











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Las formaciones vegetales con mayor distribución en México son los matorrales xerófilos, cubriendo el 28.8% de la superficie total nacional (CONAFOR, 2018). Los estados con mayores superficies de matorrales son Baja California, Baja California Sur, Sonora, Chihuahua, Jalisco, Guanajuato, Tamaulipas y Nuevo León (Instituto Nacional de Estadística y Geografía, 2012). El estado de Nuevo León presenta una superficie de 4.2 millones de hectáreas, de las cuales el 53.5% corresponde a matorrales xerófilos (CONAFOR, 2014).
Las actividades productivas como la agricultura, ganadería y silvicultura han modificado la estructura y composición de los ecosistemas del norte del país (Alanís Rodríguez et al., 2008). La ganadería es una de las principales actividades que fomenta la pérdida de cobertura vegetal natural por el cambio de uso de suelo (Arriaga, 2009; Pequeño-Ledezma et al., 2012). El pastoreo continuo es una práctica sumamente utilizada en el noreste del país en zonas denominadas “agostaderos” (Hanselka et al., 1988), término utilizado para tierras que por su precipitación pluvial, topografía o calidad, producen en forma natural o cultivada, pastos y forrajes que sirven para alimento del ganado. En este tipo de pastoreo se hace uso de la superficie al menos durante una temporada completa o de forma permanente (Molina-Guerra et al., 2013), sin embargo, el animal ejerce una amplia selección de las especies del agostadero consumiendo aquellas que resultan más palatables (Baraza & Valiente-Banuet, 2012).
En las áreas rurales del noreste de México la actividad pecuaria es el principal medio de sustento, sin embargo, existe escasa información sobre los efectos de esta actividad en la vegetación. Generar información cuantitativa de la estructura y diversidad de las especies leñosas es importante para la correcta toma de decisiones en la gestión pecuaria y de conservación (Molina-Guerra et al., 2013; Pequeño-Ledezma et al., 2012).
El Área de Conservación e Investigación RENAC es una reserva ecológica particular ubicada en el noreste de México, presenta una superficie de 6.32 ha-1 y desde hace una década conserva una comunidad vegetal de matorral xerófilo que tiene historial de uso pecuario. En esta área de conservación no se han desarrollado investigaciones que evalúen el estado actual de su comunidad vegetal. El objetivo de la presente investigación fue evaluar la estructura y diversidad de especies leñosas en dos estratos (remanente y regeneración) del Área de Conservación e Investigación RENAC. La hipótesis es que las especies con mayor abundancia en ambos estratos (remanente y regeneración) son similares y que las curvas de dominancia-diversidad presentarán el mismo modelo de ajuste.
Métodos
Área de estudio. El Área de Conservación e Investigación RENAC (ACIRENAC), se localiza en el ejido Los Álamos, en el municipio de Linares, Nuevo León (Noreste de México; Fig. 1), en las coordenadas 27° 41’ 42.1” N y 99° 41’ 20.1” W, a 27 km de la cabecera de este municipio. Se encuentra en una zona de transición entre el matorral espinoso tamaulipeco (MET) y matorral submontano (INEGI, 1988) en la cuenca San Fernando - Soto la Marina, en la subcuenca A. Camacho (INEGI, 2010). Presenta escurrimientos superficiales de tipo intermitente próximos con un coeficiente de escurrimiento de 10-20%. INEGI (2005) lo ubica en la unidad geológica compuesta principalmente de rocas de origen sedimentario. El tipo de suelo predominante es del tipo Luvisol y las topoformas próximas son de lomerío bajo.

Fig. 1 a) Estados Unidos Mexicanos resaltando el Estado de Nuevo León, b) Estado de Nuevo León resaltando el municipio de Linares, c) Municipio de Linares, ubicando Área de Conservación e Investigación RENAC (ACIRENAC).
Análisis de la vegetación. La vegetación presente en el Área de Conservación e Investigación RENAC tiene características de transición de un matorral espinoso tamaulipeco y un matorral submontano. La vegetación fue influenciada por actividad pecuaria (ganado vacuno) durante el período 1984 al 2009, basada en el consumo del sotobosque y follaje del componente arbóreo y arbustivo, y por la extracción de leña (troncos caídos y corta de ramas de árboles y arbustos) para consumo local. En el año 2009 se excluyó el área con una cerca perimetral para delimitar el ACIRENAC de los terrenos contiguos y con esto evitar la extracción de leña y el ingreso del ganado; a partir de entonces y hasta la fecha (2019) el crecimiento de la vegetación se ha regenerado de manera natural y gradualmente.
Para evaluar la comunidad vegetal, en septiembre de 2019 se establecieron 10 sitios de muestreo de 100 m2 (10 x 10 m) distribuidos aleatoriamente en una superficie de 6.32 ha-1. En los sitios se consideraron los individuos arbóreos y arbustivos remanentes del aprovechamiento, con diámetro basal (d 0.10) igual o mayor a 3 cm. Para evaluar la regeneración de los árboles y arbustos (d 0.10 < 3 cm) se establecieron sitios de 1 m2 (1 x 1 m) en cada esquina y el centro del sitio de 100 m2.
En los sitios de muestreo se desarrolló un censo de las especies arbóreas y arbustivas; las medidas consideradas fueron altura total (h) (para el cálculo del volumen m3 ha-1), diámetro basal (d 0.10) y diámetro de copa (k), el cual se obtuvo utilizando una cinta métrica midiendo el espacio ocupado por la copa en sentido norte-sur y este-oeste, esto permite calcular el diámetro promedio y con ello determinar el área de copa (m2 ha-1). La medición del diámetro basal (d 0.10) se efectuó a 0.10 m sobre la base del suelo, ya que es una medida estándar empleada para la regeneración de las especies leñosas del matorral espinoso tamaulipeco (Alanís-Rodríguez et al., 2020). Las especies se identificaron por personal calificado de la Universidad Autónoma de Nuevo León usando el libro de Molina-Guerra et al. (2019).
Análisis de la información. Para estimar la riqueza de especies se utilizó el índice de Margalef (D Mg ) y para la diversidad alfa el índice de Shannon & Weiner (H’) mediante las ecuaciones:
Donde S es el número de especies presentes, N es el número total de individuos, p i es la abundancia proporcional de la iésima especie y n i es el número de individuos de la especie i.
Para realizar el análisis de similitud florística de las comunidades i y j, se empleó el índice de Jaccard mediante la ecuación:
Donde A es el número de especies presentes en la vegetación remanente (comunidad i), B es el número de especies presentes en la regeneración (comunidad j) y C es el número de especies presentes en ambos estratos (remanente y regeneración) es decir, que están compartidas.
Para caracterizar de manera horizontal la comunidad vegetal se generó un gráfico de clases diamétricas. Se generaron gráficos distribución de abundancia de especies a través de la construcción de curvas de dominancia-diversidad de forma decreciente para cada estrato (remanente y regeneración), desde las especies más abundantes hasta las menos abundantes (Brower & Zar, 1984; Magurran, 2004; Martella et al., 2012). Cada especie presenta un rango en el eje “x”, y su respectiva abundancia sobre el eje “y”. Las curvas se ajustaron a distintos modelos de distribución de abundancia de especies (geométrico, Preemption, lognormal, Zipf y Zipf-Mandelbrot (Ulrich, Ollik, & Ugland, 2010). Los modelos de los gráficos distribución de abundancia se eligieron a partir de la máxima verosimilitud según el criterio de información de Akaike (AIC), donde el menor valor de AIC indica el mejor ajuste de las curvas entre los modelos comparados (Oksanen et al., 2017). Los gráficos se realizaron mediante el programa R versión 2.4-3.
Para evaluar el diámetro promedio de copa y el área de copa y basal se utilizaron las siguientes fórmulas (Alanís-Rodríguez et al., 2020):
Donde
El volumen (V) de cada individuo se estimó mediante la fórmula:
donde g es la superficie supuesta circular del diámetro basal (d0.10 m), h la altura total de individuo y CM el coeficiente mórfico, que para el caso del matorral espinoso tamaulipeco es 0.5 (Alanís-Rodríguez et al., 2020).
Se utilizaron los indicadores ecológicos de abundancia (N*ha-1), dominancia (m2*ha-1) y frecuencia, con el fin de determinar el valor de importancia de las especies de la comunidad vegetal (Magurran, 2004). Para la estimación de la abundancia se empleó la siguiente ecuación:
Donde A i es la abundancia absoluta, AR i es la abundancia relativa de la especie i respecto a la abundancia total, N i es el número de individuos de la especie i, y S la superficie de muestreo (ha).
La dominancia relativa se evalúo mediante:
Donde D i es la dominancia absoluta, DR i es la dominancia relativa de la especie i respecto a la dominancia total, Ab el área de copa de la especie i y S la superficie muestreada (ha).
La frecuencia relativa se obtuvo con la siguiente ecuación:
Donde F i es la frecuencia absoluta, FR i es la frecuencia de la especie i respecto a la frecuencia total, P i es el número de sitios en la que está presente la especie i y NS el número total de sitios de muestreo. El índice de valor de importancia (IVI) se define como:
Resultados
Composición. En la vegetación remanente, se registraron 8 familias, 10 géneros y 11 especies (Tabla 1). La familia con mayor número de especies fue Fabaceae, con Acaia rigidula, Acacia farnesiana, Havardia pallens y Eysenhardtia texana.
Tabla 1 Listado florístico (ordenado alfabéticamente de acuerdo a la familia) de la vegetación remanente y de la regeneración del área de estudio. Abreviaturas: Vr (vegetación remanente), Rg (Regeneración) Arb (Arbustiva), P (Palatable) T (Tóxica) Sin descripción (N/D). La palatabilidad es de acuerdo a Martínez et al. (1997).
| Familia | Nombre Científico | Nombre común | Vr | Rg | Forma de Vida | Palatabilidad |
|---|---|---|---|---|---|---|
| Cannabaceae | Celtis laevigata Willd. | Palo blanco | x | x | Arb | N/D |
| Ebenaceae | Diospyros texana Sheele. | Chapote | x | x | Arb | N/D |
| Fabaceae | Eysenhardtia texana Sceele | Vara dulce | x | x | Arb | N/D |
| Fabaceae | Havardia pallens (Benth.) Britton & Rose | Tenaza | x | x | Arb | P |
| Fabaceae | Acacia farnesiana (L.) Willd. | Huizache | x | Arb | P | |
| Fabaceae | Acacia rigidula Benth. | Chaparro prieto | x | Arb | P | |
| Oleaceae | Forestiera angustifolia Torr. | Panalero | x | Arb | N/D | |
| Rhamnaceae | Ramnus humboldtiana Willd. ex Schult. | Coyotillo | x | x | Arb | T |
| Rubiaceae | Randia obcordata S. Watson. | Crucillo | x | x | Arb | N/D |
| Rutaceae | Amyris texana (Buckley) P. Wilson. | Barreta china | x | Arb | P | |
| Rutaceae | Zanthoxylum fagara (L.) Sarg. | Colima | x | x | Arb | N/D |
| Salicaceae | Neopringlea integrifolia (Hemsl.) S.Watson | Corvagallina | x | x | Arb | N/D |
| Ulmaceae | Celtis pallida Torr. | Granjeno | x | x | Arb | P |
| Verbenaceae | Lantana camara L. | Lantana | x | Arb | P |
En la regeneración, se registraron 11 familias, 12 géneros y 12 especies (Tabla 1). Se observaron dos familias con dos especies cada una; Rutaceae con Amyris texana y Zanthoxylum fagara, y Fabaceae con Havardia pallens y Eysenhardtia texana. Las otras ocho familias presentaron un solo espécimen cada una.
Riqueza y diversidad. Los resultados de la diversidad alfa en el Área de Conservación e Investigación RENAC (ACIRENAC) calculados mediante el índice de diversidad de Shannon (H´) fue de 1.92 para el total de la vegetación, 2.00 para la regeneración y 1.58 para la vegetación remanente. El índice de diversidad de Margalef (D Mg ) para el total de la vegetación resultó en un valor de 2.23, mismo valor que se obtuvo en la regeneración, el valor obtenido para la vegetación remanente fue de 1.89.
Diversidad beta. El Análisis de similitud florística evaluado mediante el Índice de Jaccard (J ij ) muestra que existe una similitud florística del 64.29%, donde nueve de 14 especies son comunes.
Indicadores Ecológicos. La abundancia absoluta fue 1950 N ha-1, la especie más abundante de la vegetación remanente del ACIRENAC es Havardia pallens, con el 41.54% del total de los individuos registrados, seguida de Zanthoxylum fagara con 32.31% así como Acacia farnesiana y Celtis laevigata con 6.15 y 4.62% respectivamente (Tabla 2). El área basal fue de 5.90 m2 ha-1 y el área de copa 16110 m2 ha-1, donde las tres principales especies (Havardia pallens, Zanthoxylum fagara y Acacia farnesiana) presentan el 75.40 y el 76.99% respectivamente. El volumen es de 14.70 m3 ha-1, donde Havardia pallens y Zanthoxylum fagara presentan el 56.23%. Las cinco especies con mayor IVI fueron Havardia pallens, Zanthoxylum fagara, Acacia farnesiana, Celtis pallida y Celtis laevigata, las cuales juntas representan el 86.57% del valor de importancia de la vegetación remanente del ACIRENAC.
Tabla 2 Nombre científico con datos de abundancia, frecuencia, dominancia en IVI de la vegetación remanente en el ACIRENAC.
| ESPECIES | ABUNDANCIA | ÁREA BASAL | ÁREA DE COPA | FRECUENCIA | VOLUMEN | IVI | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nombre científico | N ha-1 | % | m2 ha-1 | % | m2 ha-1 | % | Sitios | % | m3 ha-1 | % | |
| Havardia pallens | 810.00 | 41.54 | 1.84 | 31.19 | 5603.29 | 34.78 | 9.00 | 20.00 | 4.37 | 29.75 | 32.11 |
| Zanthoxylum fagara | 630.00 | 32.31 | 1.62 | 27.36 | 5492.42 | 34.09 | 10.00 | 22.22 | 3.89 | 26.48 | 29.54 |
| Acacia farnesiana | 120.00 | 6.15 | 1.00 | 16.85 | 1308.32 | 8.12 | 6.00 | 13.33 | 2.50 | 16.98 | 9.20 |
| Celtis pallida | 90.00 | 4.62 | 0.38 | 6.41 | 1316.82 | 8.17 | 5.00 | 11.11 | 0.92 | 6.25 | 7.97 |
| Celtis laevigata | 120.00 | 6.15 | 0.54 | 9.18 | 964.78 | 5.99 | 5.00 | 11.11 | 1.53 | 10.40 | 7.75 |
| Eysenhardtia polystachya | 60.00 | 3.08 | 0.20 | 3.44 | 332.82 | 2.07 | 3.00 | 6.67 | 0.52 | 3.55 | 3.94 |
| Diospyros texana | 40.00 | 2.05 | 0.17 | 2.89 | 566.09 | 3.51 | 2.00 | 4.44 | 0.59 | 3.99 | 3.34 |
| Neopringlea integrifolia | 40.00 | 2.05 | 0.07 | 1.12 | 186.45 | 1.16 | 2.00 | 4.44 | 0.17 | 1.13 | 2.55 |
| Acacia rigidula | 20.00 | 1.03 | 0.07 | 1.13 | 179.00 | 1.11 | 1.00 | 2.22 | 0.17 | 1.14 | 1.45 |
| Rhamnus humboldtiana | 10.00 | 0.51 | 0.02 | 0.27 | 136.25 | 0.85 | 1.00 | 2.22 | 0.04 | 0.24 | 1.19 |
| Randia obcordata | 10.00 | 0.51 | 0.01 | 0.16 | 24.05 | 0.15 | 1.00 | 2.22 | 0.01 | 0.09 | 0.96 |
| Total general | 1950 | 100 | 5.90 | 100 | 16110 | 100 | 45 | 100 | 14.70 | 100 | 100 |
Las especies presentes en la regeneración de los sitios estudiados en el ACIRENAC presentaron una abundancia absoluta de 27400 N ha-1, donde destacan con mayor abundancia Celtis laevigata y Havardia pallens, con el 28.47% (7,800 N ha-1) y 26.28% (7,200 N ha-1) de la abundancia total observada respectivamente, seguida de Zanthoxylum fagara con 12.41% (3,400 N ha-1) (Tabla 3). Las especies que presentaron mayores valores de área basal y área de copa en la regeneración fueron Havardia pallens, Celtis laevigata y Zanthoxylum fagara, presentando el 62.28 y 66.16% de los valores de valores porcentuales de la comunidad. Las especies con mayor frecuencia fueron Celtis laevigata, Havardia pallens y Zanthoxylum fagara, representando el 67% del total en el ACIRENAC. Lantana camara y Celtis pallida, registraron la menor frecuencia en el área de estudio (juntas el 2.19%). Las cinco especies que en la regeneración mostraron mayor IVI fueron Havardia pallens, Celtis laevigata, Zanthoxylum fagara, Forestiera angustifolia y Randia obcordata, representan juntas el 80.13% del IVI en el área de estudio.
Tabla 3 Nombre científico con datos de abundancia, frecuencia y dominancia de la vegetación presente en la regeneración de los sitios estudiados del ACIRENAC.
| ABUNDANCIA | ÁREA BASAL | ÁREA DE COPA | FRECUENCIA | IVI | |||||
|---|---|---|---|---|---|---|---|---|---|
| Nombre científico | Nha-1 | % | m2 ha-1 | % | m2 ha-1 | % | Sitios | % | |
| Havardia pallens | 7200 | 26.28 | 0.66 | 30.31 | 2854.68 | 43.17 | 36 | 26.28 | 31.91 |
| Celtis laevigata | 7800 | 28.47 | 0.30 | 13.89 | 810.48 | 12.26 | 39 | 28.47 | 23.06 |
| Zanthoxylum fagara | 3400 | 12.41 | 0.39 | 18.08 | 709.80 | 10.73 | 17 | 12.41 | 11.85 |
| Forestiera angustifolia | 1600 | 5.84 | 0.17 | 8.00 | 681.53 | 10.31 | 8 | 5.84 | 7.33 |
| Randia obcordata | 1600 | 5.84 | 0.27 | 12.64 | 414.98 | 6.27 | 8 | 5.84 | 5.98 |
| Neopringlea integrifolia | 1200 | 4.38 | 0.12 | 5.38 | 469.27 | 7.10 | 6 | 4.38 | 5.28 |
| Amyris texana | 1600 | 5.84 | 0.07 | 3.01 | 112.31 | 1.70 | 8 | 5.84 | 4.46 |
| Eysenhardtia polystachya | 800 | 2.92 | 0.06 | 2.93 | 273.12 | 4.13 | 4 | 2.92 | 3.32 |
| Rhamnus humboldtiana | 1000 | 3.65 | 0.03 | 1.43 | 151.70 | 2.29 | 5 | 3.65 | 3.20 |
| Diospyros texana | 600 | 2.19 | 0.04 | 1.80 | 32.73 | 0.49 | 3 | 2.19 | 1.62 |
| Lantana camara | 400 | 1.46 | 0.05 | 2.35 | 85.61 | 1.29 | 2 | 1.46 | 1.40 |
| Celtis pallida | 200 | 0.73 | 0.00 | 0.18 | 17.11 | 0.26 | 1 | 0.73 | 0.57 |
| Total general | 27400 | 100 | 2.17 | 100 | 6613.3 | 100 | 137 | 100 | 100 |
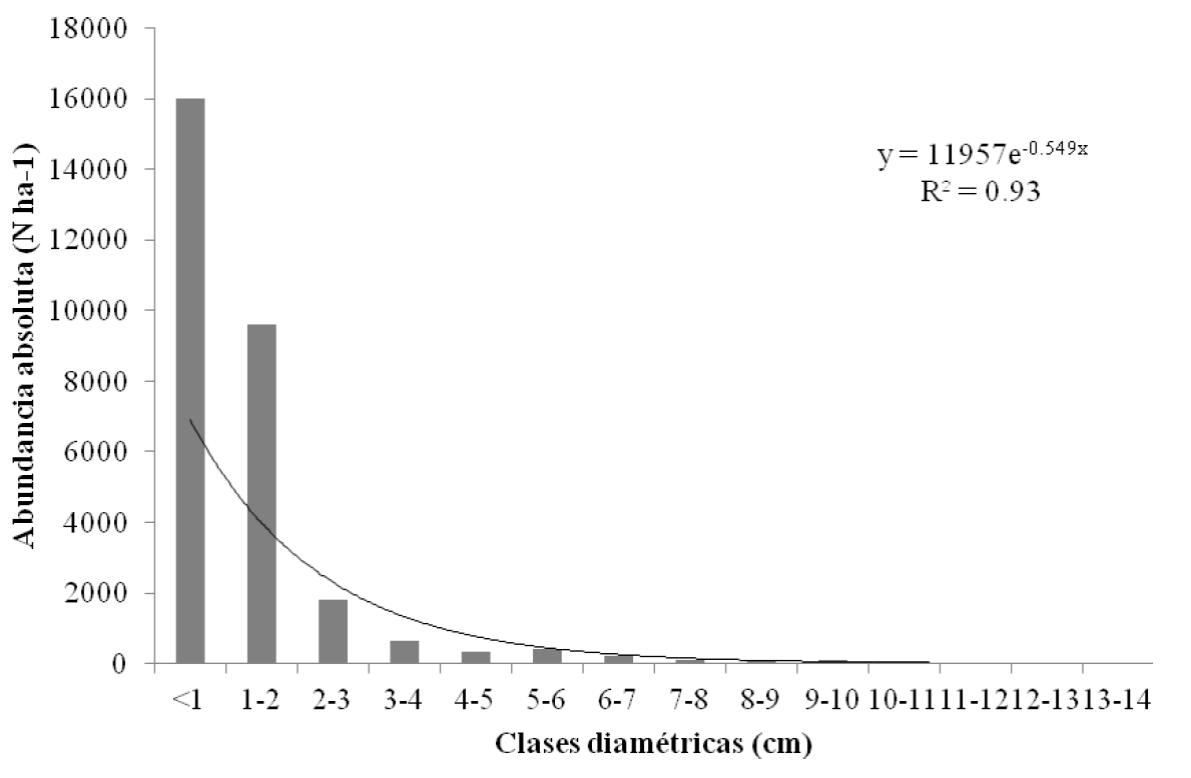
Respecto a la densidad de individuos por hectárea registradas en la comunidad vegetal (remanente y regeneración), se observa que existe un decremento exponencial (R2 = 0.93) conforme aumenta el diámetro de los mismos (Figura 2), siendo la clase menor a 1 cm de diámetro la que presentó mayor número de individuos con más de 650 N ha-1.
Respecto a las curvas de rango/abundancia para las dos condiciones remanente y regeneración (Fig. 3), de acuerdo al criterio de AIC, la vegetación remante mostró un mejor ajuste con el modelo de Preemption (AIC=273.97), perteneciente a la familia de la serie geomética, mientras que la curva de rango abundancia de la comunidad de la regeneración ajustó mejor al modelo de Zipf-Mandelbrot, de la familia de la serie logarítmica, (AIC = 1733.0). Las especies con mayor abundancia en la vegetación remanente fueron Havardia pallens (Fabaceae) y otras 4 especies de la misma familia y en la regeneración Celtis laevigata y la presencia de solo dos especies de la Familia fabaceae.

Discusión
El número de familias reportadas en nuestro estudio para la vegetación remanente y la regeneración (10 familias de especies arbóreas y arbustivas), se encuentra por debajo de las reportadas para el MET descrito en otros estudios para el municipio de Linares en sistemas maduros, con 14 (Mora Donjuán et al., 2013) y 15 familias (Domínguez Gómez et al., 2013). Sin embargo, están dentro del rango del número de familias (10-12) reportadas tanto para sistemas secundarios recuperados luego de usos de suelo moderados e intensivos respectivamente (Alanís Rodríguez et al., 2013), y como para prístinos con 10 familias observadas (García Hernández & Jurado, 2008). Esta variabilidad en el número de familias encontradas en el MET ha sido reconocida en diversos estudios, y explicada tanto por las distintas historias de uso del suelo a partir de las cuáles estas comunidades se recuperan (Alanís Rodríguez et al., 2013), así como también por la alta variabilidad en las condiciones edáficas y de relieve presentes en el municipio y en la región (Estrada-Castillón et al., 2011; García Hernández & Jurado, 2008). Como se observa en otros estudios del MET, en nuestro trabajo la familia Fabaceae fue la más diversa y mejor representada. Esto es explicado tanto por la variabilidad fisiográfica y topográfica del municipio (Mora Donjuán et al., 2013) como de la región (Estrada Castillón et al., 2004, 2005), así como por la gran capacidad que tiene esta Familia para soportar las condiciones propias de las zonas áridas, como el estrés hídrico (H. González-Rodríguez et al., 2011a; González-Rodríguez et al., 2011b; González Rodríguez & Cantú Silva, 2012) y el escaso nitrógeno presente en el suelo (Celaya & Castellanos-Villegas, 2011).
La vegetación remanente presentó 12 especies, un número inferior al registrado para el MET por Alanís-Rodríguez et al. (2013) en sistemas secundarios recuperados luego de usos como la agricultura (19 especies), la ganadería extensiva (24 especies), y la matarrasa (20 especies), pero similar a lo registrado por estos autores para ganadería intensiva (11 especies); asimismo, fue menor que las 21 especies reportadas por Mora-Donjuán et al. (2013) en sitios con un historial de uso similar, aunque con un mayor tiempo de recuperación (34 años de recuperación, vs 10 años de nuestro estudio), y al reportado para el MET en condiciones prístinas, en la cual fueron reportadas 16 especies por García-Hernández y Jurado (2008). El menor número de especies respecto a los sitios mejor conservados puede ser resultado del tipo de uso del suelo al que fue sometido el sitio. El ajuste al modelo preemption (serie geométrica) apoya en parte nuestra interpretación, toda vez que los modelos de la serie geométrica han sido utilizados como un indicador indirecto de impactos ambientales por distintos autores (Artigas, y Lima 2015). La regeneración presentó 11 especies, un número menor a la observada en la vegetación remanente, y también inferior a la observada en los casos anteriormente citados. Esto pudiera ser asociado con la presencia de un factor limitante adicional, que sería la luz. Quinteros et al. (2010) reconocen una disminución significativa en el número de especies en el sotobosque de un bosque de Nothofagus antarctica, asociada a las condiciones limitantes de luz. El ajuste de la curva de rango-abundancia de la regeneración al modelo Zipf-Mandelbrot refuerza nuestra interpretación; este tipo de distribución resulta cuando uno o unos pocos factores dominan la ecología de la comunidad (Pielou 1975; Magurran, 2004; Martella et al., 2012). Las curvas de acumulación de especies de la serie logarítmica sugieren la repartición diferencial de recursos por parte de las especies: En nuestro estudio, Celtis laevigata y la presencia de solo dos especies de la familia Fabaceae, con factor limitante luz (Fig. 4). Este modelo de serie logarítmica ha sido reportado para matorrales regenerados en España (Artigas y García, 2015), y maduros en el matorral espinoso tamaulipeco (Alanís-Rodríguez et al., 2018). Estos autores observaron que donde existe cubierta del dosel y además esta es densa, la riqueza y diversidad de especies disminuyen, y se presenta un aumento en la dominancia de algunas especies tolerantes a la sombra.
El área basal de la comunidad vegetal fue de 8.07 m2 ha-1 (remante 5.90 m2 ha-1 y regenerada 2.17 m2 ha-1), valor inferior a lo reportado por Leal et al. (2018) en una comunidad vegetal regenerada después de actividad pecuaria con mayor tiempo de abandono (20 años de recuperación, vs 10 años de nuestro estudio) con un valor de 13.76 m2 ha-1. El área de copa del estrato remante fue de 16,110 m2 ha-1, lo que indica una cobertura del dosel superior al 100% y por lo tanto una sobreposición de las copas. Esta condición de cobertura superior al 100% ha sido reportada por Mora Donjuán et al. (2013) y Graciano-Ávila et al. (2018) en el matorral espinoso tamaulipeco en condición prístina y madura, con valores de 13,973 m2 ha-1 y 16,671 m2 ha-1 respectivamente. El volumen en el estrato remante fue de 14.70 m3 ha-1, valor menor a lo registrado por Leal et al. (2018) en una comunidad vegetal regenerada después de 20 años a la actividad pecuaria con un valor de 26.74 m3 ha-1.
En este contexto, en nuestro estudio observamos como especies con mayor IVI en el dosel (correspondiente a la vegetación remanente) a Havardia pallens (32.11%), Zanthoxylum fagara (29.54%), y Acacia farnesiana (9.20%) (Tabla 3); mientras que en la regeneración dominaron Havardia pallens (31.91%), Celtis laevigata (23.06%) y Zanthoxylum fagara (11.85%) (Tabla 4). Si bien es cierto que en nuestro estudio la correspondencia entre las especies que dominan el dosel (conformado por la vegetación remanente), es alta respecto a las especies que dominan la regeneración, también lo es que existe evidencia de que éstas últimas son especies tolerantes a la sombra (Bush & Auken, 1986; Jurado et al., 2006; Souza et al., 2014); en particular Havardia pallens y Zanthoxylum fagara son especies que han sido reportadas como parte de la vegetación madura y prístina del MET (García-Hernández & Jurado, 2008; Mora Donjuán et al., 2013), mientras que Celtis laevigata en la vegetación homóloga de las planicies inundables del sur de Texas. La comunidad vegetal remanente del ACIRENAC coincide con las comunidades evaluadas por Estrada et al. (2005) y Domínguez et al. (2013), donde mencionan que Zanthoxylum fagara es un elemento estructuralmente importante en el matorral submontano, como también lo son en la comunidad contigua de matorral espinoso tamaulipeco. Además, el presente trabajo afirma lo que García-Hernández y Jurado (2008) mencionan, que Zanthoxylum fagara es la especie más abundante registrada en el matorral submontano del centro de Nuevo León.
Havardia pallens es una especie resistente a las actividades post pecuarias y los incendios forestales lo que puede también explicar su dominancia (Graciano-Ávila et al., 2018).
En las clases diamétricas, se observó un decremento exponencial en la abundancia de los individuos conforme el diámetro es mayor, mostrando un alto número de individuos menores a 6 centímetros. Esto indica que la comunidad vegetal presenta un alto número de individuos regenerando tal como lo observado por Mora-Donjuán et al. (2013) para un MET maduro. Esta información concuerda con los resultados de Pequeño-Ledezma et al. (2012) y Alanís et al. (2008) en comunidades vegetales del MET con historial de disturbio. Asimismo, la similitud florística intermedia-alta entre la vegetación remanente y la regeneración sugiere que las especies en común con mayor abundancia (Celtis laevigata, Havardia pallens y Zanthoxylum fagara) tienen la capacidad de establecerse en el estrato alto y también en el bajo, tolerando la sombra.
El valor del índice de diversidad de Shannon es similar al registrado por Mora-Donjuán et al. (2013), quienes evaluaron una comunidad vegetal madura del MET. Esto indica que, pese a que la comunidad vegetal evaluada presenta un historial de disturbio, la composición vegetal presenta una diversidad compleja, similar a la madura. El índice de diversidad de Shannon (H´) para el total de la vegetación del sitio fue de 1.92, sin embargo para el caso de la vegetación remanente y la regeneración obtuvimos unos valores de 2 y 1.58, respectivamente, esto se debe a que, a pesar de ser una comunidad vegetal que está regenerando después de un disturbio por actividad pecuaria, aún es pobre en la diversidad de especies, debido a la extracción selectiva de las mismas., lo que contrasta con los resultados presentados por Alanís Rodríguez et al. (2008) quienes presentaron resultados mayores a 3.
En el caso de riqueza de especies calculada mediante el índice de Margalef, los valores presentados por Mora-Donjuán et al. (2013) (D Mg ) 2.26 en una comunidad madura del matorral espinoso tamaulipeco, resultan muy similares a los obtenidos en el presente estudio (D Mg ) 2.23.
Conclusiones
De acuerdo con los resultados obtenidos en la presente investigación, se destacan las siguientes conclusiones: (1) a diez años del inicio de las actividades de restauración pasiva, consistente únicamente en la exclusión de fuentes de impactos, la comunidad vegetal estudiada presenta una riqueza y diversidad de especies típica en comparación con otras asociaciones vegetales de clima árido y semiárido del noreste de México, (2) tanto el patrón de clases diamétricas, como el índice de similitud de Jaccard sugieren que una activa dinámica de la regeneración en la comunidad vegetal, (3) la curva de dominancia-diversidad de las especies de la comunidad regenerativa sugiere que la regeneración de la comunidad se ajusta al patrón de dominancia-diversidad de la regeneración observada en matorrales relativamente en buen estado de conservación, y (4) la familia con mayor importancia por su contribución a la comunidad es la Fabaceae, mientras que las especies más importantes son Havardia pallens, Zanthoxylum fagara, y Celtis laevigata, familia y especies típicas de comunidades vegetales maduras del MET y submontano.

