











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El ajo (Allium sativum L.) es considerado uno de los cultivos más rentables en México (Macías-Duarte et al., 2010). Debido al tamaño del bulbo y su larga vida de anaquel, se crean dificultades en el manejo de enfermedades de postcosecha causadas por fitopatógenos ocasionando pérdidas económicas para los agricultores (Patón et al., 2017). Diferentes especies de Penicillium han sido reportadas como patógenos de ajo en diferentes países incluyendo a México (Cavagnaro et al., 2005; Fuentes et al., 2013; Overy et al., 2005). En la actualidad, el uso de fungicidas sintéticos es la estrategia principal para el control de patógenos durante la poscosecha (Singh & Sharma, 2018). Debido a lo anterior es necesaria la búsqueda de fungicidas que no generen efectos perjudiciales al medio ambiente y que sean efectivos contra fitopatógenos. En este sentido, el quitosano es el segundo polisacárido más abundante en la naturaleza, posee actividad antimicrobiana, es biodegradable y no es tóxico para los humanos ni animales (Gutierrez-Martinez et al., 2018). El quitosano ha sido efectivo contra patógenos aislados de guanábana (Ramos-Guerrero et al., 2018), aguacate (Chávez-Magdaleno et al., 2018), papaya (Hernández-López et al., 2018), entre otros. Por su parte, trabajos previos han demostrado un fuerte efecto inhibitorio al aplicar el peróxido de hidrógeno como estrategia para el control de patógenos como Listeria monocytogenes y Salmonella typhimurium en melón y arándanos respetivamente (Li et al., 2013; Upadhyay et al., 2013). También, es ampliamente conocida la actividad antifúngica de algunas sales orgánicas como son el BS (Bicarbonato de sodio) y el SP (Sorbato de potasio) en la inhibición de la germinación de esporas, la elongación del tubo germinal y la producción de enzimas pectinolíticas en varios patógenos tanto en pruebas in vitro como en evaluaciones in vivo (Mejía-Teniente et al., 2013; Yao et al., 2004; Youssef et al., 2012; Youssef & Roberto, 2014). Por lo anterior, se planteó como objetivo de este proyecto estudiar el efecto de Q (Quitosano), H2O2 y dos sales orgánicas aplicados de forma individual y en combinación como productos químicos alternativos en el control in vitro de P. citrinum.
Material y métodos
Aislamiento del patógeno
Las muestras de ajo morado (Allium sativum L.) fueron recolectadas en parcelas de la ciudad de León, Guanajuato, sin presencia de enfermedades ni daño mecánico. Los bulbos se colocaron en cámara húmeda (HR 90-95%) a 25ºC hasta el desarrollo de la enfermedad. Se realizaron pequeños cortes en las muestras de tejido con 50% de superficie sana y 50% de superficie dañada con la finalidad de garantizar que se tiene tejido infectado. Posteriormente, las secciones de tejido se desinfectaron superficialmente en una solución de hipoclorito de sodio al 1% por 3 minutos, se enjuagaron en agua destilada estéril y se colocaron sobre papel filtro. Las secciones de tejido fueron sembradas en cajas de Petri con medio APD (Agar papa dextrosa) (Difco ?) e incubadas a 25ºC ± 2 por siete días. Se realizaron explantes en función de los tipos y coloraciones de micelio desarrollados, hasta obtener un cultivo puro (Ochoa Fuentes et al., 2012).
Identificación morfológica de Penicillium sp.
Se recolectaron porciones de micelio con esporas que fueron observados al microscopio óptico (Motic, BA300) empleando los objetivos 40 y 100X, la identificación morfológica se realizó con base en las claves taxonómicas de Barnett & Hunter (1998) y Visagie et al. (2014).
Identificación molecular de Penicillium sp.
La extracción de ADN se realizó mediante el método propuesto por Doyle & Doyle (1990). Posteriormente, el ADN se amplificó mediante el método de reacción en cadena de la polimerasa (PCR, por sus siglas en inglés), empleando las regiones internas transcritas ITS1-5.8S-ITS2 utilizando el par de iniciadores de secuencia ITS4/ ITS5 siguiendo el protocolo propuesto por White et al. (1990). Las condiciones de la reacción de PCR fueron: 1 ciclo de desnaturalización inicial a 94°C por 5 min, 30 ciclos a 95°C por 10 segundos; 57°C por 30 segundos; 72°C por 2 min y 1 ciclo de extensión final a 72°C por 5 min. Los productos amplificados se visualizaron en un gel de agarosa al 1% mediante electroforesis. El producto de PCR fue purificado empleando PureLink® (Quick Gel Extraction and PCR Purification Combo Kit). Los pares de base obtenidos se compararon con las secuencias reportadas en la base de datos del banco de genes de NCBI (National Center for Biotechnology Information) mediante el programa BLAST.
Preparación de tratamientos
Las soluciones de quitosano peso molecular medio (desacetilación al 75-85%) (Sigma-Aldrich, St. Louis, MO, EE. UU.) se prepararon a 0.1, 0.5, 1.0 y 1.5% (p/v) en 100 mL de ácido acético al 2% (v/v), se agitó por 24 horas a temperatura ambiente, las soluciones se ajustaron a pH de 5.5 agregando NaOH al 1N y se añadieron 100 μL de Tween 80 (Ramos-Guerrero et al., 2018). Las soluciones de H2O2 (Sigma-Aldrich, St. Louis, MO, EE. UU.) se prepararon al 0.1, 0.5, 1.0, y 1.5% (v/v), por su parte las soluciones de BS (Jalmek, México) y SP (Jalmek, México) se prepararon a las siguientes concentraciones: 1.0, 1.5, 2.0, 2.5 y 3.0% (p/v), diluyendo el reactivo comercial en agua destilada. Todas las soluciones fueron esterilizadas en autoclave. Una vez determinada la efectividad antifúngica de los tratamientos aplicados de forma individual se procedió a seleccionar las combinaciones de tratamientos considerando que las concentraciones deberian estar por debajo del 50% (de las más efectivas) quedando de la siguiente manera: 0.05%Q+0.25%H2O2, 0.05%Q+1.0%SP, 0.05%Q+2.0%BS, 0.05%Q+2.0%BS+1.0%SP+0.25%H2O2, 1.0%SP+0.25%H2O2, 2.0%BS+1.0%SP y 0.25%H2O2+2.0%BS.
Evaluación de efectividad in vitro
Para evaluar el crecimiento micelial se tomó un disco de 7 mm de diámetro proveniente de la periferia de la colonia del hongo en estudio con crecimiento activo. Posteriormente, el disco se colocó en el centro de una caja de Petri con medio APD adicionado con el tratamiento; el control consistió en cajas APD-sin tratamiento. Una vez inoculadas las cajas Petri se incubaron a 25ºC y se midió el diámetro micelial en crecimiento con un vernier digital (TruperMR) cada 24 h, la prueba se detuvo cuando el hongo en la caja Petri control cubrió el 100% de la superficie. Los resultados se expresaron en porcentaje de inhibición del crecimiento micelial empleando la fórmula propuesta por Yahyazadeh et al. (2008). La prueba de esporulación se llevó a cabo a las 120 h de incubación del hongo fitopatógeno, empleando las mismas cajas de APD-con tratamientos. Se adicionaron 10 ml de agua destilada estéril y con una varilla de vidrio se raspo la superficie de las cajas; las esporas colectadas fueron transferidas a un tubo de ensayo. La suspensión de esporas se agitó en Vortex, se tomaron 20 µl y se colocaron sobre una cámara de Neubauer para cuantificar las esporas en un microscopio óptico (40X). Para evaluar la germinación se preparó una suspensión de esporas de una colonia del hongo de 8 días; ajustando la concentración a 1x108 esporas/mL. Se prepararon tubos Eppendorf que contenían medio APD líquido con los tratamientos. Cada tubo eppendorf fue inoculado con la misma concentración de esporas, posteriormente se incubaron a 26°C en agitación constante durante 14h. Se contaron las esporas germinadas y las no germinadas en una cámara de Neubauer cada dos horas hasta que el control germinó al 100%. Una espora ha germinado cuando el tubo germinativo sea igual o mayor a la longitud de ésta (Yao et al., 2004). Los resultados se reportaron en número de esporas ml-1. Todos los ensayos se realizaron por triplicado y los experimentos se repitieron dos veces.
Análisis estadístico
Para evaluar el crecimiento micelial y germinación de los tratamientos aplicados de forma individual se aplicó un diseño unifactorial de bloques. Se consideraron los diferentes factores que conforman el sistema de control: quitosano (con 5 niveles: 0.05%, 0.1%, 0.5%, 1.0% y 1.5%), peróxido de hidrógeno (con 7 niveles: 0.25%, 0.5%, 1.0%, 1.5%, 2.0%, 2.5% y 3.0%), bicarbonato de sodio (con 5 niveles: 1.0%, 1.5%, 2.0%, 2.5% y 3.0%), sorbato de potasio (con 5 niveles: 1.0%, 1.5%, 2.0%, 2.5% y 3.0%). Para las combinaciones se aplicó un diseño unifactorial de bloques para evaluar el crecimiento micelial, la esporulación y germinación, con el factor tratamiento con 7 niveles (0.05%Q+0.25%H2O2, 0.05%Q+1.0%SP, 0.05%Q+2.0%BS, 0.05%Q+2.0%BS+1.0%SP+0.25%H2O2, 1.0%SP+0.25%H2O2, 2.0%BS+1.0%SP y 0.25%H2O2+2.0%BS). Los resultados obtenidos se analizaron mediante un análisis de varianza (ANOVA, por sus siglas en inglés) donde de se realizaron comparaciones de medias por prueba de Tukey (α<0.05) empleando el paquete estadístico GraphPad.
Resultados
Identificación morfológica y molecular del patógeno
La colonia del hongo se desarrolló sobre medio APD a 25ºC de 5 a 9 días. El color inicialmente es blanco virando a verde-azulado. La textura de las colonias es pulverulenta y plegada (Fig. 1A). El reverso de la colonia es amarillento (Fig. 1B). A nivel microscópico el hongo presenta conidióforos simples, penicilados, que termina en un grupo de fiálides ampuliformes y conidias. Presenta ramificaciones secundarias conocidas como métulas que se forman sobre los conidios; de tres a cinco métulas por estípite (Fig. 1C). Las conidias o esporas son redondas, unicelulares y se observan como cadenas no ramificadas en el extremo de las fiálides. El patrón de ramificación observado fue biverticilada (Pitt & Hocking, 2009; Visagie et al., 2014).

Fig 1 Morfología de Penicillium sp. Estructura macroscópica: a) Anverso, b) Reverso y c) Estructura microscópica
La amplificación de ADN, por medio de PCR, produjo un producto de aproximadamente 701 pares de bases lo que permitió la identificación de la especie del hongo en estudio. Los porcentajes de similitud obtenidos en la base de datos del GenBank fueron del 99.39% (número de accesión: HQ891112.1) para Penicillium citrinum LXL-24, especie no reportada como patógeno de ajo.
Evaluación de efectividad in vitro
La aplicación de H2O2 inhibió en su totalidad el desarrollo del patógeno empleando concentraciones desde 0.5 hasta el 3.0% (Fig. 2a), observándose una diferencia significativa (p < 0.05) entre las concentraciones evaluadas. Con respecto al BS, los resultados muestran diferencia significativa (p < 0.05) entre las concentraciones aplicadas (Fig. 2b) y al aplicar la mayor concentración analizada (3%) se observó una total inhibición del hongo.

Fig 2 Efecto de los diferentes tratamientos en el desarrollo micelial de P. citrinum en APD. a) H2O2, B) BS, c) SP y d) Quitosano.
Por otra parte, los resultados muestran diferencia significativa (p < 0.05) entre las concentraciones aplicadas de SP (Fig. 2c). La máxima inhibición (87%) se obtuvo con una concentración del 3%.
Stanojevic et al. (2009) reportaron resultados similares al aplicar SP al 1.5% en el control de P. italicum.
En los tratamientos con quitosano se observa que al utilizar la concentración del 1% se logra una inhibición del 100% en el crecimiento radial del patógeno (Fig. 2d). El análisis estadístico de los datos muestra diferencia significativa entre los tratamientos (p < 0.05).
En las combinaciones de tratamientos, el análisis estadístico de los datos muestra diferencias significativas entre los tratamientos (p < 0.05). Se obtuvieron resultados favorables para las diferentes combinaciones probadas en la inhibición in vitro de P. citrinum con la excepción del tratamiento quitosano y peróxido de hidrógeno en la que la concentración de ambos compuestos no fue suficiente para presentar un control sobre el patógeno (Fig. 3).
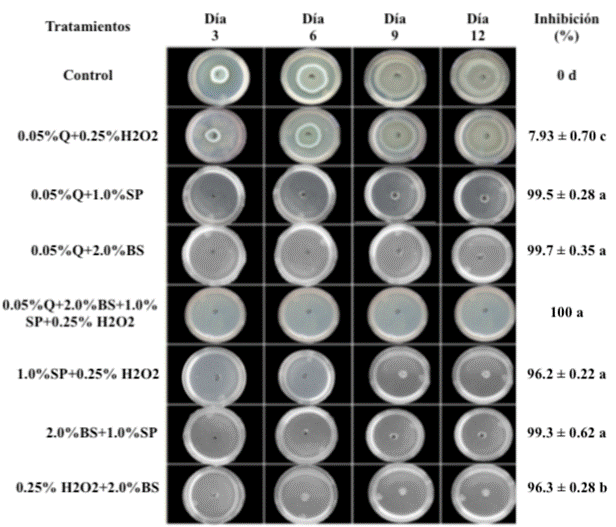
Fig 3 Efecto de la combinación de los diferentes tratamientos en el desarrollo micelial de P. citrinum
La germinación fue totalmente inhibida mediante la aplicación de los tratamientos individuales, excepto con la aplicación de quitosano (0.05% y 0.1%) (Tabla 1). La combinación de tratamientos fue efectiva en la inhibición de la producción del tubo germinal (Tabla 2).
La combinación de tratamientos resultó efectiva para el control de la esporulación, observándose diferencia estadística significativa, solo el tratamiento de quitosano combinado con H2O2 no resulto efectivo para detener el proceso de esporulación al evidenciar incluso una mayor esporulación con respecto al control (Tabla 2).
Tabla 1 Efecto de los tratamientos individuales en la germinación de conidios de P. citrinum.
| Germinación de esporas* (%) | |
|---|---|
| Control | 100 a |
| 0.25% H2O2 | 0.0±0.0b |
| 0.50% H2O2 | 0.0±0.0b |
| 1.0% H2O2 | 0.0±0.0b |
| 1.5% H2O2 | 0.0±0.0b |
| 2.0% H2O2 | 0.0±0.0b |
| 3.0% H2O2 | 0.0±0.0b |
| Control | 100a |
| 1.0% BS | 0.0±0.0b |
| 1.5% BS | 0.0±0.0b |
| 2.0% BS | 0.0±0.0b |
| 2.5% BS | 0.0±0.0b |
| 3.0% BS | 0.0±0.0b |
| Control | 100a |
| 1.0% SP | 0.0±0.0b |
| 1.5% SP | 0.0±0.0b |
| 2.0% SP | 0.0±0.0b |
| 2.5% SP | 0.0±0.0b |
| 3.0% SP | 0.0±0.0b |
| Control | 100a |
| 0.05% Q | 85±5.0ab |
| 0.1% Q | 84±6.0ab |
| 0.5% Q | 0.0±0.0b |
| 1.0% Q | 0.0±0.0b |
| 1.5% Q | 0.0±0.0b |
* Letras diferentes en la columna indican diferencias significativas (Tukey, p < 0.05)
Tabla 2 Efecto de los tratamientos combinados en la esporulación y germinación de conidios de P. citrinum.
| Tratamiento | Esporulación* (esporas x106/mL) | Germinación de esporas* (%) |
|---|---|---|
| Control | 4.36 ± 1.32 b | 100 a |
| 0.05%Q + 0.25%H2O2 | 7.66 ± 0.14 a | 0 b |
| 0.05%Q + 1.0%SP | 0.003 ± 0.001 c | 0 b |
| 0.05%Q + 2.0%BS | 0.0035 ± 0.002 c | 0 b |
| 0.05%Q + 2.0%BS + 1.0% SP + 0.25%H2O2 | 0 c | 0 b |
| 1.0% SP + 0.25%H2O2 | 0.007 ± 0.003 c | 0 b |
| 2.0%BS + 1.0% SP | 0.006 ± 0.002 c | 0 b |
| 0.25%H2O2 + 2.0%BS | 0.007 ± 0.004 c | 0 b |
* Letras diferentes en la columna indican diferencias significativas (Tukey, p < 0.05)
Discusión
La efectividad del H2O2 puede ser atribuida a daño celular masivo a nivel macromolecular; que incluye la ruptura de las capas de la membrana, inhibición enzimática, oxidación de nucleósidos, la interrupción de la síntesis de proteínas y, en última instancia, la muerte celular (Finnegan et al., 2010). Estos resultados son superiores a lo reportado por Qin et al. (2011), ya que al aplicar H2O2 al 1%, reportaron un 60% de inhibición del desarrollo micelial en P. expansum.
Se ha atribuido la capacidad inhibitoria del BS a una modificación del pH del entorno de crecimiento de los patógenos, lo que da lugar a la inactivación de enzimas extracelulares del fitopatógeno, un incremento en el estrés osmótico y una reducción en la presión de turgencia de la célula; resultado en un colapso y encogimiento de las hifas. En un reporte previo, la aplicación de BS al 0.6% inhibió el desarrollo micelial de P. expansum un 40% (Lai et al., 2015).
El efecto inhibitorio del SP parece ser debido al decremento del pH intracelular y a la ionización de moléculas ácidas afectando de esta manera el desarrollo del hongo (Gregori et al., 2008; Stanojevic et al., 2009).
La eficacia por la cual el quitosano muestra un efecto inhibitorio sobre el crecimiento micelial del hongo podría ser explicado por varios mecanismos de acción; el primero de ellos es debido a la naturaleza policatiónica del quitosano, que interacciona con los fosfolípidos que se encuentran en la pared celular del patógeno, lo que lleva a un aumento en la permeabilidad de la membrana plasmática y en segunda instancia se cree que el quitosano puede entrar en la célula fúngica, interactuar con el ADN, y alterar su conformación, por tanto, inhibir la síntesis de mRNA y proteínas (Kong et al., 2010; Ngo et al., 2015). Estos resultados concuerdan con lo informado por Wang et al. (2014) en el que se probaron concentraciones de 0.1% (1 mg/ml), 0.5% (5 mg/ml) y 1% (10 mg/ml) de quitosano sobre el crecimiento micelial de P. expansum patógeno del fruto de azufaifo (Ziziphus jujuba); reportando que la efectividad era dependiente de la concentración de quitosano empleada.
Respecto a las combinaciones de tratamientos, los resultados sugieren una interacción negativa entre el quitosano y el peróxido de hidrógeno, en este sentido Qin et al. (2002) mencionan que el H2O2 puede afectar en el quitosano el grado de polimerización y cambios en la estructura química del biopolímero como la formación de grupos carboxilo los cuales están relacionados con un proceso de desaminación debido a reacciones con radicales.
El mecanismo de acción de acción del quitosano para inhibir el proceso de germinación esta relacionado con las interacciones entre las cargas de los grupos -NH3 y las cargas negativas en la membrana celular causando una modificación de la permeabilidad de la misma, retardando el proceso de germinación de la espora (Li et al., 2015; Wang et al., 2014). Por su parte, El H2O2 es efectivo debido a que los radicales libres oxidan enzimas y proteínas provocando ruptura de cadenas de ADN (Minibayeva, Beckett, & Kranner, 2015). Se ha reportado que el BS aumenta el estrés osmótico de la célula provocando cambios estructurales como colapso y contracción de esporas (Lai et al., 2015). La actividad antifúngica del SP esta relacionado con la afectación del gradiente electroquímico de la membrana citoplasmática (transporte activo de sustancias), lo que a su vez inhibe el transporte de aminoácidos y de procesos enzimáticos (Youssef et al., 2012). Estos resultados son prometedores debido a que cualquier agente que pueda controlar el proceso de germinación coadyuva a evitar la diseminación del patógeno y disminuir la posibilidad de infecciones.
Finalmente, Hernández et al. (2007) y Reddy et al. (1998) mencionan que la excesiva esporulación del hongo sugiere una concentración sub-letal del tratamiento lo que da lugar a un proceso de estrés provocando la activación de mecanismos de defensa del patógeno asegurando la supervivencia hasta encontrar las condiciones favorables para su desarrollo. La eficacia de la combinación de los tratamientos puede ser atribuida en la capacidad de los compuestos de afectar el desarrollo del micelio y en consecuencia la formación estructuras de dispersión.
Conclusiones
De acuerdo a los datos y análisis se concluye que P. citrium es el agente causal de la pudrición azul en ajo morado (Allium sativum L). Además, el H2O2 al 0.5%, el BS al 3% así como el quitosano al 1% resultaron efectivos en la inhibición del crecimiento micelial y germinación del patógeno al 100%. Por su parte, en las combinaciones evaluadas se observó un efecto sinérgico empleando las concentraciones mínimas de cada compuesto a excepción de quitosano - H2O2. Debido a lo anterior, los tratamientos evaluados pueden ser una alternativa ecológica efectiva para el control de P. citrinum en ajo morado.

