










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.42 no.3 Texcoco abr./may. 2008
Recursos naturales renovables
Patrón de cruzamiento en poblaciones naturales de Pseudotsuga menziesii (Mirb.) Franco en México
Mating pattern in natural populations of Pseudotsuga menziesii (Mirb.) Franco in Mexico
Jorge Cruz–Nicolás1, J. Jesús Vargas–Hernández1, Porfirio Ramírez–Vallejo2 y Javier López–Upton
1 Forestal y
2 Genética. Campus Montecillo. Colegio de Postgraduados. 56230. Montecillo, Estado de México. (cnjorge@colpos.mx) (vargashj@colpos.mx)
Recibido: Abril, 2007.
Aprobado: Enero, 2008.
Resumen
Las poblaciones mexicanas de Pseudotsuga menziesii (Mirb.) Franco tienen una distribución natural restringida y fragmentada, sobre todo en la región centro; por tanto, hay riesgo de que éstas se pierdan por efectos del cambio climático. Con el propósito de sugerir medidas adecuadas para su conservación, se evaluó el patrón de cruzamiento y su efecto sobre el nivel de endogamia en tres poblaciones naturales de P. menziesii, dos del norte y una del centro de México, con base en el porcentaje de polinización cruzada. Con la información de cinco loci polimórficos se estimaron las frecuencias alélicas de polen y óvulos y la proporción de cruzamiento en cada población usando modelos de loci múltiples 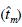 y de un solo locus
y de un solo locus 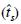 . No se encontraron diferencias en las frecuencias alélicas de polen y de óvulos, lo que indica que el polen proviene de la misma comunidad que los árboles madre. El valor de
. No se encontraron diferencias en las frecuencias alélicas de polen y de óvulos, lo que indica que el polen proviene de la misma comunidad que los árboles madre. El valor de 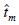 en las dos poblaciones del norte fue cercano a 90%; en la población del centro fue 49% como resultado de un alto porcentaje de autofecundación. La similitud de las estimaciones del grado de cruzamiento
en las dos poblaciones del norte fue cercano a 90%; en la población del centro fue 49% como resultado de un alto porcentaje de autofecundación. La similitud de las estimaciones del grado de cruzamiento 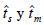 , indica que la autofecundación es la principal forma de endogamia en las poblaciones evaluadas. A pesar del elevado porcentaje de cruzamiento en las poblaciones del norte, el coeficiente de correlación de paternidad en ellas fue mayor a 60%, indicando un número reducido de árboles donadores de polen. Se discuten las implicaciones de estos resultados en la conservación de las poblaciones, sobre todo en la región centro.
, indica que la autofecundación es la principal forma de endogamia en las poblaciones evaluadas. A pesar del elevado porcentaje de cruzamiento en las poblaciones del norte, el coeficiente de correlación de paternidad en ellas fue mayor a 60%, indicando un número reducido de árboles donadores de polen. Se discuten las implicaciones de estos resultados en la conservación de las poblaciones, sobre todo en la región centro.
Palabras clave: Abeto Douglas, conservación genética, endogamia, fragmentación, polinización cruzada, tamaño de población.
Abstract
The Mexican populations of Pseudostuga menziesii (Mirb.) Franco have a restricted and fragmented natural distribution, especially in the central region, therefore there is a risk that these populations will be lost due to the effects of climatic change. With the purpose of suggesting adequate measures for their conservation, an evaluation was made of the mating pattern and its effect on the inbreeding level in three natural populations of P. menziesii, two of the north and one of the central region of México, based on the percentage of cross pollination. With the information of five polymorphic loci, the allelic frequencies of pollen and ovules were estimated, along with the proportion of mating in each population using multilocus and single locus models. No differences were found in the allelic frequencies of pollen and ovules, which indicates that the pollen comes from the same community as the mother trees. The value of in the two northern populations was close to 90%; in the central population it was 49% as a result of a high percentage of selfing. The similarity of the estimations of the degree of cross breeding 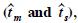 indicates that selfing is the principal form of inbreeding in the populations evaluated. Despite the high percentage of cross breeding in the northern populations, the paternity correlation coefficient in them was over 60%, indicating a reduced number of pollinating trees. The implications of these results in the conservation of the populations are discussed, especially in the central region.
indicates that selfing is the principal form of inbreeding in the populations evaluated. Despite the high percentage of cross breeding in the northern populations, the paternity correlation coefficient in them was over 60%, indicating a reduced number of pollinating trees. The implications of these results in the conservation of the populations are discussed, especially in the central region.
Key words: Douglas fir, genetic conservation, inbreeding, fragmentation, cross pollination, population size.
INTRODUCCIÓN
Pseudotsuga menziesii (Mirb.) Franco ha sido un componente importante de los ecosistemas forestales en Norteamérica desde el Pleistoceno (Hermann y Lavender, 1990). Diferentes registros palinológicos (Clisby y Sears, 1955; Lozano et al., 1993) indican que en el pasado remoto la distribución natural de ésta y otras especies de coníferas de origen boreal llegó hasta el sur de México. Sin embargo, el cambio climático asociado con el calentamiento posterior a las glaciaciones forzó a las coníferas a moverse nuevamente al norte o a mayores elevaciones en las montañas. En este proceso de contracción del área de distribución, algunas poblaciones de Pseudotsuga quedaron atrapadas, sin posibilidad de emigrar a otros sitios, lo que ocasionó una alteración importante de la distribución natural de la especie en su extremo sur, que ahora está fragmentada y discontinua, en forma de manchones o rodales pequeños aislados entre sí en el centro de México (Farjón, 1990).
El aislamiento genético, junto con otros procesos evolutivos como la selección natural, generalmente conduce a la diferenciación genética de las poblaciones (Savolainen y Kuittinen, 2000). En poblaciones mexicanas de P. menziesii (Mirb.) Franco, estudios con base en características morfológicas (Reyes et al., 2006) y fenológicas (Acevedo et al., 2006), indican una notoria diferenciación entre las poblaciones de la región norte y las del centro–sur del país, que representan el límite sur de la distribución natural del género Pseudotsuga en el mundo. El alto grado de diferenciación poblacional (Fst=0.298) ha sido confirmado con base en marcadores isoenzimáticos en un grupo de poblaciones mexicanas de esta especie, así como el reducido nivel de flujo genético entre ellas (Cruz, comunicación personal1).
Los efectos del aislamiento reproductivo son notorios en poblaciones pequeñas, ya que da lugar a un proceso de deriva genética (Falconer y Mackay, 1996), fenómeno que, además de aumentar la diferenciación entre las poblaciones, ocasiona un aumento en la homocigosis y la endogamia dentro de ellas (Savolainen y Kuittinen, 2000). Debido a que las especies de coníferas tienen un sistema reproductivo principalmente alógamo, el aumento de endogamia tiene consecuencias negativas sobre características de valor adaptativo de los individuos (Sorensen y Campbell, 1997). En P. menziesii, las poblaciones de la región central de México tienen una elevada producción de semillas vanas, y las semillas viables son de menor tamaño en comparación con las poblaciones del norte del país (Mápula–Larreta et al., 2007). Además, el germoplasma de la región norte tiene mayor vigor y capacidad de germinación, y las plántulas resultantes tienen mayor supervivencia y vigor que las de la región central (Juárez et al., 2006).
Los efectos característicos de la depresión endogámica observados en las poblaciones mexicanas de P. menziesii permiten suponer que el proceso de deriva genética está asociado con una elevada tasa de autofecundación, especialmente en las poblaciones del centro de México que son más pequeñas y tienen una menor densidad de arbolado adulto (Juárez et al., 2006; Mápula–Larreta et al., 2007). En forma similar a lo observado en Picea martinezii T. F. Pattersson (Ledig et al., 2000) y Picea mexicana Martínez (Ledig et al., 2002), especies de coníferas relictuales endémicas de México que tienen poblaciones pequeñas y aisladas donde se han encontrado altos niveles de autofecundación, en P. chihuahuana Martínez (Ledig et al., 1997) se encontró que el aumento de endogamia asociado a la autofecundación ocasiona una mayor producción de semillas vanas en las poblaciones.
A pesar de la distribución natural restringida y fragmentada que actualmente presenta P. menziesii en México, estas poblaciones tienen una elevada importancia ecológica y económica. Ecológicamente, P. menziesii es un elemento florístico clave en las comunidades naturales de los climas templado–fríos donde se encuentra; por el microambiente que genera, existe una gran diversidad de flora y fauna característica asociada con esta especie leñosa (Domínguez, 1994). Económicamente la especie tiene un alto valor maderable en al mundo (Hermann y Lavender, 1990), por lo que los recursos genéticos en estas poblaciones tienen una importancia estratégica en el proceso de domesticación de la especie. Sin embargo, los pronósticos de cambio climático (Iverson y Prasad, 2002) sugieren que los rodales ubicados en el extremo sur de su distribución natural estarán en riesgo de perderse. El riesgo es mayor si en las poblaciones existe una tasa reducida de cruzamiento, debido al efecto de la depresión endogámica con la consecuente reducción de la diversidad genética.
El presente estudio se realizó con el propósito de estimar el nivel de autofecundación en algunas poblaciones mexicanas de P. menziesii que permita sugerir medidas adecuadas para conservar estos recursos genéticos con base en las implicaciones derivadas del sistema de cruzamiento. Los objetivos específicos del trabajo fueron: a) determinar el patrón de cruzamiento en poblaciones mexicanas de P. menziesii y su efecto sobre el nivel de endogamia en ellas; b) comparar el porcentaje de polinización cruzada en poblaciones de diferente tamaño en el norte y centro de México.
MATERIALES Y MÉTODOS
Poblaciones muestreadas y análisis isoenzimático
Con el fin de comparar el patrón de cruzamiento se estudiaron tres poblaciones de P. menziesii, una del noroeste, una del noreste y una del centro de México (Cuadro 1). La población de Mohinora, en Chihuahua, es una comunidad forestal mezclada con Picea mexicana, Pinus arizonica Engelm., Populus tremuloides Michx. y Abies sp. El rodal donde se encuentra P. menziesii tiene una extensión aproximada de 69 ha, con una densidad promedio de 85 árboles adultos ha-1 de esta especie y más de 150 árboles adultos ha–1 de las otras especies. La población de Puerto Palomo, en Nuevo León, es un rodal puro y compacto de P. menziesii, con una extensión cercana a las 5 ha y una densidad promedio de 25 árboles adultos ha–1; a su alrededor hay otros rodales de la misma especie cercanos entre si. En la población de La Rosa, en Tlaxcala, P. menziesii está asociada con varias especies de Quercus, Abies religiosa (H. B. K.) Schl. et Cham., Pinus teocote Schl. et Cham. y P. rudis Endl. Este rodal tiene una superficie aproximada de 5 ha, a lo largo de una cañada de 2 km, con una densidad promedio de 35 árboles adultos ha–1 de la especie y más de 500 árboles ha–1 de las otras especies, con una dominancia notoria de encinos.
De cada población se usó una muestra variable de árboles madre dependiendo de la disponibilidad de semilla (Cuadro 1) y de cada árbol madre se usaron siete semillas. Se hizo la imbibición de la semilla en H2O2 al 1 % por 24 h y se estratificó a 4 °C por 50 d (Jarvis et al., 1997; Juárez et al., 2006). Las semillas germinaron en cajas Petri con papel filtro húmedo, a 24 °C en una cámara de germinación, hasta cuando la radícula alcanzó una longitud aproximada de 5 mm. Al momento de la extracción se separó el tejido del megagametofito (haploide) y del embrión (diploide) de cada semilla. Ambos tejidos se analizaron simultaneámente porque el megagametofito representa la contribución materna y, por tanto, tiene la misma constitución genética del óvulo que da origen al embrión de la semilla; esto permite deducir la contribución paterna (polen) por las diferencias con el genotipo del embrión. Este procedimiento permite conocer sin ambigüedad la contribución de los dos padres en el embrión (Mitton, 1983; Cheliak et al., 1985). Las muestras de cada tejido (megagametofito y embrión) fueron maceradas con un taladro eléctrico con punta de plástico, usando 100 µL de una solución de extracción (USDA Forest Service, 2003).
Los extractos obtenidos se almacenaron en un congelador a –81 °C hasta realizar el análisis. Los extractos fueron sometidos a electroforesis horizontal en geles de almidón, en el Laboratorio de Marcadores Genéticos del Colegio de Postgraduados, según la metodología descrita por el Laboratorio Nacional de Electroforesis y Genética Forestal del Servicio Forestal de los Estados Unidos (USDA Forest Service, 2003). Los sistemas enzimáticos usados fueron glutamato deshidrogenasa (GDH), fosfoglucosa isomerasa (PGI), fosfoglucomutasa (PGM) y glutamato–oxalacetato transaminasa (GOT), que en un estudio previo (Cruz, comunicación personal1) mostraron los mayores niveles de polimorfismo dentro de las poblaciones referidas. En algunos casos fue necesario ajustar los métodos de análisis para obtener un revelado adecuado de las variantes electroforéticas. Para las enzimas GOT y PGI se usó la metodología descrita por Stuber et al. (1988), ajustada y adecuada en el Laboratorio de Marcadores Genéticos.
La identificación de los alelos en cada locus se efectuó con base en la migración relativa de las enzimas a partir del punto de origen dentro de los geles. Para facilitar la interpretación de los loci y sus alelos correspondientes, en cada gel se incluyeron muestras del megagametofito de P. resinosa Ait., una especie completamente fija para estos loci. Cuando se identificaron varias zonas de actividad para una misma enzima, los loci se identificaron con numerales sucesivos a partir del cátodo. Para inferir la contribución paterna en cada embrión se analizó en forma paralela el megagametofito y el embrión de cada semilla. Al evaluar cada gel se determinó por diferencia la contribución paterna comparando las bandas electrofóreticas del megagametofito (genotipo materno) con las del embrión.
Estimación de los parámetros de cruzamiento y análisis estadístico
A partir de la contribución paterna y materna en los embriones se calcularon las frecuencias alélicas en el polen y los óvulos muestreados, para determinar si ambas representan el mismo tamaño de comunidad en la población (vecindario). La proporción de entrecruzamiento (t) se calculó con base en el modelo de genotipos de un locus génico y en el de genotipos de varios loci simultáneos de la progenie de cada árbol muestreado. Las estimaciones de  y se hicieron separadamente para cada población usando el método de máxima verosimilitud del programa MLTR (Ritland, 1994). Con el mismo programa se calculó la correlación de paternidad (rp) entre la progenie resultante del cruzamiento, que permite inferir sobre las estimaciones de y con base en el modelo de apareamiento mixto con autofecundación y polinización cruzada (Shaw y Allard, 1982). Este modelo supone que cada progenie viable (embrión) es el resultado de polinización cruzada aleatoria, con probabilidad "t", o de autofecundación, con probabilidad "s", de tal manera que s + t=1.
y se hicieron separadamente para cada población usando el método de máxima verosimilitud del programa MLTR (Ritland, 1994). Con el mismo programa se calculó la correlación de paternidad (rp) entre la progenie resultante del cruzamiento, que permite inferir sobre las estimaciones de y con base en el modelo de apareamiento mixto con autofecundación y polinización cruzada (Shaw y Allard, 1982). Este modelo supone que cada progenie viable (embrión) es el resultado de polinización cruzada aleatoria, con probabilidad "t", o de autofecundación, con probabilidad "s", de tal manera que s + t=1.
En el caso de la estimación de se supone que existe independencia entre los loci utilizados para estimar la proporción de entrecruzamiento (Furnier y Adams, 1986). Si esta condición no se cumple, el valor de estará sesgado; este sesgo es relativamente pequeño a menos que los loci involucrados estén fuertemente ligados y exista un desequilibrio de ligamento elevado en la población (Shaw et al., 1981). A pesar del posible sesgo, desde el punto de vista estadístico la estimación multilocus es más eficiente que la estimación basada en un sólo locus ya que la primera toma en cuenta varios loci simultáneamente. Además, mientras mayor sea el número de loci incluidos para estimar es posible detectar más eventos de polinización cruzada, por lo cual este estimador es menos sensible a las violaciones de los supuestos del modelo de apareamiento mixto (Shaw et al., 1981). El valor de puede variar en gran medida de un locus a otro, generando mayor heterogeneidad (error estándar) al estimarlo en diferentes poblaciones (Shaw y Allard, 1979).
Es importante estimar ambos valores de entrecruzamiento ya que cuando es menor que , se infiere que además de la autofecundación en la población se encuentran otras formas de endogamia, como el apareamiento entre parientes (Burczyk et al., 1997). En este caso, los valores de se estimaron para cada población usando el algoritmo EM para calcular las frecuencias alélicas de óvulos y polen (Ritland, 1994). Se usaron 200 muestras con reemplazo (bootstraps) para estimar el error estándar en cada uno de ellos. Para determinar si los valores de cruzamiento difieren entre las poblaciones se calcularon sus respectivos intervalos de confianza, con p<0.05.
RESULTADOS Y DISCUSIÓN
En ninguna población hubo diferencias significativas (p>0.05) en las frecuencias alélicas de los dos gametos (Figura 1). Esto sugiere que el acervo genético contenido en el polen representa las frecuencias alélicas locales de los árboles maternos y, por tanto, ambos (polen y óvulos) representan el mismo tamaño de vecindario dentro de cada población. Esto no es común en especies de polinización anemófila, donde generalmente el polen puede viajar grandes distancias y representar a una comunidad (vecindario) de mayor tamaño que la muestra de individuos maternos (Ledig, 1998). Por ejemplo, en Pinus greggii Engelm. se ha observado que el acervo de polen generalmente representa una comunidad (vecindario) de mayor tamaño que el acervo de óvulos dentro de la población (Parraguirre et al., 2004). Sin embargo, dado que los rodales de P. menziesii estudiados son mucho más pequeños, es posible que ambas muestras (polen y óvulos) representan a toda la población. Aún en la población de Mohinora, más extensa que las otras dos, no se encontraron diferencias entre las frecuencias génicas de ambos gametos.
Sistema de cruzamiento
En ninguna población hubo diferencias significativas (p>0.05) entre los valores promedio de polinización cruzada obtenidos con la información de locus múltiples o de locus simple (Cuadro 2). En la mayoría de los estudios de este tipo es común que el valor de sea ligeramente mayor al de ya que el primer estimador detecta más eficientemente la presencia de polinización cruzada (Shaw y Allard, 1982); sin embargo, en este caso, en ninguna de las poblaciones las diferencias numéricas entre los dos estimadores fueron significativas, por lo que ambos procedimientos son consistentes.
Las dos poblaciones del norte de México presentaron un porcentaje de polinización cruzada cercano o ligeramente superior a 90% (Cuadro 2) y sin diferencias significativas (p>0.05) entre ambos tipos de estimación. Estos valores están dentro del intervalo estimado para otras poblaciones naturales de esta especie en Norteamérica (Shaw y Allard, 1982; Neale y Adams, 1985; Yeh y Morgan, 1987), o incluso en plantaciones y huertos semilleros (Ritland y El–Kassaby, 1985). Este grado de polinización cruzada es común en la mayoría de las especies de coníferas en poblaciones relativamente grandes y refleja la capacidad de dispersión del polen a grandes distancias. Ledig (1998) señala que en especies con polinización anemófila, como en el género Pinus y otras coníferas, el porcentaje de polinización cruzada es cercano a 1.0 cuando no existen otros factores que interfieran en el fenómeno.
En contraste, el valor estimado de polinización cruzada en la población de La Rosa, Tlaxcala, casi 50%, es menor con respecto al de ambas poblaciones del norte del país y es muy inferior al encontrado para esta especie en su hábitat natural. En poblaciones naturales de otras especies de coníferas mexicanas que comparten similitudes en tamaño y aislamiento geográfico, también se han encontrado valores reducidos de polinización cruzada; por ejemplo, 0.399 y 0.589 para dos poblaciones de Picea martinezii (Ledig et al., 2000). En P. mexicana, el valor promedio en tres poblaciones varió de 0.590 a 0.807 (Ledig et al., 2002) y en algunas poblaciones de P. chihuahuana el valor fue incluso cercano a cero (Ledig et al., 1997).
Aunque en la población de P. menziesii de La Rosa no se encontró un valor tan bajo de polinización cruzada como en P. chihuahuana, sí refleja la presencia de un nivel importante de endogamia que podría estar asociado a la frecuencia de autofecundación en sus eventos reproductivos. En especies con un sistema reproductivo que favorece la polinización cruzada, la autofecundación generalmente ocasiona depresión endogámica en la progenie, que se refleja en la reducción en la producción de semilla viable, así como en el vigor y tasa de crecimiento de las plántulas (Frankham, 1998; Mápula–Larreta et al., 2007). Un fenómeno similar es posible que ocurra en las poblaciones naturales de P. menziesii del centro de México, según Juárez et al. (2006) y Mápula–Larreta et al. (2007), quienes documentaron una alta proporción de semillas vanas y una menor capacidad y vigor germinativo de las semillas llenas de esta región en relación con semillas de las poblaciones del norte de México.
Con base en el porcentaje de autofecundación estimado en la población La Rosa se esperaría encontrar un alto grado de endogamia y una deficiencia notoria de heterocigotos en su progenie; sin embargo, los datos de diversidad genética estimados con base en una muestra de 18 loci izoenzimáticos no corroboran esta situación (Cruz, comunicación personal1). Una posible explicación a este fenómeno es que en esta población hay una fuerte presión de selección durante las etapas tempranas de desarrollo del cigoto en contra de los embriones homocigóticos producto de la autofecundación, causando una gran cantidad de semillas vanas (Sorensen, 1982). En Picea glauca (Moench) Voss., Sproule y Dancik (1996) señalan un fenómeno similar donde, a pesar de existir un elevado porcentaje de autofecundación y endogamia, no se encontró una deficiencia de individuos heterocigotos en la progenie, lo que se atribuye a una fuerte presión de selección sobre los embriones derivados por autofecundación, asociada a la presencia de genes recesivos letales.
Correlación de paternidad
A pesar de que en las poblaciones del norte de México se encontró un bajo porcentaje de autofecundación, la correlación de paternidad entre las progenies de estas poblaciones fue relativamente elevada (rp > 0.60), lo que indica una alta probabilidad de que dos individuos del mismo árbol madre compartan un mismo padre (Cuadro 2). Una alta correlación de paternidad y una elevada tasa de polinización cruzada implica que un número reducido de padres participaron en la polinización de cada uno de los árboles madre muestreados, ya sea por la interferencia en la dispersión del polen causada por la presencia de otras especies, por el reducido número de individuos reproductivos existentes, o por problemas de sincronización fenológica en la dispersión del polen. Dado que Puerto Palomo, N. L., es un rodal puro de P. menziesii pero con una baja densidad de arbolado adulto, es posible que los problemas de sincronización fenológica, junto con el reducido número de individuos, hayan contribuido a una elevada correlación de paternidad. La participación de un número reducido de individuos en los eventos reproductivos puede conducir a una disminución de la diversidad genética en las poblaciones, lo cual pone en riesgo la persistencia de la población si no hay intercambio genético entre los individuos que la constituyen.
Las tasas de cruzamiento y de correlación de paternidad indican que las tres poblaciones muestreadas están expuestas a la reducción de la diversidad genética. Sin embargo, los esfuerzos de conservación de esta especie se deben enfocar a las poblaciones de la Región Central, como la de La Rosa, por su bajo porcentaje de polinización cruzada. En estas poblaciones es importante restaurar el intercambio genético y promover la participación de un mayor número de individuos en los eventos reproductivos. Debido a los efectos de depresión endogámica observados en estas poblaciones, se sugiere propiciar el intercambio genético (movimiento de polen) entre las poblaciones del centro. Así, la introducción de plantas producidas en vivero de otras poblaciones del centro puede ser útil para aumentar la diversidad genética y promover la polinización cruzada entre los individuos, ya que la introducción de germoplasma del norte en las poblaciones naturales del centro del país, como un mecanismo para aumentar la diversidad genética y reducir los efectos de la endogamia, no es recomendable debido a la notoria diferencia en las características adaptativas de las poblaciones del centro y del norte de México (Acevedo et al., 2006).
CONCLUSIONES
Las frecuencias alélicas del polen y de los óvulos fueron similares en las tres poblaciones estudiadas, lo que implica que las frecuencias alélicas del polen representan a las frecuencias alélicas de los árboles madre. Por tanto, no hay evidencias de participación de polen extraño a la comunidad muestreada en cada población. El porcentaje de polinización cruzada fue cercano al máximo posible en las poblaciones del norte, acorde a lo esperado para especies de coníferas con polinización anemófila. Sin embargo, en la población del centro de México (La Rosa) se encontró una reducción importante en la polinización cruzada, que puede asociarse con el aislamiento geográfico y el tamaño reducido de la población. No se encontraron diferencias significativas entre las estimaciones con locus múltiples y locus simples de la polinización cruzada, resultando que la principal forma de endogamia observada en la población de La Rosa es la autofecundación. La alta correlación de paternidad en las poblaciones del norte sugiere que aunque ocurre polinización cruzada, hay una elevada proporción de hermanos completos que resultan de una baja participación de individuos reproductivos en la población.
Para reducir los efectos negativos de la endogamia acumulada en las poblaciones de P. menziesii del centro de México, se sugiere mezclar polen de estas poblaciones y polinizar árboles en la región, para generar variación genética adicional.
AGRADECIMIENTOS
Esta investigación se realizó con apoyo de los proyectos "Diversidad Genética y Conservación de Pseudotsuga en México", financiado por el CONACYT (Proyecto 33617–B), y "Conservación y Mejoramiento Genético de Pseudotsuga spp., conífera estratégica del estado de Tlaxcala y la región central del país" financiado por el Fondo Sectorial CONAFOR–CONACYT (Proyecto 2002–C01–6416). Se agradece de manera especial al Laboratorio de Marcadores Genéticos del CP por las facilidades brindadas y en particular al C. Juan Carlos Zaragoza Ramírez por su valiosa colaboración en los análisis de laboratorio.
LITERATURA CITADA
Acevedo R., R., J. J. Vargas H., J. López U., y J. Velázquez M. 2006. Efecto de la procedencia geográfica y de la fertilización en la fenología del brote terminal en plántulas de Pseudotsuga sp. Agrociencia 40: 125–137. [ Links ]
Burczyk, J., W. T. Adams, and J. Y. Shimizu. 1997. Mating system and genetic diversity in natural populations of Knobcone pine (Pinus attenuata). For. Gen. 4(4): 223–226. [ Links ]
Cheliak, W. M., J. A. Pitel, and G. Murray. 1985. Population structure and the mating system of white spruce. Can. J. For. Res. 15: 301–308. [ Links ]
Clisby K., H., and P. B. Sears. 1955. Palynology in southern North America. Part III: Microfossil profiles under Mexico City correlated with the sedimentary profiles. Bull. Geol. Soc. Am. 66:511–520. [ Links ]
Domínguez A., F.A. 1994. Análisis histórico–ecológico de los bosques de Pseudotsuga en México. INIFAP–CIR Golfo Centro. Folleto Técnico No. 23. México, 43 p. [ Links ]
Falconer, D. S., y T. F. C. Mackay. 1996. Introducción a la Genética Cuantitativa. Caballero R., A., C. López–Fanjul, M. A. Toro I. y A. Blasco M. (trads). ACRIBIA S. A., Zaragoza, España. 469 p. [ Links ]
Farjon, A. 1990. Pinaceae: Drawings and Descriptions of the Genera Abies, Cedrus, Pseudolarix, Keteeleria, Nothotsuga, Tsuga, Cathaya, Pseudotsuga, Larix and Picea. Koeltz Scientific Books. Konigstein, Federal Republic of Germany. pp: 171–191. [ Links ]
Frankham, R. 1998. Inbreeding and extinction island populations. Conservation Biol. 12: 665–675. [ Links ]
Furnier, G. R., and W. T. Adams. 1986. Mating system in natural populations of Jeffrey pine. Am. J. Bot. 73(7): 1002–1008. [ Links ]
Hermann, R. K., and D. P. Lavender. 1990. Pseudotsuga menziesii (Mirb.) Franco. In: Burns, R. and B. H. Honkala (eds). Silvics of North America, Vol. 1. Conifers. USDA Forest Service. Washington, D. C. pp: 540–557. [ Links ]
Iverson, L. R., and A. M. Prasad. 2002. Potential redistribution of tree species habitat under five climate change scenarios in the easthern U. S. For. Ecol. Manag. 155:205–222. [ Links ]
Jarvis, S. B., M.A. Taylor, J. Bianco, F. Corbineau, and H.V. Davies. 1997. Dormancy–breakage in seeds of Douglas fir (Pseudotsuga menziesii (Mirb.) Franco) support for the hypothesis that LEA gene expression is essential for this process. J. Plant Physiol. 151: 457–464. [ Links ]
Juárez A., A., J. López U., J.J. Vargas H., y C. Sáenz R. 2006. Variación geográfica en la germinación y crecimiento inicial de plántulas de Pseudotsuga menziesii de México. Agrociencia 40: 783–792. [ Links ]
Ledig, F.T. 1998. Genetic variation in Pinus. In: Richardson, D.M. (ed). Ecology and Biogeography of Pinus. Cambridge University Press, Cambridge. pp: 251–280. [ Links ]
Ledig, F. T., B. Bermejo V., P. D. Hodgkiss, D. R. Johnson, C. Flores L., and V. Jacob C. 2000. The mating system and genetic diversity in Martínez spruce, an extremely rare endemic of México's Sierra Madre Oriental: an example of facultative selfing and survival in interglacial refugia. Can. J. For. Res. 30:1–9. [ Links ]
Ledig, F. T., P. D. Hodgkiss, and V. Jacob C. 2002. Genetic diversity, mating system, and conservation of a Mexican subalpine relict, Picea mexicana Martínez. Conservation Genetics 3:113–122. [ Links ]
Ledig, F. T., V. Jacob C., P. D. Hodgkiss, and T. Eguiluz P. 1997. Evolution and divergence among populations of a rare Mexican endemic, Chihuahua spruce, following Holocene warming. Evolution 51: 1815– 1827. [ Links ]
Lozano G., M., S. B. Ortega G., M. Caballero M., and J. Urrutia F. 1993. Late Pleistocene and Holocene paleoenviroments of Chalco Lake, central Mexico. Quat. Res. 40:332–342. [ Links ]
Mápula–Larreta, M., J. López–Upton, J. J. Vargas–Hernández, and A. Hernández–Livera. 2007. Reproductive indicators in natural populations of Douglas–fir in Mexico. Biodiversity and Conservation 16: 727–742. [ Links ]
Mitton, J. B. 1983. Conifers, In: Tanksley, S. D., and T. J. Orton (eds). Isozymes in Plant Genetic and Plant Breeding, part B. Elsevier, Amsterdam. pp: 443–472. [ Links ]
Neale, D. B., and W. T. Adams. 1985. The mating system in natural shelterwood stands of Douglas–fir. Theor. Appl. Genet. 71:201–207. [ Links ]
Parraguirre, L., C., J. J. Vargas H., P. Ramírez V., y C. Ramírez H. 2004. Sistema de cruzamiento en cuatro poblaciones naturales de Pinus greggii Engelm. Agrociencia 38: 107–119. [ Links ]
Reyes H., V. J., J. J. Vargas H., J. López U., y H. Vaquera H. 2006. Similitud fenotípica de poblaciones mexicanas de Pseudotsuga Carr. Agrociencia 40: 545–556. [ Links ]
Ritland, K. 1994. Multilocus mating system program MLTR, version 0.9 K. Ritland. Departament of Botany, University of Toronto, Toronto, Ontario, Canada. [ Links ]
Ritland, K., and Y. A. El–Kassaby. 1985. The nature of inbreeding in a seed orchard of Douglas fir as shown by an efficient multilocus model. Theor. Appl. Genet. 71:375–384 [ Links ]
Savolainen, 0., and H. Kuittinen. 2000. Small population processes. In: Young, A., D. Boshier, and T. Boyle (eds). Forest Conservation Genetics: Principles and Practice. CSIRO Publishing, Australia. pp: 91–100. [ Links ]
Shaw, D. V., and R. W. Allard. 1979. Analysis of mating system parameters and population structure in Douglas–fir using single–locus and multilocus methods. In: Proc. Symp. Isozymes of North American Forest Trees and Forest Insects. Berkeley, Cal. USDA For. Serv., Gen. Tech. Rep. PSW–48. Berkeley, Cal. pp: 18–22. [ Links ]
Shaw, D. V., and R. W. Allard. 1982. Estimation of outcrossing rates in Douglas–fir using isozymes markers. Theor. Appl. Genet. 62: 113–120. [ Links ]
Shaw, D. V., A. L. Kahler, and R. W. Allard. 1981. A multilocus estimator of mating parameters in plant populations. Proc. Nat. Acad. Sci. 78(2): 1298–1302. [ Links ]
Sorensen, F.C. 1982. The role of polyembriony and embryo viability in the genetic system of conifers. Evolution 36: 725–733. [ Links ]
Sorensen, F. C., and R. K. Campbell. 1997. Near neighbor pollination and plant vigor in coastal Douglas–fir. For. Gen. 4: 149–157. [ Links ]
Sproule, A. T., and B. P. Dancik. 1996 .The mating system of black spruce in North–Central Alberta, Canada. Silvae Genet. 45: 159–164. [ Links ]
Stuber, C.W., J.F. Wendel, M.M. Goodman, and J. S. C. Smith. 1988. Techniques and scoring procedures for starch gel electrophoresis of enzymes from maize (Zea mays L.). North Carolina Agricultural Research Service. North Carolina State University. Raleigh, N.C., U.S.A. Technical Bulletin. No. 286. 87 p. [ Links ]
USDA (United States Departament of Agriculture) Forest Service. 2003. The National Forest Genetics Laboratory (NFGEL) Standard Operating Procedures. Pacific Southwest Research Station. Placerville, CA. 90 p. [ Links ]
Yeh, F. C., and K. Morgan. 1987. Mating system and multilocus associations in a natural population of Pseudotsuga (Mirb.) Franco. Theor. Appl. Genet. 73: 799–808. [ Links ]
1 Cruz N., J. 2007. Diversidad genética y patrón de cruzamiento en poblaciones naturales de Pseudotsuga menziesii (Mirb.) Franco en México. Tesis de Maestría en Ciencias. Colegio de Postgraduados, Campus Montecillo. Texcoco, Méx. 60 p. [ Links ]regresar

