











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El término arvense es coloquialmente desconocido por la gente que no está relacionada con sistemas de producción agrícolas. Por el contrario, el termino malezas, malas hierbas o hierbas indeseables; fácilmente es identificado por la mayoría de las personas, lo cual conlleva per se a una connotación negativa acerca de la hierba, mientras que el término arvense simplemente denomina aquellas plantas acompañantes, espontáneas o adventicias asociadas a los cultivos en los espacios modificados para actividades agrícolas (Chacón y Gliessman, 1982). En esta tesitura, en las últimas décadas se han identificado valores positivos de las arvenses en los agroecosistemas, entre los que cabe resaltar su influencia en el control de la erosión del suelo, valor como fuente de alimento de fauna silvestre o doméstica, reserva de germoplasma con utilidad potencial y papel hospedante de enemigos naturales de plagas de los cultivos entre otros (Pareja, 1986; Rivera y Gómez, 1992; Gliessman, 1998; Jones, 2017; Mateos-Maces et al., 2020). Los estudios biológicos acerca de arvenses incluyen conocimientos botánico-taxonómicos, ya que a través de una correcta identificación de especies es posible generar información sobre sus características morfológicas y fisiológicas, lo cual es fundamental para determinar los estadios más vulnerables en cuanto a épocas de aparición, medios de propagación y germinación de semillas (Pareja, 1986), útiles en el diseño de métodos y programas de manejo de la arvenses; sobre todo, para el correcto control mediante métodos químicos, mecánicos, culturales o biológicos.
Conforme a lo descrito, es pertinente agregar que cuando los cafetales están libres de arvenses por períodos muy prolongados, pueden tener problemas tan serios como si se mantuvieran completamente enyerbados, de forma tal que donde el manejo de las arvenses se hace con fines de erradicación completa, puede causarse deterioro en el suelo al exponerse éste a la erosión y lixiviación de nutrientes (Salazar e Hincapié, 2007). También en algunos casos podrían presentarse cambios en la dinámica poblacional de especies, dando lugar a la aparición de otras menos comunes cuyas características las hagan más difíciles de controlar. No obstante, debido a la alternativa del control químico y el relativo bajo costo de este ante el control manual en asociación con otros métodos; se ha dado lugar en ciertas tipologías de productores, a un uso irracional de herbicidas en el control de arvenses, sobre todo cuando hay escases de mano de obra. El contrarrestar poblaciones de arvenses en el cafetal no es una práctica sencilla, si se considera a éstas como el subsistema del estrato bajo del agroecosistema café, cuya presencia implica una interacción compleja de factores de orden ecológico. En este sentido, los estudios de ecología de poblaciones de arvenses están centrándose en el marco de programas de manejo integrado de arvenses (MIA), lo cual puede contribuir notablemente al desarrollo de modelos que permitan más de una estrategia para reducir densidades y coberturas de la comunidad vegetal presente en el sotobosque del cafetal (Salazar e Hincapié, 2009).
Un sistema de MIA enfoca el problema hacia la utilización de formas compatibles con el ambiente, procurando que, mediante la interacción de técnicas adecuadas y conocimientos existentes, la regulación de la comunidad de arvenses se efectúe a niveles tales que los daños económicos que produzcan estén por debajo de un umbral económico aceptable (Salazar-Gutiérrez, 2020). Sin embargo, dado que la falta de solución al problema de las arvenses aún persiste, el MIA a través de estudios básicos de biología y ecología de arvenses, puede incorporar métodos físicos, químicos, mecánicos, entre otros; en conjunto con medidas preventivas. Un enfoque en el diseño de programas eficaces para el control de las arvenses contempla conocer con exactitud las especies que están presentes en el cafetal, a fin de contribuir en su manejo y no en su erradicación (Salazar e Hincapié, 2007). Por lo que el presente trabajo tiene como finalidad contribuir al conocimiento sobre las especies de arvenses más comunes en los cafetales de la zona centro del estado de Veracruz, analizar su estructura, diversidad y parámetros edafoclimáticos que influyen en su distribución.
Área de estudio
El trabajo se realizó en la zona centro del estado de Veracruz (ZCEV) ya que es la principal zona compacta productora de café en México y representa el 96.4 por ciento de la superficie cultivada con café en el estado (López et al., 2021). Para el propósito de este trabajo se dividió la ZCEV en siete regiones productoras de café, que obedecen a características agroecológicas y factores político-administrativos relacionados con aspectos gubernamentales. Para análisis y planeación estas regiones se agruparon en tres subzonas: la Norte (regiones de Atzalan y Misantla), Centro (regiones de Coatepec y Huatusco) y Sur (regiones de Córdoba, Tezonapa y Zongolica) (Figura 1).

Metodología
Se levantó un inventario de la composición florística de arvenses presentes en el sotobosque de cafetales en el periodo de julio a septiembre del año 2013. El muestreo se analizó por subzona, región y sitio de muestro. Los sitios de muestreos dentro de cada región (Figura 1) se hicieron tratando de seguir un transecto altitudinal para encontrar diferencias en la presencia de especies. Se georreferenció su ubicación con ayuda de un GPS marca Garmin modelo 60cs para obtener posteriormente datos de mapotecas digitales, acerca de las características ambientales de estos puntos.
En cada sitio de muestreo se establecieron de manera aleatoria tres rectángulos de un metro cuadrado (2 m x 0.5 m), con base en la metodología de cuadrantes rectangulares propuesta por García (2008) y Caamal-Maldonado y Castillo-Caamal (2011). Se censaron todos los individuos presentes; determinando su identidad taxonómica recolectando ejemplares botánicos que fueron secados y prensados de acuerdo con la metodología propuesta por Pennington y Gentry (1994) para su ingreso y determinación por comparación en el herbario XAL (Instituto de Ecología A. C.), apoyados de claves taxonómicas como las de Schmid (1989), Sánchez-Ken et al. (2012), Soreng et al. (2015) y Sanchez-Ken (2018). Las plantas colectadas debían cumplir ciertos requisitos: la mayoría de sus hojas estar en buen estado, con flores, raíz, de ser posible con fruto y no presentar daño por algún agente patógeno. Se realizó un listado de nombres comunes de las especies de arvenses con la ayuda de los pobladores de las comunidades visitadas el cual se usó en la determinación al consultarlos con un catálogo de nombres vulgares y posteriormente corroborarlas en el herbario (Flores-Olvera y Lindig-Cisneros, 2005).
Se estimó a diferentes niveles (sitio de muestreo, región, subzona y general) las variables de densidad (número de individuos de la especie “i” / área muestreada), frecuencia (número de sitios con presencia de la especie “i” / número total de sitios muestreados) y cobertura (área total proyectada por la especie “i” / área muestreada) por especie y por familia. Estas variables discretas son determinadas a partir de unidades puntuales y su combinación permite una inspección detallada de la dinámica de las especies dentro de un agroecosistema (Aleman, 2004).
Posteriormente se calculó el Índice de Valor de importancia (IVI= densidad relativa + frecuencia relativa + cobertura relativa) de acuerdo con la metodología señalada por Anzalone y Silva (2010). Se elaboró la curva de acumulación de especies para corroborar la fiabilidad del muestreo graficando el número de especies encontradas respectos a los muestreos realizados y con la ayuda del programa EstimateS versión 9.1.0 se calculó la riqueza promedio con el estimador no paramétrico de Chao 1 (basado en la abundancia de cada especie) con la finalidad de precisar el número de especies esperadas para este trabajo (Colwell, 2013). El cálculo de este estimador se generó a partir de un intervalo de 95% de confianza para determinar la significancia estadística de los valores. Por último, se estableció el orden de importancia y se discriminaron especies raras (< al 5% de presencia), que es un criterio sugerido para estudios florísticos (Anzalone y Casanova, 2004).
Se llevaron a cabo análisis de agrupamiento y ordenación haciendo un ajuste de acuerdo con lo sugerido por McCune y Grace (2002); discriminándose aquellas especies con porcentajes menores al 5% del total de sitios muestreados. Para este caso fueron las que solo se presentaron en un sitio de muestreo, por lo que, de las 167 especies registradas se descartaron 80 (especies raras). Llevando a cabo los análisis que a continuación se describen con un total de 87 especies únicamente.
Analisis Cluster (CA). Se realizó con el programa PAST (2007), para relacionar las regiones productoras de café respecto a su similitud florística de ausencia-presencia, empleando UPGMA (unweighted pair-group method with arithmetic averaging) como medida de agrupamiento y el índice de similitud de Sorensen como medida de proximidad. Se eligió este índice por ser de los más robustos para datos ecológicos ya que introduce relativamente poca distorsión en la distancia entre agrupamientos con respecto a la matriz de distancia original y evita el efecto de encadenamiento generado con otros métodos de unión (Taylor et al., 1987; Ludwig y Reynolds, 1989).
Análisis de correlación canónica (CCA). Para analizar los factores ambientales relacionados con la distribución de las arvenses en los cafetales. Se utilizó una matriz de abundancia de especies por sitio y otra con las características ambientales de esos sitios. Se realizaron dos análisis de CCA; con y sin especies raras, se determinó mediante la comparación del porcentaje de la varianza explicada por los ejes principales, que al descartar especies el porcentaje explicado es mayor. Por tal razón únicamente se muestra este CCA. La primera matriz contenía 87 especies (no raras) y la segunda cuatro variables ambientales (temperatura media anual, precipitación media anual, altitud y profundidad del suelo).
Resultados
Riqueza de especies, familias y estructura
En las fincas de la zona centro de estado de Veracruz se cuantificaron un total 5,132 individuos distribuidos en 167 morfotipos de arvenses (Tabla 1), de cuales se pudo determinar taxonómicamente hasta nivel de especie 148 (97.5% de individuos); hasta género 11 (1.0% de individuos) y ocho fueron desconocidas (1.5% de individuos) debido a la carencia de estructuras reproductiva. Las especies identificadas quedaron enmarcadas en 121 géneros y 52 familias; ocho familias acumularon alrededor del 45 por ciento del total de especies; Poaceae (la más rica) con 21 especies, Asteraceae con 20 especies, Lamiaceae con ocho especies, Convolvulaceae y Fabaceae con siete especies cada una, Euphorbiaceae, Malvaceae y Solanaceae con seis especies cada una.
Tabla 1 Listado de familias y especies encontradas en la ZCEV.Table 1. List of families and species found in the ZCEV.
| Familia | Especie (Nombre científico) | Clave | Nombre común |
|---|---|---|---|
| Acanthaceae | Hypoestes phyllostachya Baker | Hyph | Hierba pinta, hoja pinta, percalillo |
| Acanthaceae | Thunbergia alata Bojer ex Sims | Thal | Gorrito (bejuco) |
| Aizoaceae | Trianthema portulacastrum L. | Trpo | Verdolaga cimarona |
| Amaranthaceae | Amaranthus hybridus L. | Amhy | Quelite |
| Amaranthaceae | Amaranthus spinosus L. | Amsp | Quelite |
| Amaranthaceae | Cyathula achyranthoides (Kunth) Moq. | Cyac | Cadillo de bola |
| Anacardiaceae | Toxicodendron radicans (L.) Kuntze | Tora | Caquis, caquixtle, corralilla |
| Anemiaceae | Anemia phyllitidis (L.) Sw. | Anph | Espiga larga, helecho |
| Apiaceae | Eryngium nasturtiifolium Juss. ex F. Delaroche | Ermo | Cilantro cimarrón |
| Apiaceae | Sanicula liberta Cham. & Schltdl. | Sali | 5 hojas |
| Apiaceae | Spananthe paniculata Jacq. | Sppa | Hierba apestosa, sabañon, tallo hueco, zamañon, tronadora |
| Apocynaceae | Asclepias curassavica Griseb. | Ascu | Mata ganado |
| Araceae | Anthurium sp. | Ansp | Hoja corazón |
| Araceae | Caladium bicolor (Aiton) Vent. | Cabi | Mafafa |
| Araceae | Syngonium neglectum Schott | Syne | Lengua de vaca, malaste, malastle, malastre, malaxtle |
| Araceae | Xanthosoma robustum Schott | Xaro | Mafafa |
| Asteraceae | Ageratum corymbosum Zuccagni | Agco | Zopilote |
| Asteraceae | Ageratum houstonianum Mill. | Agho | Hierba del zopilote |
| Asteraceae | Bidens pilosa L. | Bipi | Amozo, mozote blanco |
| Asteraceae | Conyza canadensis (L.) Cronquist | Coca | Vara |
| Asteraceae | Eclipta alba (L.) Hassk. | Ecal | S/N |
| Asteraceae | Eclipta prostrata (L.) L. | Ecpr | Escobo |
| Asteraceae | Elephantopus tomentosus L. | Elto | Lengua de perro, oreja de cochino |
| Asteraceae | Galinsoga parviflora Cav. | Gapa | Arrocillo |
| Asteraceae | Lactuca intybacea Jacq. | Lain | Lechuguilla |
| Asteraceae | Melampodium divaricatum (Rich.) DC. | Medi | Amozo amarillo, hierbabuenilla, mozote amarillo |
| Asteraceae | Melampodium microcephalum Less. | Memi | Flor amarilla |
| Asteraceae | Parthenium hysterophorus L. | Pahy | Altamisa, artemisa |
| Asteraceae | Porophyllum macrocephalum DC. | Poma | Pápalo |
| Asteraceae | Pseudoelephantopus spicatus (B. Juss. ex Aubl.) Rohr ex Gleason | Pssp | Lengua de perro, oreja de burro, totopillo |
| Asteraceae | Schistocarpha bicolor Less. | Scbi | S/N |
| Asteraceae | Senecio vulgaris L. | Sevu | Brujita, cabezuela, lechuga, lechuguilla, lechuguilla erecta |
| Asteraceae | Simsia amplexicaulis (Cav.) Pers. | Siam | Hierba amarilla, hierba amarilla cabezona |
| Asteraceae | Taraxacum officinale F.H. Wigg. | Taof | Diente de león |
| Asteraceae | Tithonia tubaeformis (Jacq.) Cass. | Titu | Cachampa, chimalaca, chimalaco, gigantón, orejona, tabaquillo, tlancahuale (nahuatl), zacaxte-chimalaco |
| Asteraceae | Youngia japonica (L.) DC. | Yoja | Lechuguilla |
| Balsaminaceae | Impatiens mexicana Rydb. | Imme | China gachupina, gachupina |
| Basellaceae | Anredera ramosa (Moq.) Eliasson | Anra | Bejuco baboso, bejuco malo, corrihuela de camote |
| Campanulaceae | Lobelia cardinalis L. | Loca | Flor roja mirto, mirto, vara roja mirto |
| Campanulaceae | Lobelia laxiflora Kunth | Lola | Aretillo, pasmo chilillo |
| Cannabaceae | Trema micrantha (L.) Blume | Trmi | Mata caballo |
| Caryophyllaceae | Drymaria cordata (L.) Willd. ex Schult. | Drco | Berrillo, oreja de ratón, hoja corazón, maltanche, platina |
| Caryophyllaceae | Stellaria cuspidata Willd. ex D.F.K. Schltdl. | Stcu | Maltanche |
| Colchicaceae | Colchicum autumnale L. | Coau | Cebollina |
| Commelinaceae | Commelina coelestis Willd. | Coco | Matlale, matlali, matlalin |
| Commelinaceae | Commelina diffusa Burm. f. | Codi | Hierba del pollo |
| Commelinaceae | Commelina erecta L. | Coer | Hierba del pollo, matlali |
| Commelinaceae | Tripogandra serrulata (Vahl) Handlos | Trse | Matlali de flor blanca, matlali flor blanca |
| Convolvulaceae | Cuscuta corymbosa Ruiz & Pav. | Cuco | Tripa de judas, tripa de pollo |
| Convolvulaceae | Dichondra micrantha Urb. | Dimi | Oreja de ratón |
| Convolvulaceae | Ipomoea batatas (L.) Lam. | Ipba | Camotillo |
| Convolvulaceae | Ipomoea nil (L.) Roth | Ipni | Bejuco, bejuco lechocilla |
| Convolvulaceae | Ipomoea purpurea (L.) Roth | Ippu | Bejuco, bejuco cola de ratón |
| Convolvulaceae | Ipomoea sp. | Ipsp | Nueve hojas |
| Convolvulaceae | Merremia quinquefolia (L.) Hallier f. | Mequ | Bejuco cinco hojas |
| Crassulaceae | Bryophyllum pinnatum (Lam.) Oken | Brpi | Oreja de burro, siempre viva |
| Cucurbitaceae | Cyclanthera ribiflora (Schltdl.) Cogn. | Cyri | Meloncillo, pepinillo, sandia de ratón |
| Cucurbitaceae | Momordica charantia L. | Moch | Bejuco 3 hojas |
| Cucurbitaceae | Sicyos deppei G. Don | Side | Calabacillo |
| Cyperaceae | Cyperus rotundus L. | Cyro | Zacate |
| Cyperaceae | Kyllinga brevifolia Rottb. | Kybr | Coquillo, coquito |
| Cyperaceae | Rhynchospora radicans (Schltdl. & Cham.) H. Pfeiff. | Rhra | Cebollinilla, pasto, zacate chiche |
| Dennstaedtiaceae | Dennstaedtia distenta (Kunze) T. Moore | Dedi | Pesma |
| Euphorbiaceae | Acalypha alopecuroidea Jacq. | Acal | Chinahuatillo, hierba del cáncer, hierba del pastor |
| Euphorbiaceae | Cnidoscolus multilobus (Pax) I.M. Johnst. | Cnmu | Mala mujer |
| Euphorbiaceae | Euphorbia heterophylla L. | Euhe | Lechocilla, nochebuena, lechocilla |
| Euphorbiaceae | Euphorbia hirta L. | Euhi | Lechocilla, lechoza |
| Euphorbiaceae | Euphorbia sp. | Eusp | Lechocilla |
| Euphorbiaceae | Ricinus communis L. | Rico | Higuerilla |
| Hypoxidaceae | Hypoxis decumbens L. | Hyde | Cebollina, lirio amarillo |
| Heliotropiaceae | Heliotropium indicum L. | Hein | Hierba alacrán, hierba del alacrán |
| Lamiaceae | Clerodendrum philippinum Schauer | Clph | S/N |
| Lamiaceae | Hyptis capitata Jacq. | Hyca | Cabezuela, tabardillo |
| Lamiaceae | Hyptis mutabilis (Rich.) Briq. | Hymu | Chichan |
| Lamiaceae | Leonurus sibiricus L. | Lesi | Mariguanilla |
| Lamiaceae | Ocimum micranthum Willd. | Ocmi | Hierbabuenilla, zopilotillo |
| Lamiaceae | Salvia hispanica L. | Sahi | Chante, cola de borrego |
| Lamiaceae | Salvia hyptoides M. Martens & Galeotti | Sahy | Chante, |
| Lamiaceae | Salvia tiliifolia Vahl | Sati | Hierba de rana |
| Fabaceae | Arachis pintoi Krapov. & W.C. Greg. | Arpi | Cacahuatillo |
| Fabaceae | Desmodium adscendens (Sw.) DC. | Dead | Cadillo, pega pega |
| Fabaceae | Desmodium sp. | Desp | Pega pega |
| Fabaceae | Enterolobium cyclocarpum (Jacq.) Griseb | Ency | Guanacaxtle |
| Fabaceae | Leucaena leucocephala (Lam.) de Wit | Lele | Guaje |
| Fabaceae | Mimosa pigra L. | Mipi | Zarza |
| Fabaceae | Mimosa pudica L. | Mipu | Pinahuixtle |
| Malvaceae | Heliocarpus americanus L. | Heam | Tecolixtle |
| Malvaceae | Heliocarpus sp. | Hesp | Cadillo de árbol, jonote |
| Malvaceae | Hybiscus sp. | Hysp | S/n |
| Malvaceae | Sida acuta Burm. f. | Siac | Cadillo, escobilla, escobillo |
| Malvaceae | Waltheria indica L. | Wain | Escobilla, escobillo, escobillo real |
| Malvaceae | Wissadula amplissima (L.) R.E. Fr. | Wiam | Tronadora |
| Melastomataceae | Miconia argentea (Sw.) DC. | Miar | Teshuate |
| Meliaceae | Trichilia havanensis Jacq. | Trha | Rama tinaja |
| Onagraceae | Lopezia racemosa Cav. | Lora | S/N |
| Onagraceae | Oenothera rosea L'Hér. ex Aiton | Oero | Hierba del ángel, hierba del cáncer, hierba del golpe |
| Oxalidaceae | Oxalis corniculata L. | Oxco | Trébol chico |
| Oxalidaceae | Oxalis latifolia Kunth | Oxla | Trébol grande |
| Passifloraceae | Passiflora sp. | Pasp | Granada-maracuyá |
| Petiveriaceae | Rivina humilis L. | Rihu | Coral, verbena |
| Phyllanthaceae | Phyllanthus amarus Schumach. & Thonn. | Pham | Tamarindo |
| Piperaceae | Piper amalago L. | Piam | Listoncillo |
| Piperaceae | Piper marginatum Jacq. | Pima | Como gigantón |
| Piperaceae | Piper sp. | Pisp | Canutillo |
| Piperaceae | Piper umbellatum L. | Pium | Acuyo hoja santa, alajan, hierba santa cimarrona, hoja santa, hoja santa cimarrona, omequelite, tlanopa |
| Piperaceae | Pothomorphe umbellata (L.) Miq. | Poum | Pata de león |
| Plantaginaceae | Plantago major L. | Plma | Llanten |
| Poaceae | Cenchrus echinatus L. | Ceec | Zacate cadillo |
| Poaceae | Cynodon dactylon (L.) Pers. | Cyda | Navajita |
| Poaceae | Dichanthelium acuminatum (Sw.) Gould & C.A. Clark | Diac | Pasto fino |
| Poaceae | Dichanthelium laxiflorum (Lam.) Gould | Dila | Pasto |
| Poaceae | Digitaria ciliaris (Retz.) Koeler | Dici | Gramilla, pasto, pasto conejo, zacate conejo |
| Poaceae | Digitaria insularis (L.) Fedde | Diin | Pasto, zacate varilla |
| Poaceae | Digitaria sp. | Disp | Zacate |
| Poaceae | Echinochloa colona (L.) Link | Ecco | Pasto, zacate |
| Poaceae | Eleusine indica (L.) Gaertn. | Elin | Pasto, pata de gallo |
| Poaceae | Lasiacis procerrima (Hack.) Hitchc. | Lapr | Pasto |
| Poaceae | Oplismenus hirtellus (L.) P. Beauv. | Ophi | Pasto, pasto matlali |
| Poaceae | Panicum laxum Sw. | Palax | Pasto, pasto fino, zacate |
| Poaceae | Panicum maximum Jacq. | Pama | Zacate |
| Poaceae | Panicum mexicanum Scribn. & Merr. | Pame | Pasto |
| Poaceae | Paspalum langei (E. Fourn.) Nash | Pala | Grama, pasto, pasto cola de burro, pasto de burro, zacate, zacate burro |
| Poaceae | Paspalum notatum Flüggé | Pano | Frente de toro |
| Poaceae | Paspalum virgatum L. | Pavi | Frente de toro, pasto |
| Poaceae | Pseudechinolaena polystachya (Kunth) Stapf | Pspo | Panza de burro, pasto, pasto matlali, pasto panza de burro |
| Poaceae | Rottboellia cochinchinensis (Lour.) Clayton | Roco | Pasto aguatoso, pasto invasor |
| Poaceae | Setaria geniculata P. Beauv. | Sege | Cola de zorra, pasto chinahuatillo, zacate, zacate chinahuatillo, zacate gusano |
| Poaceae | Urochloa plantaginea (Link) R.D. Webster | Urpl | Pasto |
| Polygalaceae | Polygala paniculata L. | Popa | Arrocillo |
| Polygonaceae | Rumex acetosella L. | Ruac | Lengua de vaca |
| Polygonaceae | Rumex obtusifolius L. | Ruob | Lengua de vaca |
| Polypodiaceae | Polypodium sp. | Posp | Helecho, ocopetate, palma helecho, palmilla |
| Portulacaceae | Portulaca oleracea L. | Pool | Verdolaga |
| Primulaceae | Anagallis arvensis L. | Anar | Maltanche |
| Rosaceae | Eriobotrya japonica (Thunb.) Lindl. | Erja | Níspero |
| Rubiaceae | Borreria alata (Aubl.) DC. | Boal | Cacahuatillo |
| Rubiaceae | Diodia brasiliensis Spreng. | Dibr | Escobillo chino, romerillo |
| Rubiaceae | Hamelia patens Jacq. | Hapa | Aretillo, carne de perro, cadillo, canilla de pollo, canutillo, chilillo, cacahuatillo, hierba de negro, tres hojas, trompetilla, cacahuapaxtle |
| Rubiaceae | Richardia scabra L. | Risc | Cacahuatillo |
| Scrophulariaceae | Buddleja cordata Kunth | Buco | Hierba apestosa |
| Selaginellaceae | Selaginella apoda (L.) C. Morren | Seap | Doradillo, helecho, liquen, pesmilla |
| Solanaceae | Capsicum annuum L. | Caan | Chiltepín |
| Solanaceae | Physalis angulata L. | Phan | Jaltomate, tomatillo |
| Solanaceae | Solanum americanum Mill. | Soam | Cenizo |
| Solanaceae | Solanum nigrescens M. Martens & Galeotti | Soni | Hierba mora, tomatillo-mora |
| Solanaceae | Solanum sp. | Sosp | Ojo de totol |
| Solanaceae | Solanum umbellatum Mill. | Soum | Muela |
| Tectariaceae | Tectaria heracleifolia (Willd.) Underw. | Tehe | Helecho calihuale |
| Thelypteridaceae | Macrothelypteris torresiana (Gaudich.) Ching | Mato | Pesma |
| Urticaceae | Cecropia peltata L. | Cepe | Guarumbo |
| Urticaceae | Pilea hyalina Fenzl | Pihy | Hierbabuenilla, ortiga |
| Urticaceae | Pilea pubescens Liebm. | Pipu | Hierbabuenilla |
| Urticaceae | Urtica chamaedryoides Pursh | Urch | Chichicaxtle |
| Urticaceae | Urtica dioica L. | Urdi | Chichicaxtle, hierba buenilla |
| Verbenaceae | Aloysia triphylla Royle | Altr | Hoja nervada |
| Verbenaceae | Lantana camara L. | Laca | Orozus |
| Verbenaceae | Lantana hirta Graham | Lahi | Orozus |
| Viburnaceae | Sambucus canadensis L. | Saca | Hierba de espanto, sauco |
| Viburnaceae | Sambucus nigra L. | Sani | Sauco |
| No determinada | No identificado 1 | Noi1 | Cordoncillo |
| No determinada | No identificado 2 | Noi2 | 3 hojas |
| No determinada | No identificado 3 | Noi3 | Totolquelite |
| No determinada | No identificado 4 | Noi4 | Copalillo |
| No determinada | No identificado 5 | Noi5 | Cadillo |
| No determinada | No identificado 6 | Noi6 | S/N |
| No determinada | No identificado 7 | Noi7 | Tipo helecho |
| No determinada | No identificado 8 | Noi8 | S/N |
La subzona centro (Coatepec y Huatusco) resultó tener la mayor riqueza en especies y familias con el 73 y 84.9 por ciento respectivamente del total (Figura 2). La subzona con el menor número de especies y familias encontradas fue la sur (Córdoba, Zongolica y Tezonapa). En el caso de especies y familias compartidas, el mayor número de coincidencias fue entre las subzonas norte y centro con 57 especies y 29 familias. Únicamente 33 especies y 22 familias tuvieron presencia en las todas las subzonas (Figura 2). La subzona centro presentó el mayor número de familias y especies exclusivas; la subzona norte y la sur el menor en especies y familias respectivamente.
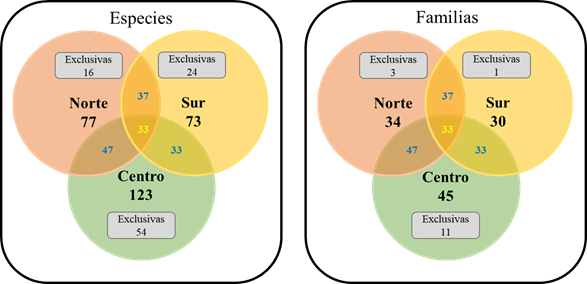
Figura 2 Diagramas de Venn mostrando la distribución de especies y familias entre subzonas de la ZCEV.
Tabla 2 Familias de arvenses no compartidas entre subzonas. Table 2. Weed families not shared among subzones.
| Subzona | Familias Exclusivas |
|---|---|
| Norte | Cannabaceae, Liliaceae, Primulaceae |
| Centro | Aizoaceae, Ameniaceae, Boraginaceae, Melastomataceae Meliaceae, Passifloraceae, Polygalaceae, Portulacaceae, Rosácea, Scrophulariaceae |
| Sur | Dennstaedtiaceae |
Tabla 3 Especies no compartidas entre subzonas.Table 3. Species not shared among subz ones.
| Subzona | Especies exclusivas |
|---|---|
| Norte | Anagallis arvensis, Cnidoscolus multilobus, Colchicum autumnale, Dichanthelium acuminatum, Dichanthelium laxiflorum, Heliocarpus americanus, Lactuca intybacea, Lobelia cardinalis, Mimosa pudica, Paspalum notatum, Salvia tiliifolia, Sambucus nigra, Solanum americanum, Stellaria cuspidate, Trema micrantha. |
| Centro | Ageratum corimbosum, Ageratum houstonianum, Aloysia tryphylla, Amaranthus spinosus, Anemia phyllitidis, Anthurium sp., Buddleja cordata, Cecropia peltata, Cenchrus equinatus, Commelina difusa, Desmodium adscendens, Digitaria insularis, Digitaria sp., Echinochloa colonum, Eclipta alba, Eclipta postrata, Eleusine indica, Enterolobium cyclocarpum, Eriobotrya japonica, Eryngium nasturtiifolium, Euphorbia hirta, Euphorbia sp., Heliotropium indicum, Ipomoea sp., Lantana camara, Lantana hirta, Lasiacis procerrima, Leucaena leucocephala, Lopezia racemosa, Melampodium microcephalum, Merremia quinquefolia, Miconia argentea, Mimosa pigra, Momordica charantia, Panicum mexicanum, Passiflora sp., Pilea pubescens, Piper amalago, Piper marginatum, Polygala paniculata, Porophyllum macrocephallum, Portulaca oleracea, Rottboellia cochinchinensis, Rumex acetosella, Sanicula liberta, Sicyos deppei, Solanum sp., Taraxacum officinale, Tectaria heracleifolia, Triantema portulacastrum, Trichilia havanenesis, Urtica chamaedryodes, Wissadula amplissima |
| Sur | Borreria alata, Caladium bicolor, Clerodendrum philliphinum, Cyathula achyranthoides, Cynodon dactylon, Dennstaedtia distinta, Desmodium sp., Hybiscus sp., Hyptis mutabilis, Ipomoea batatas, Leonurus sibiricus, Macrothelypteris torresiana, Panicum maximum, Salvia hyptoides, Schistocarpha bicolor, Simsia amplexicaulis, Solanum umbellatum, Thunbergia alata |
Los valores del estimador Chao 1 calculados para el total de muestreos realizados en el área de estudio es de 219.52 (línea punteada en la Figura 3) existiendo una diferencia de 52 especies con respecto a las registradas en la ZCEV (línea sólida en la Figura 3), por lo que se esperaría la adición de nuevas especies al listado florístico.

Figura 3 Curva de acumulación de especies para la ZCEV y cálculo de estimador Chao1 con un intervalo de confianza del 95%.
En cuanto a las familias con los valores más altos de IVI son: Commelinaceae seguida de Asteraceae, Acanthaceae y Poaceae, acumulando estas cuatro casi el 25% del total calculado (Tabla 4).
Tabla 4 Índice de Valor de Importancia (IVI) de las principales familias. Table 4. Importance Value Index (IVI) of the main families.
| Familia | Densidad | Cobertura | Frecuencia | IVI | |||
|---|---|---|---|---|---|---|---|
| (Ind/m2) | Relativa | (%/m2) | Relativa | Absoluta | Relativa | ||
| Commelinaceae | 26.8 | 10.4 | 11.2 | 7.5 | 23 | 6.5 | 8.2 |
| Asteraceae | 15.6 | 6.1 | 6.7 | 4.5 | 26 | 7.4 | 6.0 |
| Poaceae | 14.7 | 5.7 | 6.2 | 4.2 | 25 | 7.1 | 5.7 |
| Acanthaceae | 15.4 | 6.0 | 11.8 | 7.9 | 5 | 1.4 | 5.1 |
| Crassulaceae | 18.0 | 7.0 | 5.5 | 3.7 | 8 | 2.3 | 4.3 |
| Urticaceae | 11.1 | 4.3 | 5.7 | 3.8 | 16 | 4.5 | 4.2 |
| Malvaceae | 6.5 | 2.5 | 6.3 | 4.2 | 20 | 5.7 | 4.1 |
| Oxalidaceae | 17.9 | 7.0 | 1.9 | 1.3 | 7 | 2.0 | 3.4 |
| Basellaceae | 3.0 | 1.2 | 11.0 | 7.4 | 3 | 0.9 | 3.1 |
| Rubiaceae | 4.2 | 1.6 | 3.5 | 2.4 | 19 | 5.4 | 3.1 |
| Cyperaceae | 7.0 | 2.7 | 2.6 | 1.8 | 17 | 4.8 | 3.1 |
| Araceae | 4.8 | 1.8 | 3.7 | 2.5 | 16 | 4.5 | 3.0 |
| Apiaceae | 6.9 | 2.7 | 4.0 | 2.7 | 12 | 3.4 | 2.9 |
| Caryophyllaceae | 10.4 | 4.1 | 3.9 | 2.6 | 6 | 1.7 | 2.8 |
| Euphorbiaceae | 2.9 | 1.1 | 2.3 | 1.5 | 18 | 5.1 | 2.6 |
| 36 Familias restantes | 91.7 | 35.7 | 62.8 | 42.1 | 131 | 37.2 | 38.2 |
| Total | 256.9 | 100.0 | 149.0 | 100.0 | 352 | 100.0 | 100.0 |
En el mismo sentido, las especies mejor representadas de manera general en la ZCEV fueron: Bidens pilosa, Commelina coelestis, Melampodium divaricatum, Pseudechinolaena polystachya, Commelina erecta, Paspalum langei, Digitaria ciliaris, Pilea hyalina, Richardia scabra y Bryophyllum pinnatum (Tabla 5).
Tabla 5 Índice de Valor de Importancia (IVI) de las principales especies. Table 5. Importance Value Index (IVI) of the main species.
| Especie (nombre científico) | Densidad | Cobertura | Frecuencia | IVI | |||
|---|---|---|---|---|---|---|---|
| (Ind/m2) | Relativa | (%/m2) | Relativa | Absoluta | Relativa | ||
| Bidens Pilosa | 861 | 16.8 | 237 | 9.3 | 20 | 3.5 | 9.9 |
| Commelina coelestis | 566 | 11.0 | 273 | 10.7 | 22 | 3.9 | 8.5 |
| Melampodium divaricatum | 500 | 9.7 | 263 | 10.3 | 22 | 3.9 | 8.0 |
| Pseudechinolaena polystachya | 295 | 5.7 | 155 | 6.1 | 12 | 2.1 | 4.6 |
| Commelina erecta | 278 | 5.4 | 69 | 2.7 | 6 | 1.1 | 3.1 |
| Paspalum langei | 118 | 2.3 | 84 | 3.3 | 14 | 2.5 | 2.7 |
| Digitaria ciliaris | 119 | 2.3 | 73 | 2.9 | 13 | 2.3 | 2.5 |
| Pilea hyaline | 95 | 1.9 | 72 | 2.8 | 12 | 2.1 | 2.3 |
| Richardia scabra | 65 | 1.3 | 69 | 2.7 | 12 | 2.1 | 2.0 |
| Bryophyllum pinnatum | 144 | 2.8 | 44 | 1.7 | 8 | 1.4 | 2.0 |
| Sida acuta | 68 | 1.3 | 54 | 2.1 | 14 | 2.5 | 2.0 |
| Spananthe paniculata | 68 | 1.3 | 42 | 1.6 | 10 | 1.8 | 1.6 |
| Setaria geniculate | 80 | 1.6 | 41 | 1.6 | 8 | 1.4 | 1.5 |
| Hypoestes phyllostachya | 75 | 1.5 | 58 | 2.3 | 4 | 0.7 | 1.5 |
| Kyllinga brevifolia | 101 | 2.0 | 23 | 0.9 | 6 | 1.1 | 1.3 |
| 152 especies restantes | 1699 | 33.1 | 1002 | 39.2 | 381 | 67.6 | 46.6 |
| Total | 5132 | 100 | 2559 | 100 | 564 | 100 | 100 |
Dentro de las zonas de muestreo se encontraron similitudes en cuanto a las especies registradas, en algunos casos, compartiendo valores similares, tal es el caso de; Bidens pilosa (Asteraceae) que para las subzona centro y sur registra el primer lugar para el cálculo de IVI con 8.1 y 12.5%, respectivamente; Commelina coelestis (Commelinaceae) segundo lugar en la zona centro (7.3%) y norte (11.5%), tercera en la sur (7%); y Melampodium divaricatum (Asteraceae) quinto lugar en la subzona centro (3.7%), primera en la norte (11.9%) y segunda en la sur (9.4%). Dentro de cada subzona se sigue la misma tendencia que en la totalidad de la zona de estudio, en donde otras especies se agregan acaparando los primeros puestos sobresaliendo: Sida acuta (3.3%) y Spananthe paniculata (2.1%) en la subzona centro; Setaria geniculata (3%) en la norte; Hypoestes phyllostachya (3.1%) y Urtica dioica (2.5%) en la sur (Tabla 6).
Tabla 6 Índice de Valor de Importancia (IVI) de las principales especies por subzona.Table 6. Importance Value Index (IVI) of the main species by subzone.
| Especie (nombre científico) | Densidad | Cobertura | Frecuencia | IVI | ||||
|---|---|---|---|---|---|---|---|---|
| (Ind/m2) | Relativa | (%/m2) | Relativa | Absoluta | Relativa | |||
| Subzona centro | Bidens pilosa | 111 | 8.0 | 134 | 13.0 | 9 | 3.4 | 8.1 |
| Commelina coelestis | 122 | 8.8 | 101 | 9.8 | 9 | 3.4 | 7.3 | |
| Paspalum langei | 76 | 5.5 | 59 | 5.7 | 6 | 2.3 | 4.5 | |
| Digitaria ciliaris | 76 | 5.5 | 50 | 4.9 | 8 | 3.1 | 4.5 | |
| Melampodium divaricatum | 56 | 4.0 | 45 | 4.4 | 7 | 2.7 | 3.7 | |
| Pseudechinolaena polystachya | 67 | 4.8 | 43 | 4.2 | 4 | 1.5 | 3.5 | |
| Sida acuta | 47 | 3.4 | 37 | 3.6 | 8 | 3.1 | 3.3 | |
| Pilea hyalina | 34 | 2.4 | 36 | 3.5 | 5 | 1.9 | 2.6 | |
| Spananthe paniculata | 34 | 2.4 | 24 | 2.3 | 4 | 1.5 | 2.1 | |
| Otras 114 | 770 | 55.3 | 499 | 48.5 | 202 | 77.1 | 60.3 | |
| Subzona norte | Melampodium divaricatum | 234 | 16.4 | 126 | 14.9 | 8 | 4.4 | 11.9 |
| Commelina coelestis | 229 | 16.0 | 120 | 14.2 | 8 | 4.4 | 11.5 | |
| Bidens pilosa | 92 | 6.4 | 63 | 7.4 | 7 | 3.9 | 5.9 | |
| Pseudechinolaena polystachya | 96 | 6.7 | 54 | 6.4 | 6 | 3.3 | 5.5 | |
| Richardia scabra | 43 | 3.0 | 55 | 6.5 | 4 | 2.2 | 3.9 | |
| Pilea hyalina | 48 | 3.4 | 28 | 3.3 | 6 | 3.3 | 3.3 | |
| Setaria geniculata | 51 | 3.6 | 26 | 3.1 | 4 | 2.2 | 3.0 | |
| Digitaria ciliaris | 43 | 3.0 | 23 | 2.7 | 5 | 2.8 | 2.8 | |
| Commelina erecta | 56 | 3.9 | 16 | 1.9 | 3 | 1.7 | 2.5 | |
| Otras 68 | 537 | 37.6 | 335.5 | 39.6 | 129 | 71.7 | 49.6 | |
| Subzona sur | Bidens pilosa | 658 | 28.5 | 40 | 5.9 | 4 | 3.3 | 12.5 |
| Melampodium divaricatum | 210 | 9.1 | 92 | 13.5 | 7 | 5.7 | 9.4 | |
| Commelina coelestis | 215 | 9.3 | 52 | 7.6 | 5 | 4.1 | 7.0 | |
| Commelina erecta | 218 | 9.4 | 51 | 7.5 | 2 | 1.6 | 6.2 | |
| Pseudechinolaena polystachya | 132 | 5.7 | 58 | 8.5 | 2 | 1.6 | 5.3 | |
| Bryophyllum pinnatum | 128 | 5.5 | 36 | 5.3 | 3 | 2.5 | 4.4 | |
| Hypoestes phyllostachya | 45 | 1.9 | 33 | 4.8 | 3 | 2.5 | 3.1 | |
| Urtica dioica | 71 | 3.1 | 18 | 2.6 | 2 | 1.6 | 2.5 | |
| No identificado 7 | 45 | 1.9 | 20 | 2.9 | 1 | 0.8 | 1.9 | |
| Otras 64 | 588 | 25.5 | 281.5 | 41.3 | 93 | 76.2 | 47.7 | |
Análisis de correlación canónica
Con base en las coordenadas obtenidas con el GPS de los sitios de muestreo, en la Tabla 7 se muestran datos de suelo obtenidos en mapotecas digitales (INEGI, 2013a) así también de precipitación y temperatura que se obtuvieron a través de promedios de los valores estimados del modelo “raster” de las variables antes mencionadas, interpolados con el método de “Thin plate smoothing spline” del software Anuspline (Hutchinson, 2004) a una resolución de píxel de 90 m. Los datos promediados de altitud provienen del modelo de elevación digital de INEGI de igual resolución (INEGI, 2013b).
Tabla 7 Ubicación geográfica y características edafoclimáticas de los sitios de muestreo. Table 7. Geographical location and soil-climatic characteristics of the sampling sites.
| Clave sitio | Municipio | Localidad | Latitud | Longitud | Altitud (msnm) |
Suelo dominante |
Textura | Temperatura media (°C) |
Precipitación media (mm/año) |
|---|---|---|---|---|---|---|---|---|---|
| ATZ1 | Atzalán | Napoala | 19.898 | -97.2266 | 750 | Andosol | Media | 20 | 2083 |
| ATZ2 | Atzalán | Pilares | 19.9147 | -97.2166 | 679 | Andosol | Media | 20 | 2099 |
| ATZ3 | Atzalán | Arroyo colorado | 19.8922 | -97.2347 | 889 | Andosol | Media | 19 | 2085 |
| ATZ4 | Altotonga | Temimilco | 19.8133 | -97.0971 | 944 | Andosol | Media | 19 | 1811 |
| COA1 | Teocelo | Llano grande | 19.3718 | -96.8741 | 839 | Acrisol | Media | 21 | 1577 |
| COA2 | Teocelo | Llano grande | 19.3718 | -96.8741 | 839 | Acrisol | Media | 21 | 1577 |
| COA3 | Teocelo | Monte blanco | 19.3737 | -96.929 | 985 | Acrisol | Media | 20 | 1812 |
| COA4 | Teocelo | Independencia | 19.3886 | -96.9415 | 1066 | Acrisol | Media | 20 | 1840 |
| COA5 | Teocelo | La virgen | 19.3883 | -96.9905 | 1227 | Acrisol | Media | 18 | 1985 |
| COA6 | Emiliano Zapata | Finca roma | 19.4663 | -96.8763 | 1070 | Acrisol | Fina | 20 | 1472 |
| COA7 | Jilotepec | Vista hermosa | 19.618 | -96.9247 | 1161 | Andosol | Media | 18 | 1592 |
| COA8 | Jilotepec | Vista hermosa | 19.6313 | -96.9266 | 1180 | Litosol | Media | 18 | 1613 |
| COR1 | Atoyac | Los nogales | 18.9647 | -96.8305 | 943 | Litosol | Fina | 20 | 1981 |
| COR2 | Atoyac | Manzanillo | 18.9525 | -96.8262 | 847 | Litosol | Fina | 21 | 1997 |
| COR3 | Fortin | Tlacotengo | 18.9291 | -96.9963 | 1088 | Andosol | Media | 19 | 2083 |
| HUA1 | Huatusco | Ixpila | 19.1188 | -96.9616 | 1154 | Acrisol | Media | 19 | 2015 |
| HUA2 | Tlacotepec | Tlacotepec | 19.1983 | -96.8275 | 850 | Acrisol | Media | 21 | 1566 |
| HUA3 | Huatusco | Capulapa | 19.0977 | -96.8872 | 960 | Acrisol | Media | 20 | 1864 |
| MIS1 | Tenochtitlan | Colorado | 19.8355 | -96.9272 | 970 | Luvisol | Fina | 19 | 1910 |
| MIS2 | Tenochtitlan | Colorado | 19.8388 | -96.9408 | 980 | Luvisol | Fina | 19 | 1917 |
| MIS3 | Misantla | Vicente Guerrero | 19.8561 | -96.9091 | 863 | Luvisol | Fina | 20 | 1934 |
| MIS4 | Juchique de Ferrer | Plan de las hayas | 19.7688 | -96.6605 | 797 | Feozem | Fina | 21 | 1402 |
| MIS5 | Juchique de Ferrer | Plan de las hayas | 19.7513 | -96.675 | 1102 | Feozem | Fina | 19 | 1403 |
| TEZ1 | Tezonapa | Villa nueva | 18.6177 | -96.8241 | 780 | Litosol | Fina | 21 | 2878 |
| ZON1 | Zongolica | Nepopualco | 18.6866 | -96.9913 | 1335 | Acrisol | Media | 18 | 2344 |
| ZON2 | Zongolica | Loma de dolores | 18.6794 | -96.9297 | 892 | Acrisol | Media | 20 | 2562 |
| ZON3 | Zongolica | Xochiatepec | 18.6869 | -96.923 | 822 | Acrisol | Media | 20 | 2566 |
Los resultados del análisis arrojaron que el primer eje explicaba el 43.22 por ciento de la varianza y el segundo 34.05%; lo cual sugiere que hay buena separación entre los sitios de muestreo y variables ambientales a lo largo de los ejes de variación.
La ordenación de los sitios de muestreo y especies, en grupos compactos de sitios que están relacionados con variables ambientales, por ejemplo, en el eje1 la precipitación es la que mayor influencia tiene, en tanto que para el eje 2 es la altitud, ya que como se observa, éstas presentan el menor ángulo respecto al eje. En el eje 1 los sitios más diferentes en cuanto a su composición florística y características ambientales son: ZO3 (extrema derecha) y CO2 (extrema izquierda); el primero con altas precipitaciones y suelos poco profundos, el segundo con características contrarias, sin embargo, los dos tienen una altitud similar. En el eje 2 los sitios más contrastantes son TE1 (superior) y CO7 (inferior), que difieren en altitud, precipitación y temperatura, sin embargo, su profundidad de suelo es muy similar (Figura 4).

Figure 4 Analysis of the relationship between the presence of weeds and the conditions of each sampling site.
Se identificaron cuatro grupos bastante compactos; el primero constituido por sitios de la subzona norte (AT3, AT4, MI1, MI2 y MI3), el segundo en la subzona centro (CO3, CO4, CR3 y HU3), el tercero, aunque más disperso también pertenecen a la subzona centro (HU2, CO2 y CO1), un cuarto es una mezcla de la subzona norte y centro (CO6, CO7, MI4, MI5) y el resto de los sitios están dispersos ya que no presentan semejanza florística con algún otro. El primer grupo (AT3, AT4, MI1, MI2 y MI3) presenta una relación con las variables altitud y precipitación, lo que indica que la semejanza florística puede deberse a estos factores y que especies como Paspalum langei (Pala) y Setaria geniculata (Sege) siguen el mismo patrón de comportamiento; menor altitud y mayor precipitación, además se puede observar la existencia de una relación negativa con la profundidad del suelo en estos sitios. El segundo grupo (CO3, CO4, CR3 y HU3) presenta suelos profundos, pero precipitaciones menores, estos sitios se encuentran a mayor altitud que el primer grupo y las especies que caracterizan a este grupo son Cuscuta corymbosa (Cuco), Senecio vulgaris (Sevu) y Bryophyllum pinnatum (Brpi). El tercer grupo de (CO1, CO2 y HU2) es el grupo más alejado del centro respecto al eje 1, por lo tanto, presenta poca precipitación y suelos profundos, así como temperaturas mayores, la especies que más se asocia con este grupo es: Digitaria ciliaris (Dici), el cuarto grupo (CO6, CO7, MI4, MI5) es más heterogéneo que los demás, en cuanto a composición florística y factores ambientales, la característica ambiental más semejante entre sitios es la baja precipitación, seguido de la baja profundidad del suelo.
Algunas arvenses como Cyperus rotundus (Cyro), Youngia japonica (Yoja), Oenothera rosea (Oero), Cyclanthera ribiflora (Cyri) se encuentran relacionadas con precipitaciones bajas. Por el contrario, Hyptis capitata (Hyca), Salvia hispanica (Sahi), Oplismenus hirtellus (Ophi) son afines a sitios con precipitaciones altas como TE1, ZO3 y ZO1. Aquellas que mayor valor de importancia obtuvieron, aparecen al centro de los ejes como en el caso de Bidens pilosa (Bipi), Commelina coelestis (Coco), Paspalum langei (Pala), Pseudechinolaena polystachya (Pspo), Sida acuta (Siac) y Richardia scabra (Risc), ya que su frecuencia de aparición y distribución es a todo lo largo de la zona centro de estado.
Discusión
Los datos obtenidos sobre la composición florística de arvenses en los cafetales de la ZCEV discrepan con el número de especies identificadas en otros trabajos que analizan estas comunidades vegetales, Sanginés-García et al. (2014) registra 58 especies de arvenses en cafetales bajo un sistema silvopastoril con ovinos y señala que la familia Fabaceae es la mejor representada. De la misma forma, Córdoba-Varón y Patiño-Bocanegra (2017) enlista 56 especies en tres cafetales con diferentes tipos de manejo en el municipio del Líbano, Colombia. El menor número de especies de arvenses puede estar asociado al manejo intensivo que los cafeticultores realizan en sus unidades de producción (sistemas especializados o policultivos simples), con controles químicos y/o mecánicos de las arvenses; el presente estudio fue realizado en cafetales bajo sistemas de policultivo diverso principalmente, donde el control de las arvenses es la actividad más representativa respecto al costo anual de producción (López et al., 2020), por lo que se realiza generalmente una o dos veces por año, conservándose en buena medida la diversidad de especies de arvenses, debido al bajo o nulo manejo. Para el caso de las familias siguen una tendencia similar en trabajos sobre diversidad de arvenses en cafetales, Torres et al. (2001) reconoce a las familias Asteraceae y Poaceae como elementos dominantes en el estrato herbáceo de fincas cafetaleras del municipio de Tlaltetela, Veracruz. En el mismo sentido, Castro-Cepero et al. (2019) refiere a las mismas familias como parte importante del sotobosque de cafetales en el Perú. Ambas se encuentran ampliamente distribuidas a nivel mundial con aproximadamente 11,000 y 23,000 especies respectivamente (Brako y Zarucchi, 1993; Ulloa-Ulloa et al., 2004; Simon, 2007; Rodríguez et al., 2017). La Asteraceae es la familia botánica más extensa en México, con 417 géneros distribuidos en 3,113 especies. Su gran variabilidad fenotípica y genotípica le permite adecuarse a diversas condiciones ambientales, en su mayoría como arvenses (Villaseñor, 2018). Algo similar ocurre con la Poaceae, que para México registra 215 géneros distribuidos en 1,312 especies y que en algunos casos son clasificadas como “arvenses nocivas” tanto para especies cultivadas como nativas (Sánchez-Ken, 2018). La amplia distribución de ambas familias sumado a su gran capacidad de adaptación puede reconocerse en el análisis de ordenación de este estudio, especies como Bidens pilosa (Asteraceae) y Paspalum langei (Poaceae) parecen no seguir ningún patrón ambiental, registrando a su vez altos valores de IVI. Otras especies que siguen la misma tendencia son Melampodium divaricatum (Asteraceae), Pseudechinolaena polystachya (Poaceae) y Commelina erecta (Commelinaceae), esta última correspondiente a la familia con el valor de IVI más alto registrado. Generalmente las Commelinaceae son reconocidas como elementos comunes dentro de cafetales, en algunos casos son consideradas como coberturas valiosas y plantas trampa de ciertos microorganismos, sin embargo, cuando su población excede cierto límite puede propiciar la aparición de ciertos patógenos, principalmente hongos como la roya del cafeto; y plagas de importancia económica como la broca del cafeto y ácaros (Alvarado-Huaman et al., 2019; Sosa et al., 2020). Es importante señalar que así como en algunas circunstancias las arvenses tienen efectos negativos sobre el cafetal como: competencia por nutrientes, espacio y luz, hospederos de organismos perjudiciales, producción de sustancias alelopáticas y encarecer labores culturales dentro de la finca, una gran mayoría contribuyen a la estabilidad de los agroecosistemas y son apreciadas por considerarse plantas forrajeras, melíferas, útiles para la gastronomía, medicinales, hospederas de organismos benéficos, coberteras verdes, retenedoras de humedad y suelo y proveedoras de semillas para aves, lo que les otorga un gran valor ecológico, cultural y económico que los productores de café aprovechan a pequeña o mediana escala y en algunos casos llegan a comercializarlas dentro y fuera de su localidad lo que les genera ingresos extra finca que les ayuda a la sustento en el hogar (Martínez et al., 2007; Sanginés-García et al., 2014). Biden pilosa es una arvense con alta presencia y, persistencia en el cafetal ademas de ser buena fuente de alimento por sus características bromatológicas (López et al., 2021), con más de 201 compuestos identificados en esta planta es capaz de tratar más de 40 trastornos de la salud, con potencial para el tratamiento tumores, inflamación/modulación inmune, diabetes, virus, microbios, protozoos, enfermedades gastrointestinales, hipertensión y enfermedades cardiovasculares (Bartolome et al., 2013). El cálculo del estimador Chao1 muestra que de aumentar el esfuerzo de muestreo es posible seguir adicionando especies nuevas al listado florístico, sin embargo, para lograr un comportamiento asintótico es necesario considerar el número de especies exclusivas o raras en la muestra ya que podría afectar en el número total de especies estimadas. El cafetal por sí solo es un agroecosistema diverso, por lo que es posible que sean necesarias más unidades de muestreo para afinar el inventario florístico (Bandeira et al., 2005; García et al., 2015; López-Gómez y Williams-Linera, 2017).
Las especies con mayor IVI coinciden en las diferentes subzonas de la ZCEV, lo que podría indicar una fuerte persistencia en el cafetal por parte de estas especies y una gran adaptabilidad a las condiciones climáticas presentes en la región, sumado posiblemente a una alta capacidad de reproducción y producción de semillas, facilitando su dispersión (Salazar e Hincapié, 2005; López et al., 2013). Aunque las arvenses son generalmente consideradas cosmopolitas algunas especies presentan un rango de distribución restringido para alguna subzona dentro del área de estudio. La orografía accidentada podría estar generando condiciones particulares que propician el desarrollo de estas especies exclusivas y al mismo tiempo limitar su distribución. Un ejemplo de lo anterior se ve reflejado en algunas de las especies únicas registradas para la subzona sur: Borreria alata, Caladium bicolor, Dennstaedtia distenta, Hyptis mutabilis, Simsia amplexicaulis que requieren precipitaciones abundantes con clima semicálido y vegetación propia de zonas templadas. Esta subzona también presenta una mayor heterogeneidad de valores en las variables edafoclimáticas registradas entre sus localidades, acumula anualmente la mayor precipitación de toda el área de estudio con 2878 mm (Villa Nueva: TE1) y cuentan con la mayor altitud registrada en todos los sitios de muestreo, estas condiciones podrían estar limitando el desplazamiento comunidades de arvenses a la subzona disminuyendo al mismo tiempo el número de total de especies exclusivas (Nepopualco: ZO1) (Santander et al., 2003; Agudelo, 2008; Herzog et al., 2010; Rzedowski et al., 2011; Charrupi-Riascos, 2019). La subzona centro presentó el mayor número de familias exclusivas, es decir, aquellas que solo se registraron en esta zona. Asimismo, las técnicas utilizadas (CA y CCA) reafirman que existe una alta composición de arvenses en los cafetales de la ZCEV siguiendo un patrón de distribución influido principalmente por el gradiente altitudinal y precipitación, y en menor medida por las condiciones de sitio tales como la profundidad del suelo.
Conclusiones
En la zona centro del estado de Veracruz existe una elevada riqueza de arvenses, siendo Bidens pilosa, Commelina coelestis y Melampodium divaricatum, las de mayor valor de importancia, al igual que las familias Commelinaceae, Asteraceae y Poaceae; en ambos casos acumulando alrededor de una cuarta parte del total, lo que representa una comunidad con baja dominancia de unas pocas especies o familias.
Se registró un elevado número de especies con escasa representatividad, así como especies exclusivas; siendo la subzona centro la que registró más (54 especies); lo que expone la necesidad de realizar más estudios de manera periódica, para identificar posibles cambios en la composición florística; tomando como premisa que estas especies son más sensibles a perturbaciones ambientales.
La distribución de arvenses está influenciada principalmente por el gradiente altitudinal y de precipitación; en menor medida por las condiciones de sitio, como la profundidad del suelo. Los resultados indican alta heterogeneidad en arvenses presentes en los cafetales debido a su capacidad de adaptación y dispersión.

