mostró capacidad de penetración directa, lo cual confirma su parasitismo débil.
LITERATURA CITADA
Agustí, M. 2003. Citricultura. Mundi–Prensa. Madrid, España. 422 p. [ Links ]
Ahrens, U., and E. Seemüller. 1992. Detection de DNA of plant pathogenic mycoplasmalike organisms by polymerase chain reaction that amplifies a sequence of the 16S rRNA gene. Phytopathology 82: 828–832. [ Links ]
Álvarez V., G. 1997. La muerte descendente y el cáncer del tallo en el zapote. Tikalia 15: 37–46. [ Links ]
Atia, M. M. M., A. Z. Aly, M. R. A. Tohamy, H. El–Shimy and M. A. Kamhawy. 2003. Histopathological studies on grapevine die–back. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz 110: 131–142. [ Links ]
Balerdi, C. F., J. H. Crane, and I. Maguire. 1996. Mamey sapote growing in the Florida home landscape. Horticultural Sciences Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida, http://edis.ifas.ufl.edu/pdffiles/MG/MG33100.pdf (Consultado: abril de 2008). [ Links ]
Barnett, L. H., and B. B. Hunter. 2006. Illustrated Genera of Imperfect Fungi. Fourth Edition. Am. Phytopathol. Soc. St. Paul, Minnesota, USA. 218 p. [ Links ]
Burgess, T. I., P. A. Barber, S. Mohali, G. Pegg, W. de Beer, and M. J. Wingfield. 2006. Three new Lasiodiplodia spp. from the tropics, recognized based on DNA sequence comparisons and morphology. Mycologia 98: 423–435. [ Links ]
Curtis P., J. 1986. Microtécnica Vegetal. Trillas, México. 106 p. [ Links ]
Davis, R. M., C. J. Farrald, and D. Davila. 1987. Botryodiplodia trunk lesions in Texas citrus. Plant Dis. 71: 848–849. [ Links ]
INEGI (Instituto Nacional de Estadística Geografía e Informática). 2006. Anuario Estadístico de Guerrero. México. 383 p. [ Links ]
Johansen, D. A. 1940. Plant Microtechnique. McGraw Hill, New York, USA. 503 p. [ Links ]
Ko, W. H., I. T. Wang, and P. J. Ann. 2004. Lasiodiplodia theobromae as a causal agent of Kumquat dieback in Taiwan. Plant Dis. 88: 1383. [ Links ]
Milholland, R. D. 1970. Histology of Botryosphaeria canker of susceptible and resistant highbush blueberries. Phytopathology 60: 70–74. [ Links ]
NCBI (National Center for Biotechnology Information). 2008. http://www.ncbi.nlm.nih.gov/ (Consultado: abril de 2008). [ Links ]
Patel, P. B., and H. L. Chauhan. 1993. Twig blight of sapota caused by Botryodiplodia theobromae Pat.: a new record. Indian J. Mycol. and Plant Pathol. 23: 219. [ Links ]
Pavlic, D., B. Slippers, T. A. Coutinho, M. Gryenhout, and M. J. Wingfield. 2004. Lasiodiplodia gonubiensis sp. nov., a new Botryosphaeria anamorph from native Syzygium cordatum in South Africa. Studies in Mycol. 50: 313–322. [ Links ]
Ploetz, R. C., D. Benscher, A. Vazquez, A. Colls, J. Nagel, and B. Schaffer. 1996. A reexamination of mango decline in Florida. Plant Dis. 80: 664–668. [ Links ]
Punithalingam, E. 1976. Botryodiplodia theobromae. CMI description of pathogenic fungi and bacteria. No. 519. Commonwealth Mycological Institute. Kew, Surrey, England. [ Links ]
Reckhaus, P., and I. Adamou. 1987. Hendersonula dieback of mango in Niger. Plant Dis. 71: 1045. [ Links ]
Román M., P., M. Cambra, J. Juárez, P. Moreno, N. Durán–Vila, F. A. O. Tanaka, E. Alves, E. W. Kitajina, P. T. Yamamoto, R. B. Bassanezi, D. C. Teixera, W. C. Jesus Junior, A. J. Ayres, N. Gimenes–Fernandes, F. Rabenstein, L. F. Girotto, and J. M. Bové. 2004. Sudden death of citrus in Brazil: A graft–transmissible bud union disease. Plant Dis. 88: 453–467. [ Links ]
SIAP (Servicio de Información y estadística Agroalimentaria y Pesquera). 2006. Anuario Estadístico de la Producción Agrícola de los Estados Unidos Mexicanos. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación, http://www.siap.gob.mx/ (Consultado: abril de 2008). [ Links ]
Zamora M., T., E. Cárdenas S., J. F. Cajuste B., y M. T. Colinas L. 2001. Anatomía del daño por rozamiento y por Colletotrichum gloeosporioides Penz. en frutos de aguacate 'Hass'. Agrociencia 35: 237–244. [ Links ]

