










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgrociencia
versión On-line ISSN 2521-9766versión impresa ISSN 1405-3195
Agrociencia vol.42 no.3 Texcoco abr./may. 2008
Recursos naturales renovables
Variation in quantitative vessel element features of Juglans regia wood in the western black sea region of Turkey
Variación en características cuantitativas de elementos de vaso en madera de Juglans regia en la región occidental del Mar Negro, Turquía
Barbaros Yaman
Zonguldak Karaelmas University, Bartin Faculty of Forestry, Department of Forest Botany, Bartin/ Turkey (yamanbar2000@yahoo.com)
Recibido: Marzo, 2007.
Aprobado: Febrero, 2008.
Abstract
Variations in quantitative vessel element characters of Juglans regia L. wood were studied in relation to altitude and growth–ring width. Multiple regression analysis using altitude and growth–ring width as independent variables were performed. Significant correlations were obtained for tangential vessel and radial vessel diameter (p< 0.001), vulnerability ratio and xeromorphy ratio (p<0.01), and for the number of vessels per group (p<0.05). Vessel frequency is not correlated with independent variables. Non–anatomical factors (altitude and growth–ring width) explain 40.4–80.4% of variation for dependent variables.
Key words: Juglans regia, altitude, ecology, growth–rings, wood anatomy.
Resumen
Se estudiaron variaciones en características cuantitativas de los elementos de vaso en la madera de Juglans regia L., en relación con la altitud y el ancho de los anillos de crecimiento. Se hicieron análisis de regresión, utilizando la altitud y el ancho de los anillos de crecimiento como variables independientes. Se obtuvieron correlaciones significativas para los diámetros de vaso tangencial y radial (p< 0.001), las tasas de vulnerabilidad y xeromorfia (p<0.01), y para el número de vasos por grupo (p<0.05). La frecuencia de vaso no se correlaciona con las variables independientes. Factores no anatómicos (altitud y ancho del anillo de crecimiento) explican 40.4–80.4% de la variación en las variables dependientes.
Palabras clave: Juglans regia, altitud, ecología, anillos de crecimiento, anatomía de la madera.
INTRODUCTION
Anatomical features in wood are affected by environmental factors such as temperature, light intensity, water, nutrients (soil fertility), gravity, photoperiod, climate periodicity, wind and frost (Wodzicki, 2001). The basic knowledge on ecological wood anatomy comes from Carlquist (1966 and 1977a,b), Baas (1973), Graaff and Baas (1974); besides, several studies have been carried out on this subject (Alves and Angyalossy–Alfonso, 2000; Lens et al., 2003; Rodríguez et al., 2006). Most of the studies reveal variations in wood anatomical properties, especially vessel elements, in response to different ecological conditions. Vessel element properties are affected by climatic factors such as temperature, water availability, etc., in connection with latitude, altitude, and different habitats (Baas, 1982; Graaff and Baas, 1974; Carlquist, 1988). Carlquist (1977b) and Carlquist and DeBuhr (1977) introduced two equations relating to ecological wood anatomy: vulnerability (mean vessel diameter divided by mean vessel density) and mesomorphy (vulnerability multiplied by vessel element length).
Anatomical variations of Turkish maples (Yaltirik, 1970) and beech woods (Sanli, 1977) were examined with relation to habitat and altitude. Wood structures of Ostrya carpinifolia (Gercek et al., 1998), Rhododendron sp. (Merev and Yavuz, 2000), Populus tremula (Yaman and Saribas, 2004) and Cerasus avium (Yaman, 2006) were studied with regard to ecological perspective.
Paleobotanical and archaeological investigations in Turkey show that J. regia has been in Anatolia for many thousands of years (Aytug and Sanli, 1974; Sanli, 1989; Aytug and Gorcelioglu, 1987). It is presumed native in the north–eastern and eastern Anatolia and it has been widely planted and naturalized throughout Turkey (Yaltirik, 1982; Leslie and McGranahan, 1988). Due to its valuable wood the trunks of J. regia were exported from Anatolia to European countries until 1950 (Yaltirik, 1982). In terms of both fruit and wood, J. regia is one of the most economically important taxa in the world and Turkey (Voulgaridis and Vassiliou, 2006). Its wood anatomical range and physical and mechanical properties have been studied by Verani (1993), Merev (1998) and Kantay et al. (2000).
Of both individuals in humid sites and cold and drought tolerant genotypes in Anatolia having extreme differences in geographic and climatic conditions (Sykes, 1975), there is an opportunity for studying wood anatomical variation in J. regia wood. Therefore, the aim of the present study was to study variations in quantitative vessel element features of J. regia wood in relation to altitude and growth–ring width in the western Black Sea region of Turkey.
MATERIALS Y METHODS
The 12 sample trees were selected from different altitudes but with the same exposition (northern) in the western Black Sea region of Turkey. Trunk diameters at breast height and locality data of sample trees are shown in Table 1.
The Black Sea region is a macroclimate type with mild temperatures and precipitation almost uniformly distributed throughout the year. In some degree, its sub–macroclimate type in western Black Sea region has lower average values of both temperature and precipitation than that of eastern Black Sea region (Erinc, 1996).
The wood specimens (increment cores with both 12 mm and 5 mm in diameter) were extracted from the same side (west) at breast height of the trunks using two different increment borers. The former was used for anatomical investigation, and the latter for growth–ring measurements.
In the laboratory the samples were boiled and placed in a solution with equal parts of water, glycerine and ethyl alcohol for softening. Transverse, radial, and tangential sections (15 µm thick), including growth–rings of 2002 and 2003, were taken from them using an Euromex sliding microtome. Preparation of the sections was done according Yaltirik (1971). The following anatomical variables were determined: tangential and radial lumen diameter of vessel, vessel frequency, and the number of vessels per group; vulnerability ratio was calculated for every sample tree; 25 measurements or counting were performed for each variable (1200 measurements for all samples). Additionally, for every sample tree, growth–ring widths from 1984 to 2003 were measured using VIAS Time–Table.
Xeromorphy ratio (XERO) was developed for practical studies of ecological wood anatomy: the inner wall–surface in contact with water column/volume in vessel element is multiplied by vessel frequency; if it is assumed that a vessel element is approximately elliptic cylinder, the lateral wall–surface (S) to volume (V) ratio is:
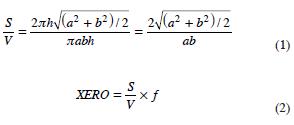
where, a = major radius of ellipse; b = minor radius of ellipse (a = half radial vessel diameter; b = half tangential vessel diameter); h = length of elliptic cylinder (vessel element length); f = vessel frequency.
Data on altitude, growth–ring width, selected anatomical characters, vulnerability ratio and xeromorphy ratio were analysed by multiple regression using the SPSS 9.0 program. In the analysis, altitude and growth–ring width were considered as independent variables, and wood anatomical characters selected, vulnerability ratio and xeromorhy ratio were used as dependent variables.
RESULTS AND DISCUSSION
J. regia has a semi–ring porous wood. Growth–ring boundaries are distinct because of differences in vessel diameter between early and late–wood in successive rings. Vessels are mostly solitary, or partly in radial multiples of 2–5. Solitary vessels are oval to round in outline. Vessel elements have simple perforation plates with oblique end walls. Inter–vessel pits, with slit–like apertures, are alternate and their shape range from polygonal to round. Tangential and radial vessel diameter on average are 135.3 µm (114–152.8 µm) and 177.1 µm (152–214.8 µm). Vessel frequency ranges from 7.8 to 11.8, (x 9.1), whereas the number of vessels per group ranges from 1.16 to 1.68 (1.39). Vulnerability ratio and xeromorphy ratio of J. regia range from 9.7 to 18.6 and from 0.18 to 0.36. Vulnerability ratio, xeromorphy ratio, and quantitative data belonging to anatomical properties selected were tabulated separately for each sample tree (Table 2).
9.1), whereas the number of vessels per group ranges from 1.16 to 1.68 (1.39). Vulnerability ratio and xeromorphy ratio of J. regia range from 9.7 to 18.6 and from 0.18 to 0.36. Vulnerability ratio, xeromorphy ratio, and quantitative data belonging to anatomical properties selected were tabulated separately for each sample tree (Table 2).
Altitude and growth–ring width as independent variables showed significant R2 (0.404 to 0.804) for tangential and radial vessel diameter (p<0.001), vulnerability ratio and xeromorphy ratio (p<0.01), and for the number of vessels per group (p<0.05). Thus, non–anatomical factors (ALT and GRW) explain 40.4–80.4% of total variation for those dependent variables. For ALT, standardized regression coefficients were significant for tangential and radial vessel diameter (p<0.001), vulnerability ratio and xeromorphy ratio (p<0.01), and for the number of vessels per group (p<0.05), but for growth–ring width only radial vessel diameter was significant (p<0.05). The largest standardized R2 between dependent variables and non–anatomical factors were –0.917 for tangential vessel diameter, –0.717 for radial vessel diameter, –0.760 for vulnerability ratio, and 0.721 for xeromorphy ratio (Table 3). Trends in the values of these dependent variables in respect of independent variables are plotted in Figures 1–4 (Figures 1, 2, 3 and 4).
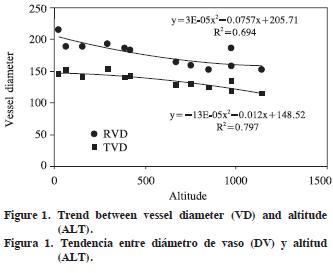

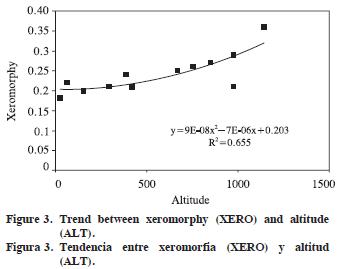
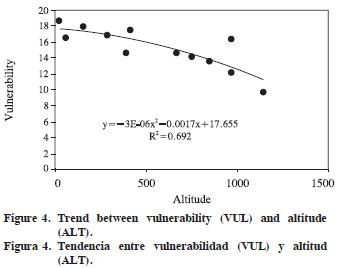
Vessel frequency and tangential vessel diameter of J. regia wood in Israel and adjacent regions are 2–20 mm–2 and 40–150 µm (Fahn et al., 1986). The values of Himalayan J. regia wood are 5–8 mm–2 and 50–250 µm (Suzuki et al., 1991). As for Turkish J. regia (Merev, 1998), tangential and radial vessel diameters on average are 136.13 µm and 161.80 µm in early–wood, 84.55 µm and 75.26 µm in late–wood. Mean vessel frequency and the number of vessels per group are 12.82 mm–2 and 1.36. In the present study, tangential vessel diameter, vessel frequency, and the number of vessels per group are 135.3 µm, 9.1 mm–2, and 1.39 on average.
The present results point out the impacts of altitudinal gradients on tangential and radial vessel diameters, the number of vessels per group, vulnerability ratio and xeromorphy ratio of J. regia wood in western Black Sea region. With increasing altitude in the field, while vulnerability ratio and both tangential and radial diameter of vessel elements tend to decrease, xeromorphy ratio and the number of vessels per group tend to increase. However, vessel frequency is not correlated with altitude. The close relations between vessel element properties and altitude also appear in other anatomical studies: in intraspecific level while vessel frequency is positively correlated with altitude (Noshiro and Suzuki, 1995; Yaman and Saribas, 2004; Yaman, 2006), there are negative correlations between altitude and vessel diameter (Merev and Yavuz, 2000; Yaman and Saribas, 2004; Yaman 2006), vessel element length (Noshiro et al., 1994; Gercek et al., 1998; Yaman, 2006), solitary vessel ratio (Noshiro et al., 1994). However, in Rhododendron campanulatum wood, contrary to aforementioned relations, vessel element length and vessel frequency are positively and negatively correlated with altitude (Noshiro and Suzuki, 1995). Xinying et al. (1988) also report that the negatively correlation appears between altitude and vessel frequency, and that the positively correlation occurs between altitude and vessel diameter, and vessel element length. Moreover, Graaff and Baas (1974) point out that there are not correlations between above mentioned features of vessel elements and altitude in the species of Pittosporum, Calophyllum, and Schuurmansia with a wide altitudinal range. In the absence of altitudinal trends in Dodonaea viscosa wood, a species with a worldwide distribution, Liu and Noshiro (2003) suggest that the species–level variation in wood anatomy is not controlled by ecological gradients in some species.
Carlquist (1988) points out that the correlations between some quantitative vessel element features and altitude should be related to water availability and temperature factors. Extreme low temperatures affect the viscosity of water both in soil and in stem, and are a major factor limiting the taking up of water from soil and the transport of water in the secondary xylem (Cepel, 1993; Cavender–Bares, 2005). According to Roderick and Berry (2001), the effect of temperature on the viscosity of water will affect vessel size or number and, therefore, wood density. The physiological link between water viscosity and wood anatomy in Eucalyptus camaldulensis has been reported by Thomas et al. (2004). Very narrow vessel diameter–low vulnerability values (<1)–denote high conductive safety, and very wider vessel diameter–high vulnerability values (> 3)–are an indicator of high conductive efficiency (Carlquist, 1977b). The mean vulnerability ratio (15.2) shows that J. regia wood in the field has high conductive efficiency, but it is vulnerable to embolism. However, in comparison with those in lower altitudes, narrower vessel diameter, lower vulnerability ratio, and more vessels grouping in J. regia wood in higher altitude might be explained as enhancement of conductive safety in some degree.
The extremely low xeromorphy ratios of J. regia denote mesomorphic wood structure for this species. In spite of non–significant relationship between vessel frequency and altitude, the increasing of xeromorphy ratio from low–to high–altitudes is due to increasing of wall–surface area to volume ratio together with narrowing of vessel element diameter along altitudinal gradients. The variation in vessel elements along altitudinal gradient might be a response to decreasing in temperatures along the gradients in the region.
In our study, except for radial vessel diameter, significant correlations do not occur between growth–ring widths and other anatomical properties examined. However, Yaman (2006) found significant correlations between growth–ring widths and vessel frequency, vulnerability ratio, and mesomorphy ratio in Cerasus avium wood.
In the base of data of Sanli (1977), Carlquist and Hoekman (1985) and Merev (1998) it might be proposed as a pre–determination that xeromorphy ratios above 9 denote xeromorphic–, and those under 9 indicate mesomorphic– wood structure. In some woody plants, in case of the probable xeromorphy ratios contrast to the ecological sites represented, another ways of ecological adaptation might be considered (Carlquist, 1988).
CONCLUSIONS
Like in most of other species examined in relation to ecological wood anatomy, some quantitative features of vessel elements in J. regia wood also correlate with altitude. Xeromorphy ratio, introduced in the present study, appears to be related to ecological factors. This ratio, as well as Carlquist's vulnerability and mesomorphy index, might be descriptive of xeromorphic or mesomorphic species in respect of ecological wood anatomy. However, to determine whether it might be used in ecological, paleobotanical, and archeological investigations of wood, further field studies should be carried out.
LITERATURE CITED
Alves, E. S., and V. Angyalossy–Alfonso. 2000. Ecological trends in the wood anatomy of some Brazilian species 1. Growth rings and vessels. IAWA J. 21: 3–30. [ Links ]
Aytug, B., and I. Sanli. 1974. Forets de la fin du tertiaire aux environs du Bosphore. Revue De La Faculté Forestière De L'Université D'Istanbul 24: 64–78. [ Links ]
Aytug, B., and E. Gorcelioglu. 1987. Wood and wooden furniture found in the royal tomb at Gordion. Review of the Faculty of Forestry, University of Istanbul 37: 1–27. [ Links ]
Baas, P. 1982. Systematic, phylogenetic and ecological wood anatomy–history and perspectives. In: Baas, P. (ed.). New Perspectives in Wood Anatomy. Martinus Nijhoff Pub. Boston. pp: 23–58. [ Links ]
Carlquist, S. 1977a. Ecological factors in wood evolution: a floristic approach. Am. J. Bot. 64: 887–896. [ Links ]
Carlquist, S. 1977b. Wood anatomy of Onagraceae: additional species and concepts. Annals of the Missouri Botanical Garden 64: 627–637. [ Links ]
Carlquist, S. 1988. Comparative Wood Anatomy. Springer– Verlag. Berlin. 358 p. [ Links ]
Carlquist, S., and L. DeBuhr. 1977. Wood anatomy of Penaeaceae (Myrtales): comparative phylogenetic and ecological implications. Bot. J. of the Linnean Soc. 75: 211–227. [ Links ]
Carlquist, S., and D. A. Hoekman. 1985. Ecological wood anatomy of the woody southern California flora. IAWA Bulletin n.s. 6: 319–347. [ Links ]
Cavender–Bares, J. 2005. Impacts of freezing on long–distance transport in woody plants. In: Holbrook, N. M., M. Zwieniecki and P. Melcher (eds). Vascular Transport in Plants. Elsevier Inc. Oxford. pp: 401–424. [ Links ]
Cepel, N. 1993. Soil–Water–Plant Relationships. Istanbul Univ. Press. Istanbul. 236 p. [ Links ]
Erinc, S. 1996. Climatology and Methods. Alfa Press. Istanbul. 538 p. [ Links ]
Fahn, A., E. Werker, and P. Baas. 1986. Wood Anatomy and Identification of Trees and Shrubs from Israel and Adjacent Regions. The Israel Academy of Sciences and Humanities Jerusalem. 221 p. [ Links ]
Gercek, Z., N. Merev, R. Ansin, Z. C. Ozkan, S. Terzioglu, B. Serdar, and T. Birturk. 1998. Ecological wood anatomy of Ostrya carpinifolia Scop. in Turkey. In: Elicin, G. (ed). Symposium on Quercus vulcanica and Flora of Turkey. Cantay Pub. Istanbul. pp: 302–316. [ Links ]
Graaff, N. A.v.d., and P. Baas. 1974. Wood anatomical variation in relation to latitude and altitude. Blumea 22: 101–121. [ Links ]
Kantay, R., N. As, and O. Unsal. 2000. The mechanical properties of walnut (Juglans regia L.) wood. Turkish J. Agric. and For. 24: 751–756. [ Links ]
Lens, F., P. Gasson, E. Smets, and S. Jansen. 2003. Comparative wood anatomy of Epacrids (Styphelioideae, Ericaceae s.l.). Annals of Bot. 91: 835–856. [ Links ]
Leslie, C. A., and G. McGranahan. 1988. Native populations of Juglans regia – a draft. In: Ozelkok, S., and M. Buyukyilmaz (eds). International Conference on Walnuts. Ataturk Central Horticultural Research Institute Pub. Yalova–Turkey. pp: 111–124. [ Links ]
Liu, J., and S. Noshiro. 2003. Lack of latitudinal trends in wood anatomy of Dodonea viscosa (Sapindaceae), a species with a worldwide distribution. Am. J. Bot. 90: 532–539. [ Links ]
Merev, N. 1998. Wood anatomy of natural Angiospermae taxa in Eastern Black Sea Region I–A. Black Sea Technical University – Forestry Faculty Press. Trabzon. 396 p. [ Links ]
Merev, N., and H. Yavuz. 2000. Ecological wood anatomy of Turkish Rhododendron L. (Ericaceae). Intraspecific variation. Turkish J. Bot. 24: 227–237. [ Links ]
Noshiro, S., and M. Suzuki. 1995. Ecological wood anatomy of Nepalese Rhododendron (Ericaceae). 2. Intraspecific variation. J. Plant Res. 108: 217–233. [ Links ]
Noshiro, S., L. Joshi, and M. Suzuki. 1994. Ecological wood anatomy of Alnus nepalensis (Betulaceae) in East Nepal. J. Plant Res. 107: 399–408. [ Links ]
Roderick, M. L., and S. L. Berry. 2001. Linking wood density with tree growth and environment: a theoretical analysis based on the motion of water. New Phytologist 149: 473–485. [ Links ]
Rodriguez, S. A., T. Terrazas, and L. L. Mata. 2006. Anatomical wood variation of Buddleja cordata (Buddlejaceae) along its natural range in México. Trees 20: 253–261. [ Links ]
Sanli, I. 1977. Anatomical Researches on Wood of Fagus orientalis Lipsky. Growing in Different Regions of Turkey. Matbaa Teknisyenleri Press. Istanbul. 282 p. [ Links ]
Sanli, I. 1989. Le bois et l'archéologie. Revue De La Faculté Forestière De L'Université D'Istanbul 39: 130–141. [ Links ]
Suzuki, M., S. Noshiro, A. Takahasi, K. Yoda, and L. Joshi. 1991. Wood structure of Himalayan plants. In: Ohba, H., and S. B. Malla (eds). The Himalayan Plants II. University of Tokyo Press. pp: 17–65. [ Links ]
Sykes, J. T. 1975. The influence of climate on the regional distribution of nut crops in Turkey. Econ. Bot. 29:108–115. [ Links ]
Thomas, D. S., K. D. Montagu, and J. P. Conroy. 2004. Changes in wood density of Eucalyptus camaldulensis due to temperature: the physiological link between water viscosity and wood anatomy. For. Ecol. and Manag. 193: 157–165. [ Links ]
Verani, S. 1993. Some mechanical characteristics and wood seasoning in Prunus avium, Juglans regia and Robinia pseudoacacia. Quaderni di Ricerca–Centro di Sperimentazione Agricola e Forestale / Instituto di Sperimentazione per la Pioppicoltura. No. 35, 5 p. [ Links ]
Voulgaridis, V., and V. G. Vassiliou. 2006. The walnut wood and its utilisation to high value products. Acta Horticulturae 705: 69–81. [ Links ]
Wodzicki, T. J. 2001. Natural factors affecting wood structure. Wood Sci. and Technol. 35: 5–26. [ Links ]
Xinying, Z., D. Liang, and P. Baas. 1988. The ecological wood anatomy of the Lilacs (Syringa oblata var. graldii) on mount Taibei in North–western China. IAWA Bulletin n.s. 9: 24–30. [ Links ]
Yaltirik, F. 1970. Comparison of anatomical characteristics of wood in Turkish maples with relation to the humidity of the sites. J. Wood Sci. 5: 43–48. [ Links ]
Yaltirik, F. 1971. Taxonomical Study on the Macro –and Micro–morphological Characteristics of Indigenous Maples (Acer L.) in Turkey. Istanbul Univesity Press. Istanbul. 232 p. [ Links ]
Yaltirik, F. 1982. Juglans L. In: Davis, P. H. (ed). Flora of Turkey and the East Aegean Islands. University Press. Edinburgh. pp: 654. [ Links ]
Yaman, B. 2006. Variations in quantitative vessel element characters of Cerasus avium (Rosaceae) wood in Euxine province of Turkey. In: Peev, D. (ed). IV Balkan Botanical Congress (Book of abstracts). Sofia–Bulgaria. pp: 24. [ Links ]
Yaman, B., and M. Saribas. 2004. Vessel size variability of Poplar (Populus L.) species in relation to altitude in Euxine Region of Turkey. Faculty of For. J. Suleyman Demirel University A (1): 111–123. [ Links ]

