










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgrociencia
versão On-line ISSN 2521-9766versão impressa ISSN 1405-3195
Agrociencia vol.42 no.5 Texcoco Jul./Ago. 2008
Fitociencia
Sucrose application suppresses infection of the parasitic plant Orobanche ramosa in tomato (Lycopersicon esculentum)
La aplicación de sacarosa suprime la infección de la planta parásita Orobanche ramosa en tomate (Lycopersicon esculentum)
Clara I. González–Verdejo1*, Miguel A. Dita2, Salvador Nadal1, M. Teresa Moreno1 and Belen Román1
1 IFAPA, Centro Alameda del Obispo, Área de Mejora y Biotecnología, Apdo. 3092, 14080 Córdoba, Spain *Author for correspondence: (clara.gonzalez.ext@juntadeandalucia.es).
2 EMBRAPA, Mandioca e Fruticultura Tropical, Cruz Das Almas, 44380–000. Bahia, Brasil.
Received: July, 2007.
Aproved: June, 2008.
Abstract
Orobanche ramosa is an achlorophyllous holoparasitic weed that causes devastating losses in many commercial crops. An experiment was made using sucrose to evaluate its effect on the early growth stages of the parasite. Although O. ramosa germination was not statistically affected by sucrose, the radicle length, the number of attachments sites and the nodules formed decreased significantly.
Keywords: Lycopersicon sculentum, growth regulation, parasitism, sugar.
Resumen
Orobanche ramosa es una maleza aclorófila haloparásita que ocasiona pérdidas devastadoras en muchos cultivos comerciales. Se realizó un experimento con sacarosa para evaluar su efecto en las etapas tempranas de crecimiento del parásito. Aunque la germinación de O. Ramosa no fue estadísticamente afectada por la sacarosa, la longitud radicular, el número de sitios de adhesión y los nódulos desarrollados disminuyeron significativamente.
Palabras clave: Lycopersicon sculentum, regulación del crecimiento, parasitismo, azúcar.
Introduction
Soluble sugars like glucose and sucrose are important regulators of various plant processes (Graham, 1996; Gibson, 2004). In O. ramosa, the addition of sucrose (30 g L–1) to in vitro germinating seeds in the absence of the host, results in dramatic changes in radicle growth pattern (González–Verdejo et al., 2006). This sugar mimics the later stages of infection when the parasite is connected to the host vascular system and obtains its carbohydrate nutrients mostly as sucrose (Aber, 1984). This prompted us to investigate if development alterations caused by exogenous sucrose application takes place when Orobanche grows directly on tomato (Lycopersicon esculentum) roots. In this study, tomato plants infected with O. ramosa were watered with a solution of sucrose (0, 15, 30 and 60 g L–1). Possible alterations could influence the infection process and could lead to control this parasitic plant.
MATERIALS AND METHODS
Tomato seeds (Lycopersicon sculentum cv. Money Maker) were sterilized in 3.5% sodium hypochlorite for 10 min, then rinsed three times with sterile distilled water. O. ramosa seeds were collected from plants which were parasitizing tobacco fields in Granada (Spain) and sterilized 2 h with 0.5% formaldehyde, followed by a 20 min incubation at 50 °C. Finally, seeds were rinsed three times with sterile distilled water ( González–Verdejo et al. 2005).
Assays were performed following the methodology described by Pérez de Luque et al. (2004) with slight modifications: tomato seeds were pre–germinated in vermiculite and then transferred to Petri dishes containing perlite covered by a sheet of glass–fibre filter (Whatman International, Kent, UK); plates had a hole in its upper part to allow for free growth of the aerial part of the plant. Approximately 6 mg of O. ramosa seeds were sown on each sheet of glass–fibre filter with the tomato roots; then the plates were wrapped in aluminium foil to maintain O. ramosa seeds in the dark. Plates were watered and vertically stored in a growth chamber (21 °C; 16 h light). After 10 d of conditioning period, sucrose (Merk 141621) was added to the water (0, 15, 30, and 60 g L–1; treatments): plates without sucrose were used as negative controls.
Four independent plates (each a randomized block), were set for each of the treatments. Percentages of seed germination were determined based on 400 seeds per plate (100 seeds in each section of the four analysis fields established per plate). The relationship between the germination data to root length (germination rate per root. m) was determined by the intercept method of Tennant (1975). The radicle length was determined in 40 seeds per plate (10 seeds in each of the four analysis fields) per treatment. All experiments were repeated three times with similar results.
The experimental design was randomized complete blocks with four treatments. Means were compared using the Tukey test (p<0.05). Percentages of broomrape germination data were transformed with arcsin (√/x /100) before analysis.
RESULTS AND DISCUSSION
O. ramosa seed germination in presence of sucrose
Low and intermediate sucrose concentrations (15 and 30 g L–1) did not reduce the number of germinating O. ramosa seeds, and only the highest concentration (60 g L–1) caused a moderate reduction on the germination rate (Table 1). This has been also observed in Arabidopsis, where 90–100% of seeds treated with high glucose or sucrose showed similar results (Laby et al., 2000).
Effect of sucrose on early seedling development of O. ramosa
Radicle lenght was reduced by sucrose (30 and 60 g L–1); with 60 g L–1 reduction was approximately 60% (Table 1). In our study, differences on seed morphology were also observed (data not shown). The sucrose treatment seemed to increase the number of seedlings with no radicle development showing an apresorium emerging directly from the testa. Therefore, although sucrose did not inhibit the germination, it was effective in a second stage of the seedling development.
O. ramosa infection and tubercles formation in the presence of sucrose
All treatments with sucrose effectively prevented successful seedling establishment and tubercles formation (Table 1; Figure 1). Since 15 g L–1 of sucrose controled nodules development, a wider range around this concentration should be tested.
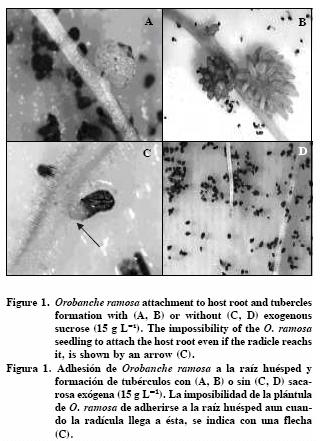
Even when the radicle reached the host root, it was unable to infect it (Figure 1C). This finding is consistent with similar results observed by Ben–Hod et al. (1991) with O. aegyptiaca callouses developed in vitro. Protusions of these callii behaving like radicles of germinating seeds were unable to infect host roots when grown on media containing organic nutrients.
The results, both on seedling development and total amount of attachments, could be attributed to the modified osmotic potential of the media with the sucrose concentrations, rather than to sucrose concentration itself. In studies with the model plant Arabidopsis thaliana, the changes produced by 0.11 M glucose on germinating seeds were not due to osmotic stress (To et al., 2002). We used 15, 30 and 60 g L–1 (0.044, 0.088 and 0.18 M), concentrations lower or similar to those used for Arabidopsis. For this reason, it is improbable that O. ramosa seeds sensitivity to the sugar concentrations tested could be due to the osmotic potential of the media. Further studies should consider sorbitol treatment in the O. ramosa/tomato system in order to neject this hypothesis.
Sucrose effect on host plant
The effect of sucrose on the host plant was tested by determining the root length for each treatment: 5.90, 5.61, 4.55 and 3.47 m for 0, 15, 30 and 60 g L–1. There were significant differences between the 60 g L–1 and 30 g L–1 as well as between the control and the 30 g L–1 concentration (Table 1). Thus, it is highly desirable to determinate the sucrose concentration needed to control O. ramosa without interfering with the host root development.
CONCLUSIONS
Sucrose blocks O. ramosa life cycle by modifying the normal germination step and preventing its attachment to the host plant. Nevertheless, the results of sucrose application could be different under field conditions, where precipitations or soil microbe interaction might alter initial concentrations or interfere with the host development. Studies about different Orobanche species attacking other crops (O. crenata–legumes or O. cumana / sunflower systems) could also be of utmost interest.
ACKNOWLEDGEMENTS
This work was financially supported by the INIA_RTA04–067 project.
LITERATURE CITED
Aber, M. 1984. Cytophysiological aspects of Orobanche crenata Forsk. parasitizing Vicia faba L. Third International Symposium on Parasitic Weeds, Aleppo. [ Links ]
Ben–Hod, G., D. Losner, D. M. Joel, and A. M. Mayer. 1991. Infection of tomato roots using callii of broomrape. Physiologia Plantarum. 83: 353–356. [ Links ]
Gibson, S. I. 2004. Sugar and phytohormone response pathways: navigating a signalling network. J. Exp. Bot. 55: 253–264. [ Links ]
González–Verdejo, C. I., X. Barandiaran L, M. T. Moreno Y, J. I. Cubero S, and A. Di Pietro. 2005. An improved axenic system for studying pre–infection development of the parasitic plant Orobanche ramosa. Annals Bot. 96: 1121–1127. [ Links ]
González–Verdejo, C. I., X. Barandiaran L, M. T. Moreno Y, J. I. Cubero S, and A. Di Pietro. 2006. A peroxidase gene expressed during early developmental stages of the parasitic plant Orobanche ramosa. J. Exp. Bot. 57: 185–192. [ Links ]
Graham, I. A. 1996. Carbohydrate control of gene expression in higher plants. Res. Microbiol. 147: 572–580. [ Links ]
Laby, R. J., M. S. Kincaid, D. Kim, and S. I. Gibson. 2000. The Arabidopsis sugar–insensitive mutants sis4 and sis5 are defective in abscisic acid synthesis and response. The Plant J. 23: 587–596. [ Links ]
Pérez–de–Luque, A., J. V. Jorrín, and D. Rubiales. 2004. Crenate broomrape control in pea by foliar application of benzothiadiazole (BTH). Phytoparasitica. 32: 21–29. [ Links ]
Tennant, D. 1975. A test of a modified line intersect method of estimating root lenght. J. Ecol. 63: 955–1001. [ Links ]
To, J. P. C., W. D. Reiter, and S. I. Gibson. 2002. Mobilization of seed storage lipid by Arabidopsis seedlings is retarded in the presence of exogenous sugars. BMC Plant Biol. 2: 4. [ Links ]

