










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgrociencia
versão On-line ISSN 2521-9766versão impressa ISSN 1405-3195
Agrociencia vol.43 no.1 Texcoco Jan./Fev. 2009
Recursos naturales renovables
Site index and soil properties in young plantations of Eucalyptus grandis and E. urophylla in Southeastern Mexico
Índice de sitio y propiedades del suelo en plantaciones jóvenes de Eucalyptus grandis y E. urophylla en el sureste de México
Carina E. Delgado–Caballero1, Armando Gómez–Guerrero1*, José R. Valdez–Lazalde1, Hector De los Santos–Posadas1, Aurelio M. Fierros–González1, William R. Horwath2
1 Forestal, Campus Montecillo. Colegio de Postgraduados. Km. 36.5 Carr. México–Texcoco. 56230. Montecillo, Estado de México. *Author for correspondence: (agomez@colpos.mx).
2 University of California, Davis. (wrhorwath@usedavis.edu)
Received: April, 2007.
Aprobed: October, 2008.
Abstract
Research on the ability of tropical soils to support fast–growing forests species is required to optimize, preserve and select lands for timber production. In this work site index (SI) of Eucalyptus grandis and E. urophylla plantations in Southeastern Mexico, was related to chemical and physical soil properties. Based on a joint model of SI vs. sand content, the fit suggests that SI improves as the sand content increases (p<0.01). Sand content of 28% in the surface soil (0–5 cm) and 24% in the sub–surface soil (5–30 cm) is a desired minimum for establishing highly productive stands. The best SI conditions were found on sandy soils with a sand content of 65 %. Other variables, such as soil bulk density, N mineralization and organic matter were also measured but were not correlated with stand growth or SI. Cation exchange capacity was inversely related to tree growth, which was explained as an indirect relationship since soil with higher content of fine soil particles (clay + silt ) were less productive.
Key words: Forest plantations, forest soils, site productivity, soil physical properties.
Resumen
La investigación sobre la capacidad de los suelos tropicales de favorecer el crecimiento rápido de especies forestales es necesaria para optimizar, preservar y seleccionar tierras para la producción de madera. En este trabajo el índice de sitio (SI) de las plantaciones de Eucalyptus grandis y E. urophylla en el sureste de México se asoció a propiedades químicas y físicas. A partir de un modelo conjunto de SI vs. contenido de arena, el ajuste indica que SI mejora mientras el contenido de arena aumenta (p<0.01). El contenido de arena de 28% en la superficie del suelo (0–5 cm) y de 24% en la capa subyacente del suelo (5–30 cm) es el mínimo deseado para establecer rodales de alta productividad. Las mejores condiciones de SI se encontraron en suelos arenosos con un contenido de arena de 65%. Se midieron también otras variables, tales como densidad aparente del suelo, mineralización de N y materia orgánica, pero no se correlacionaron con el crecimiento rodal o SI. La capacidad de intercambio catiónico estuvo inversamente relacionada al crecimiento del árbol, lo cual se explicó como una relación indirecta ya que los suelos con mayor contenido de partículas finas del suelo (arcilla+limo) fueron menos productivos.
Palabras clave: Plantaciones forestales, suelos forestales, productividad del sitio, propiedades físicas del suelo.
INTRODUCTION
Site index (SI) refers to the height of dominant trees in a stand at a specific age and has been a useful criterion for identifying sites with high potential for tree growth (Spurr and Barnes, 1980; Daniel et al., 1982; Clutter et al., 1983;). SI can be directly estimated only in wooded areas; however, relating SI with soil properties is a useful approach to predict forest productivity in lands without trees (Carmean, 1975). Screening sites by soil productivity is a useful method to effectively use the potential of each site and plan forestry activities in a sustainable way (Mckenney and Pedlar, 2003). Studies of productivity in forest plantations are also important as their establishment in lands not previously used for forestry purposes contributes to sequestering carbon by transferring atmospheric CO2 to the plant and soil system (Zinn et al., 2002; Forrester et al., 2006; Lima et al., 2006). Lemma et al. (2006) found that a 20–year afforestation site with Eucalyptus grandis in lands previously used for agricultural purposes returned the levels of soil organic carbon content to that of the native forests. This work focused on finding simple equations to relate SI of E. grandis and E. urophylla with soil properties in plantations established in one of the regions with the highest potential for establishing tropical plantations in México, in order to predict productivity in lands to be planted in the future.
MATERIALS AND METHODS
The study area is located at 17° 07'–17° 54' N and 94° 52'–95° 54' W around the boundary of Oaxaca and Veracruz states (Figure 1). The dominant climate is tropical wet (Aw) with mean annual temperature of 27 °C and mean annual precipitation between 1500 to 2400 mm. Mean elevation is 150 m. Soil types in the area belong to the orders of Alfisols and Ultisols (Soil Survey Staff, 1997). A summary of the soil characteristics in the study sites is presented in Table 1.
Plantations description
The establishment of the plantations under investigation started in 1998 with E. grandis and E. urophylla on lands used for grazing in the last 40 years. The native vegetation in the area was tropical forest, but an intense program promoted by the Mexican government encouraged land use change in the 1960's and 1970's (Masera et al., 1992; Paz, 1995). Plantations were 28 to 77 months–old at the beginning of the study and were planted 3 m apart in a square grid fashion. Plantations were fertilized with 60–60–30 kg ha–1 for NPK three times a year during the two first years of establishment. Competition removal was performed during the first three months; afterwards, the establishment of the plantation is not at risk by plant competition.
Soil sampling
At the PLANTEH project, 154 500–m2–circle permanent plots were established in 2004 and the dasometric data was used for the tree growth and yield study reported by Gómez et al. (2006). In 2006 forty five of the 154 plots were re–measured and every plot was divided into four quadrants where soil samples were obtained from 0–5, 5–15 and 15–30 cm depth. Samples from the four quadrants were used to make a composite sample for laboratory soil analyses. Additionally, soil bulk density samples were taken at every quadrant at the same soil depths using a core sampler with hammer as illustrated in Figure 2. Since soil bulk density is more variable at the soil surface, an additional sample was taken for the first soil depth. In our study, 720 soil samples for bulk density were taken.
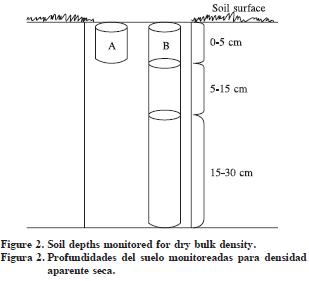
Soil samples were air dried and sieved through a 2–mm mesh. Soil analyses were: pH, organic matter (Walkley–Black), soil texture (Bouyoucos), cation exchange capacity (amonium acetate, pH 7) (Rowell, 1994) and an anaerobic N–mineralization (Powers, 1980).
Site index estimates
The dominant height models used to obtain the SI estimates at each plot (Gómez et al., 2008) were:
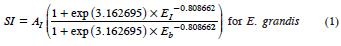
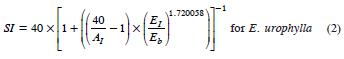
where SI is site index; AI = dominant height at the EI age of 80 months.
These equations are based on a Hossfeld IV model (Cieszewski, 2001; Kiviste et al., 2002). More details for the conditions of fit and validation of these models can be found in Gómez et al. (2006). The SI estimates were separated into 3 m intervals.
Statistical analysis
Soil variables were initially correlated to SI and tree growth using PROC CORR (SAS, 1998). Soil variables were also plotted against SI to identify patterns and trends. A simple trend between sand content and SI was described on all soil depths by the following segmented model:

where S = sand content in %; j = inflection point at which IS = a0 becomes SI = α0 + α0 (j – S)2 ; Ij = the indicator variable equal to 1 for S>j, zero otherwise.
The model has the advantage that the inflection point is treated as a parameter and has to be estimated using non–linear regression. This parameter represents the lowest acceptable sand content before SI increases a function of the sand content. Segmented models help to simplify prediction equations with the minimum number of variables.
Marginal increments for SI after the inflection (j) can be calculated as:
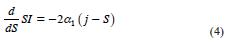
The model was fitted using PROC MODEL (SAS, 1998). Multiple regression was also used; however, adding more than one variable did not improve significantly the explanation of IS variation, as the segmented model did with one variable. Although using only one variable is a simple approach, the advantage is the easier context in the interpretation.
RESULTS AND DISCUSSION
Tree growth parameters for the 45 study sites are summarized in Table 2. Range for tree age was between 24 to 73 months and for SI between 16 to 38 m at the base age of 80 months. Basal area varied from 2 to 26 m2 ha–1, and stand volume from 8 to 229 m3 ha–1. Current increments in stem ranged from 0.3 a 6.8 m–3 month , that is 3.6 to 81.6 m3 ha–1 year–1.
Soil properties and tree growth relationships are summarized in Tables 3 to 5.(Table 3, 4 and 5). The soil variable more correlated to SI was soil texture, correlation was positive for larger soil particles and negative for fine soil particles like silt and clay. This trend was found at three soil depths. Cation exchange capacity (CEC) was negatively related to the SI and periodic increment (PI). Organic matter and CEC showed a significant correlation (0.64), only at 0–5 cm soil depth. The highest correlation was between CEC and it was negative for the sand component and positive for finer soil particles like silt and clay; this strong correlation was held for the three soil depths. Dry bulk density (DBD) and N mineralization were not significantly related to tree growth.
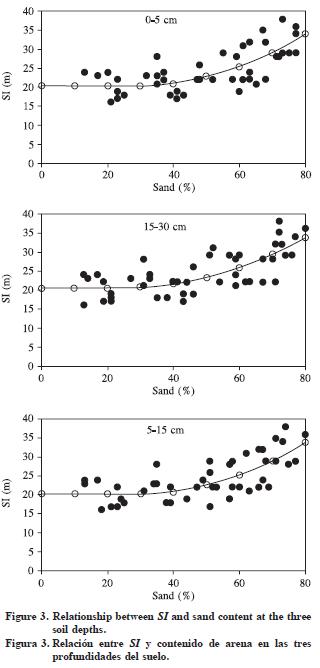
The fit for equation 3 on each depth is shown in Table 6. All parameters in the model are statistically significant including the inflection point j. When testing for differences between species, no statistical differences were found; therefore, a single equation per depth was used. The relationship between SI and sand content explained 51 to 53% of the SI variation (Figure 3).
The wide range of the values found for stand volume, basal area and periodic increments is a consequence of the variation in age of the plantations. However, differences in SI indicate variation in soil productivity at the sites sampled. The highest level of productivity found in this study was 81 m3 ha–1 year–1 which is a productivity as high as that reported in very productive sites of Brazil. For example, Stape et al. (2006) reported productivities of 62 m3 ha–1 year–1 for intensive managed eucalyptus plantations.
Productivity is influenced by water availability, and eucalyptus plantations with high water use efficiency yield up to 30 m3 ha–1 year–1 in sites with 1600 mm of annual precipitation (Stape et al., 2004). In the study area the average annual precipitation is 2000 mm, which explains the higher productivity levels. Using wood stem density data and allometric equations for branch and leaf components from the studied plantations, aboveground net primary productivity was estimated to be around 3200 g m2 year–1, which is a high level of productivity as compared to other forest communities (Lieth, 1975; Powers, 1999).
Sand content was one of the most important variables related to SI variation, a relationship that may be explained by the soil aeration regime which is improved with higher contents of coarse soil particles. The relationship between sand and SI fit followed the same pattern when the analysis was performed by soil depth. This is a consistent result that indicates that sandy texture in the soil surface is a critical condition for tree growth on these sites. According to the parameters of equation (3), the amount of sand in the 0–5 cm soil depth starts influencing SI at sand contents around 28%, with marginal increments from 0.016 to 0.67 m per percent unit of sand content. The influence of the 15–30 cm sand particles on tree growth stars at 24% of sand content with marginal increments from 0.04 to 0.64 per percent unit of sand content. The difference in sand content suggests that the best sites for tree growth require coarse textures. The selection of the main variable related to IS was a simple process of correlation analysis, but the segmented model explained as much variation as multiple linear models including more than one variable. Therefore, the simple one–variable model was preferred for the purposes of our study.
From a soil physical perspective, the sites with better drainage in the upper 30 cm soil layer are more suitable for eucalyptus plantations. The relatively flat topography and the high precipitation in the area may lead to a poor aeration regime so that soil texture becomes an important factor influencing soil productivity. Our results are consistent with those of Aparicio and López (1995) and Gonçalves et al. (1990) who reported similar results for E. grandis in Brazil and Chile. It is important to note that some prediction models assume a bell shape to predict tree growth (Spurr and Barnes, 1980), indicating that forest productivity in extreme soil texture conditions (very clayey or very sandy) lead to lower rates in tree growth. Nonetheless, to explain more accurately the soil texture influence in forest productivity, changes in the soil moisture and soil air regime should be taken into account (Gomez et al., 2002).
The lack of relationship DBD and forest productivity was unexpected. In general, soils with higher DBD are less productive due to the biological implications for root growth (Fisher and Binkley, 2000). Goncalvez et al. (1997) found that the productivity of E. camaldulensis was more than three times higher when DBD changed from 1.25 to 1.06 Mg m–3 in a Inceptisol in Brazil. Correlation between soil texture and DBD was low for all soil depths, with the highest value of 0.38 at the 0–5 cm depth (Table 6). The lack of correlation between DBD and soil texture maybe due to differences in soil structure. Although there is a general correlation between DBD and soil particle distribution, the level of organization of the soil which defines its structure may obscure a direct relationship between these soil variables (Rowell, 1994).
DBD values found for the 0–5 cm soil depth varied from 0.82 to 1.24 Mg m–3. For the deeper soil layers values were 1.47 and 1.57 Mg m3– in the 5–15 and the 15–30 cm depths. DBD in the first 5 cm is favorable for tree root growth, but the higher values at the 5– 10 and 15–30 cm soil depths were expected to have some effect on tree growth as soil porosity is reduced and soil strength increases (Fisher and Binkley, 2000). However, some eucalyptus plantations seem to perform well in Oxisols, Ultisols and Entisols with DBD values up to 1.6 mg m–3, as long as plants are not water stressed (Stape et al., 2004). One explanation for the lack of association between tree growth and DBD in this study is the fact that the soil is relatively wet all year. DBD influences tree root growth especially when decreases in soil moisture lead to increases in soil strength (Gomez et al., 2002). Although DBD showed values relatively high in the subsoil, soil strength did not reach critical levels to impair root growth and hence productivity.
The poor correlation between tree growth (SI) and N mineralization and soil organic matter is another unexpected result. The availability of N is an index that generally is related to tree growth as N is the most limiting nutrient in plant communities (Fisher and Binkley, 2000). Ranges of N mineralization for the soil depths 0–5, 5–15, 15–30 cm were 13–104, 8–66 and 4–47 mg kg–1, values which, according to Powers (1980), are influenced by total soil N and are common for forest soils. The poor relationship between N mineralization and SI may be explained by the fact that this is the first rotation for the plantation and soils were cropped with grasses for 40 years previous to establishment. The organic matter accumulated in the soil surface during that time may have contributed to maintaining adequate levels of organic matter and available nitrogen. Soil organic matter in the soil surface ranged from 0.5 to 4% and soil N from 0.05 to 0.34% in the upper 30 cm. The availability of N is influenced by the total amount of N and organic matter. It is very likely that the pools of organic carbon in the soil led to an adequate provision of N.
Eucalyptus is highly efficient at recycling nutrients (Stape et al., 2004) and the correlation with N mineralization will be evident as N status of the plantations is expected to decrease in subsequent rotations. Nitrogen is important and mixed plantations of Eucalyptus and leguminous species show higher productivity than monoculture (Forrester et al., 2006). Even removal of leguminous species in the herbaceous understory of Eucalyptus stands can lead to a deficit of 140 kg–1 N ha–1 seven years after of the establishment of the plantation (Laclau et al., 2005). Since monoculture is not a sustainable system after the first rotation, we speculate that successive rotations in the study sites will be more N limited, leading to a better relationship between tree growth and soil N availability. Coarse–textured soils with good nutrient status seem to provide the best environments for the productivity of eucalyptus. This trend was also seen by Stape et al. (2004) who reported that coarse–textured soils were twice as responsive to fertilization as sites with finer soil texture.
CONCLUSIONS
Site index (SI) of E. grandis and E. urophylla were related to soil texture. Sand contents higher than 28% at 0–5 cm soil depth positively influenced tree growth. In deeper layers (5–30 cm) this influence occurred at sand contents higher than 24%. Surface soil sand content explained 51 to 53% of SI variation. No correlation was found between soil bulk density measured at 0–5, 10–15 and 15–30 cm and tree growth (SI). Adequate soil moisture levels may prevent the soil from reaching soil strength values that would impair root growth. Similarly, neither N mineralization nor organic matter content were correlated to soil productivity, which may be explained by the fact that in these soils there were grasses for at least 40 years before the plantations were established, providing enough N to the system. Nevertheless, subsequent rotations may experience N shortages depending on the management system applied to the plantations. To go further with this study two things are needed. The first one is spatial information of soil texture of surface soil and the second one is to establish an experiment to confirm that forest productivity can be predicted from soil texture in new plots.
ACKNOWLEDGEMENTS
This work was sponsored by the National Council for Science and Technology (CONACYT) and the National Forestry Commission of Mexico (CONAFOR). Project: 2003 CO3–9925.
LITERATURE CITED
Aparicio, J. L., y J. A. López. 1995. Potencial de Eucalyptus grandis en el sudeste de la provincia de Corrientes y algunos factores edáficos relacionados con la producción de madera. Bosque 16: 81–89. [ Links ]
Carmean, W. H. 1975. Forest site quality evaluation in the United States. Adv. Agron. 27: 209–269. [ Links ]
Cieszewski, C. J. 2001. Comparing fixed–and variable–base–age site equations having single versus multiple asymptotes. For. Sci. 48: 7–23. [ Links ]
Clutter, L. J., C. J. Fortson, V. L. Pienaar, and L. R. Bailey. 1983. Timber Management. A Quantitative Approach. New York Wiley. 333 p. [ Links ]
Daniel, P. W., U. E. Helms, y F. S. Baker. 1982. Principios de Silvicultura. Editorial McGraw Hill. México. 492 p. [ Links ]
Fisher, R. F., and D. Binkley. 2000. Ecology and Management of Forest Soils. 3rd ed. Wiley. USA. 489 p. [ Links ]
Forrester, D. I., J. Bauhus, A. L. Cowie, P. A. Mitchell, and J. Brockwell. 2006. Productivity of three young mixed–species plantations containing N 2 –fixing Acacia and non–N2–fixing Eucalyptus and Pinus trees in south–eastern Australia. For. Ecol. Manage. 233: 275–284. [ Links ]
Gomez, A., R. F. Powers, M. J. Singer, and W. R. Horwath. 2002. Soil compaction effects on growth of young ponderosa pine following litter removal in California's Sierra Nevada. Soil Sci. Soc. Am. J. 66: 1334–1343. [ Links ]
Gómez, T. J., H. De los Santos P., y A. M. Fierros G. 2008. Modelos de crecimiento en altura dominante para Eucalyptus grandis y E. urophylla en María Lombardo, Oaxaca, México. In Press. Fitotec. Mex. [ Links ]
Gonçalves, J. L. M., J. L. I. Demattê, J., and H. T. Z. do Couto. 1990. Relacoes entre productividade sitios florestais de Eucalyptus grandis e Eucalyptus saligna com as propriedades de alguns solo de textura arenosa e media no Estado de Sau Paulo. IPEF. Piracicaba. 43/44: 24–39. [ Links ]
Gonçalves, J. L. M., N. F. Barros, E. K. S. Nambiar, and R. F. Novais. 1997. Soil and Stand magement for short–rotation plantations. In: Nambiar, E. K. S., and A. G. Brown (eds). Management of Soil, Nutrients, and Water in Tropical Plantation Forester, ACIAR monograph Canberra 43: 397–417. [ Links ]
Kiviste, A., J., A. Alboreca R., y D. R. González A., 2002. Funciones de Crecimiento en el Ámbito Forestal. Comunicaciones INIA. Serie: Recursos Naturales. No. 4. 189 p. [ Links ]
Laclau, J. P., J. Ranger, P. Deleporte, Y. Nouvellon, L. Saint–Adre, S. Martel, and J. P. Bouillet. 2005. Nutrient cycling in a clonal stand of Eucalyptus and an adjacent savanna ecosystem in Congo. 3. Input–output budgets and consequences for the sustainability of plantations. For. Ecol. Manage. 210: 375–395. [ Links ]
Lemma, B., D. B. Kleja, I. Nilsson, and M. Olsson. 2006. Soil carbon sequestration under different exotic tree species in the southwestern highlands of Ethiopia. Geoderma 136: 886–898. [ Links ]
Lieth, H. 1975. Modeling the primary productivity of world. In: Lieth H., and R. H. Wihittaker (eds). Primary Productivity of the Biosphere. Springer Verlagi, New York. pp: 237–263. [ Links ]
Lima, A. M. N., I. R. Silva, J. C. L. Neves, R. F. Novais, N. F. Barros, E. S. Mendonza, T. J. Smyth, M. S. Moreira, and F. P. Leite. 2006. Soil organic carbon dynamics following afforestation of degraded pastures with eucalyptus in southeastern Brazil. For. Ecol. Manage. 235: 219–231. [ Links ]
Masera, O., M. J. Ordóñez, y R. Dirzo. 1992. Emisiones de carbón a partir de la deforestación en México. CIENCIA: Revista de la academia de la investigación científica 43: 151 p. [ Links ]
McKenney, D. W., and J. H. Pedlar. 2003. Spatial models of site index based on climate and soil properties for two boreal tree species in Ontario, Canada. For. Ecol. Manage. 175: 497–507. [ Links ]
Paz, M. F. 1995. Selvas tropicales y deforestación. Apuntes para la historia reciente del trópico húmedo mexicano. In: Paz, M. F. (Coord). De Bosques y Gente. Aspectos Sociales de la Deforestación en América Latina. Cuernavaca, Morelos. CRIM, UNAM. pp: 53–87. [ Links ]
Powers, R. F. 1980. Mineralizable soil nitrogen as an index of soil nitrogen availability to forest trees. Soil Sci. Soc. Am. J. 44: 1314–1320. [ Links ]
Powers, R. F. 1999. On the sustainability productivity of planed forest. New Forest 17: 253–306. [ Links ]
Rowell, D. L. 1994. Soil Science, Methods and Applications. Longman Scientific and Technical. UK. 350 p. [ Links ]
SAS, Institute 1998. SAS User Guide. SAS Institute Inc., Cary, NC. 595 p. [ Links ]
Soil Survey Staff, U.S.D.A. 1997. Keys to Soil Taxonomy. 7th,/Ed. Soil Conservation Service. U. S. Department of Agriculture, Blacksburg, Virginia, USA. 545 p. [ Links ]
Spurr, S. H., and B. V. Barnes. 1980. Forest Ecology. John Wiley and Sons, Inc. 687 p. [ Links ]
Stape, J. L., D. Binkley, S. Jacob, and E. N. Takahashi. 2006. A twin–plot approach to determine nutrient limitation and potential productivity in Eucalyptus plantations at landscape scales in Brazil. For. Ecol. Manage. 223: 358–362. [ Links ]
Stape, J. L., D. Binkley, and M. G. Ryan. 2004. Eucalyptus production and the supply, use and efficiency of use of water, light and nitrogen across a geographic gradient in Brazil. For. Ecol. Manage. 193: 17–31. [ Links ]
Zinn Y. L., D. V. S. Resck, and J. E. Silva. 2002. Soil organic carbon as affected by afforestation with Eucalyptus and Pinus in the Cerrado region of Brazil. For. Ecol. Manage. 166: 285–294. [ Links ]

